
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ИЗБИРАТЕЛЬНАЯ АКТИВАЦИЯ ТРАНС-РЕПРЕССОРНОЙ ФУНКЦИИ ГЛЮКОКОРТИКОИДНОГО РЕЦЕПТОРА
В КЛЕТКАХ ГЕМОБЛАСТОЗОВ
Специальность 14.01.12 – онкология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
МОСКВА
201![]() 1
1
Работа выполнена в лаборатории механизмов химического канцерогенеза НИИ Канцерогенеза Учреждения Российской академии медицинских наук Российский онкологический научный центр имени РАМН (директор – академик РАН и РАМН, д. м.н., проф. )
|
Научный руководитель: доктор медицинских наук |
Якубовская Марианна Геннадиевна |
|
Официальные оппоненты: | |
|
доктор биологических наук, профессор |
Красильников Михаил Александрович |
|
доктор химических наук |
Евстафьева Александра Георгиевна |
|
Ведущая организация: |
Московский научно-исследовательский онкологический институт |
Защита диссертации состоится «18» ноября 2011 года в_10_ часов на заседании диссертационного совета Д.001.017.02 РОНЦ имени РАМН Москва, Каширское шоссе, 24
С диссертацией можно ознакомиться в библиотеке РОНЦ им. РАМН.
Автореферат разослан «___» октября 2011 года
 Ученый секретарь диссертационного совета
Ученый секретарь диссертационного совета
![]()
![]() д. м.н., профессор
д. м.н., профессор
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы
Гемобластозы занимают 5-6 место среди всех онкологических заболеваний и составляют 50% злокачественных новообразований у детей. Классическая химиотерапия гемобластозов включает в себя цитостатические препараты, и ее стандартным компонентом являются глюкокортикоиды (GC). Они являются важными физиологическими и фармакологическими регуляторами роста и жизнедеятельности клеток крови. Высокие дозы GC применяются для снижения количества бластных клеток в острой фазе заболевания при индукции ремиссии. Для успешной консолидации ремиссии в протокол химиотерапии также включают GC. Эти препараты обладают не только системным противовоспалительным действием на организм, но и специфической способностью вызывать гибель лимфоидных клеток.
Биологический эффект GC осуществляется посредством активации глюкокортикоидного рецептора (GR), хорошо изученного транскрипционного фактора. GR регулирует экспрессию генов по двум механизмам – транс-активационному и транс-репрессорному. Терапевтическое действие GC в основном реализуется через ДНК-независимую транс-репрессию – белок-белковое взаимодействие GR с факторами транскрипции (NF-κB, AP-1 и пр.). Эти транскрипционные факторы являются эффекторами медиаторов воспаления, антиапоптотических генов и молекул клеточной адгезии. Под действием GC наблюдается снижение активности этих факторов и их эфферентных путей, что ведет к подавлению пролиферации опухолевых клеток. Развитие многих осложнений терапии GC, связано с транс-активацией, опосредованной взаимодействием гомодимера GR с палиндромными респонсивными элементами в промоторах и энхансерах генов, участвующих в регуляции метаболизма. Недавно были описаны селективные агонисты GR (SEGRA). Эти соединения связываются с GR, но активированный ими рецептор реализует свое действие в основном по транс-репрессорному механизму. Несколько соединений, обладающих подобными свойствами, в настоящее время проходят доклинические или клинические испытания в качестве противовоспалительных нестероидных препаратов со сниженными побочными эффектами. К этому типу соединений относится 2-(4-ацетоксифенил)-2-хлор-N-метилэтиламмоний хлорид, или CpdA, более стабильный аналог предшественника азиридина, выделенный из африканского растения Salsola tuberculatiformis Botschantzev. Для проведения данного исследования CpdA был предоставлен проф. , университет Норсвестерн, Чикаго, США.
Применение SEGRA представляется наиболее целесообразным при лечении гемобластозов, однако антипролиферативные и проапоптотические эффекты этих соединений на клетки лейкозов и лимфом не изучены. Таким образом, изучение действия CpdA на клетки лейкозов в сравнении со стандартными GC, используемыми в клинической онкологии, представляет собой приоритетное перспективное исследование, направленное на усовершенствование химиотерапии гемобластозов.
В настоящее время при лечении онкологических заболеваний обычно применяется комбинированная химиотерапия, при которой одновременное использование двух и более препаратов позволяет снизить их дозы, и добиться их синергического или аддитивного эффектов. В частности, было продемонстрировано потенцирующее действие GC при лечении гемобластозов Винкристином, Адриамицином, Морфоланом и другими препаратами. Одним из самых перспективных современных противоопухолевых препаратов для химиотерапии гемобластозов является Бортезомиб (BZ), который начали использовать в США и Европе в 2003–2004 гг. Этот препарат активно применяется при терапии множественных миелом и лимфом, а также было продемонстрировано его успешное использование при лечении лейкозов. BZ относится к классу ингибиторов протеасом: он связывается с каталитическим доменом 26S протеасомы, предотвращая расщепление определенных клеточных белков, и, в частности, GR. Было установлено, что ингибиторы протеасом индуцируют апоптоз в опухолевых клетках, не влияя на нормальную ткань. При этом наблюдается подавление активности NF-kB, стресс эндоплазматического ретикулума и связанные с ним реакции. При сочетанном применении глюкокортикоидов и BZ был показан синергизм антипролиферативных и проапоптотических эффектов на клетках множественной миеломы, лимфом и лейкозов. Поскольку мы полагали, что СpdA активирует транс-репрессию, в частности снижает активность NF-kB, то была выдвинута гипотеза, что Бортезомиб может оказывать потенциирующее действие на эффекты данного селективного агониста GR. Это объясняет наш интерес к изучению совместного действия CpdA, модулятора GR, и BZ.
Таким образом, настоящая работа, посвященная исследованию эффектов нового нестероидного лиганда GR, CpdA, на клеточные линии гемобластозов, сравнению их с эффектами GC, а также оценке совместного действия CpdA и ингибитора протеасом Бортезомиба, представляет собой актуальную задачу современной экспериментальной онкологии и молекулярной биологии.
Основные цели и задачи исследования
Целью настоящей работы являлась оценка противоопухолевого потенциала нового модулятора GR, CpdA, в клетках лейкозов и лимфом, а также исследование совместного антипролиферативного и проапоптотического действия Бортезомиба и CpdA.
Для достижения поставленной цели были выдвинуты следующие задачи:
1. Охарактеризовать клеточные линии лейкозов СЕМ и К562, и лимфом NCEB, Granta и Jeko, по экспрессии и функциональной активности GR.
2. Изучить закономерности антипролиферативного и проапоптотического действия нестероидного аналога GC, CpdA, в клетках лейкозов и лимфом.
3. Оценить генотоксический эффект СpdA.
4. Определить транс-активационный и транс-репрессорный потенциал CpdA при его действии на клетки лейкозов и лимфом.
5. Оценить потенциальную способность CpdA вызывать развитие резистентности в сравнении с GC.
6. Изучить закономерности антипролиферативного и проапоптотического действия различных комбинаций BZ и CpdA.
7. Исследовать влияние BZ на способность CpdA избирательно активировать GR-опосредованную транс-репрессию.
Научная новизна и практическая значимость исследования
При выполнении данного исследования был получен ряд приоритетных данных. В частности, впервые было показано, что клеточные линии лейкозов СЕМ и К562, и лимфом NCEB, Granta и Jeko, широко используемые при изучении новых противоопухолевых препаратов, экспрессируют функционально активный GR, не несущий в себе полиморфизмов и мутаций. Используя данные клеточные линии в качестве модельной системы, было продемонстрировано антипролиферативное и проапототическое действие нового нестероидного лиганда GR, CpdA, на клетки лейкозов и лимфом. В то же время было показано, что длительное применение CpdA не вызывает уменьшения экспрессии GR и, как следствие, развития резистентности. Способность CpdA вызывать транс-репрессию ряда генов, но не запускать транс-активацию были продемонстрированы на клетках гемобластозов также впервые. Было показано, что механизм транс-репрессии, запускаемый CpdA, является GR-зависимым процессом. Важными приоритетными данными являются результаты, свидетельствующие о кооперативном антипролиферативном и проапоптотическом эффектах CpdA и Бортезомиба на клетки лейкозов и лимфом, а также о потенцирующем действии BZ на способность CpdA избирательно активировать GR-опосредованную транс-репрессию.
Апробация работы
Диссертация апробирована и рекомендована к защите 14 сентября 2011 года на совместной научной конференции отдела химического канцерогенеза, отдела трансформирующих генов опухоли, лаборатории механизмов прогрессии эпителиальных опухолей, лаборатории молекулярной эндокринологии, лаборатории онкогеномики, лаборатории цитогенетики НИИ Канцерогенеза РОНЦ им. РАМН.
Публикации
По теме диссертации опубликовано 7 печатных работ, в том числе 4 статьи опубликованы в журналах, рекомендованных ВАК МОН РФ соискателям ученой степени кандидата биологических наук.
Структура и объем диссертации
Диссертация изложена на 138 страницах машинописного текста, содержит 36 рисунков, 4 таблицы, состоит из глав: «Введение», «Обзор литературы», «Материалы и методы», «Результаты исследования», «Обсуждение результатов», «Выводы», «Приложения», «Список литературы».Список литературы включает 273 источника, в том числе 8 публикаций в отечественных рецензируемых изданиях.
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
ОБЗОР ЛИТЕРАТУРЫ
Обзор литературы включает современные данные о классификации гемобластозов, наиболее распространенных в этих типах опухолей генетических нарушениях и особенностях терапии различных форм этих заболеваний. Вторая часть обзора посвящена современным представлениям о стурктуре и функциях глюкокортикоидного рецептора, и двух основных механизмах его действия в клетках гемобластозов – транс-активационном и транс-репрессорном. Описаны различные типы лигандов глюкокортикоидного рецептора и их особенности в регуляции механизмов его действия на опухолевые клетки.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использованы следующие методы исследования: проточная цитофлуориметрия, выделение ДНК, РНК и белков, иммуноблоттинг, секвенирование, определение активности эукариотической топоизомеразы I, бактериальный тест Эймса на мутагенную активность, полимеразная цепная реакция, обратная транскрипция, молекулярное клонирование, трансформация бактериальных клеток, транзиентная трансфекция, трансдукция лентивирусных векторов, измерение активности люциферазы, флуоресцентная микроскопия, статистическая обработка данных.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
1. Характеристика модельной системы лейкозов и лимфом человека
Для исследования эффектов CpdA на клетки гемобластозов были выбраны клеточные линии, происходящие от Т - или В-лимфоцитов, а также клетки миелоидного ростка.
В качестве модельной системы были использованы линии клеток СЕМ (острый лимфобластный лейкоз), К562 (хронический миелобластный лейкоз), NCEB, Granta и Jeko (диффузная В-клеточная лимфома из клеток мантийной зоны). Культура клеток СЕМ характеризуется полиморфностью и способностью давать клоны с различными хромосомными аномалиями, которые приводят к чрезмерной активации пропролиферативных и антиапоптотических сигнальных путей [van Etten et al, 2007, Tefferi et al, 2007, Chiaretti and Foa, 2009]. Особенностью клеток линии К562 является наличие хромосомной транслокации t(9;22), известной под названием «филадельфийская хромосома» [Klein et al., 1976]. Отличительной чертой клеток NCEB, Granta и Jeko является хромосомная транслокация между 11 и 14 хромосомами, t(11;14)(q13;q32), которая приводит к перегруппировке в гене Bcl-1 и гиперэкспрессии циклина D1 [Jeon et al, 1998], что в свою очередь приводит к избыточной пролиферации.
В литературе были описаны следующие причины развития резистентности к терапии GC для линии СЕМ: (1) нарушение рецептора в результате точечных мутаций; (2) низкая экспрессия функционально активного рецептора [Schmidt et al, 2006, Ramos et al, 1999, Ruiz et al, 2001, Beesley et al, 2009, Irving et al, 2005]; (3) преобладание неполноразмерных изоформ GR [Zawydiwski et al, 1983, Thompson et al, 1986, Thompson et al, 1987].
В связи с этим были получены данные, свидетельствующие о полном соответствии последовательности нуклеотидов в 2-9 экзонах гена GR субклона клеточной линии СЕМ, полученной из банка замороженных образцов РОНЦ им. . Было продемонстрировано, что данный субклон линии СЕМ не несет мутаций в гене GR, и экспрессия полноразмерного функционального рецептора представлена на высоком уровне.
Линия К562, имеющая признаки гранулоцитарной, эритроцитарной и Т-клеточной дифференцировки, использовалась ранее в исследованиях эффектов GC, но наличие полиморфизмов и точечных мутаций гена GR в клетках этой линии ранее не проводилось [Agius et al, 1982]. Линии В-клеточного происхождения NCEB, Granta и Jeko широко используются в качестве модельных систем лимфом. Однако литературных данных о структурных и функциональных особенностях GR в указанных клетках не было. В представленной работе впервые было продемонстрировано, что последовательность нуклеотидов гена GR клеточных линий К562, NCEB, Granta и Jeko соответствует дикому типу, то есть не содержит полиморфизмов и точечных мутаций.
Все используемые в работе клеточные линии отличались по уровню экспрессии GR: по данным денситометрического анализа иммуноблотов, наибольшее количество рецептора было зафиксировано в клетках СЕМ и NCEB, в то время как в клетках линии Jeko уровень экспрессии GR был очень низким (рис. 1).

Рис.1. Экспрессия GR в клетках СЕМ, К562, NCEB, Granta и Jeko. Уровень экспрессии GR в клетках определяли методом иммуноблоттинга. Денситометрический анализ блотов проводили с помощью пакета программ Odyssey v2.1
После действия глюкокортикоидов на клетки СЕМ, К562, Granta, NCEB и Jeko количество GR падало, что согласуется с литературными данными о снижении экспрессии рецептора после обработки клеток GC (рис. 2) [Chen et al, 2010, Wallace et al, 2001].
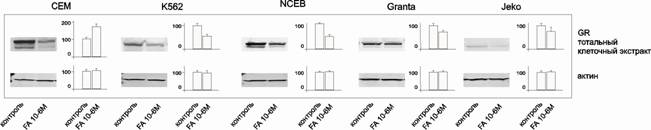
Рис.2. Влияние FA на количество GR в клетках CEM, K562, NCEB, Granta и Jeko. Уровень экспрессии GR определяли методом иммуноблоттинга. Денситометрический анализ блотов проводили с помощью пакета программ Odyssey v2.1
Ядерная транслокация GR под действием глюкокортикоидов является еще одним важным показателем его функциональной активности. Было показано, что использованные в нашей работе клеточные линии лейкозов и лимфом содержат рецептор, способный к перемещению в ядро после активации GC (рис. 3).
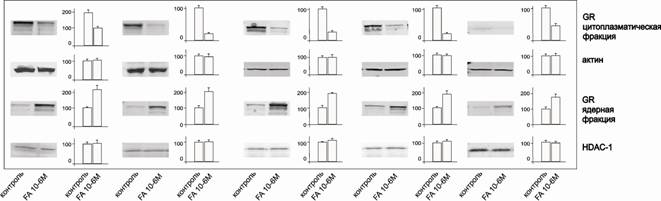
Рис.3. Ядерная транслокация GR. Уровень транслокации GR в ядро определяли методом иммуноблоттинга ядерной и цитоплазматической фракций. Денситометрический анализ блотов проводили с помощью пакета программ Odyssey v2.1
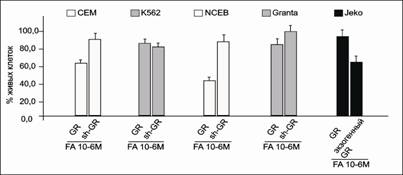
Важной частью дальнейших исследований было доказательство того, что все наблюдаемые эффекты CpdA являлись GR-зависимыми. Для этого мы провели трансдукцию в клетки линий CEM, K562, Granta и NCEB лентивирусных конструкций, экспрессирующих короткую шпилечную РНК к GR (shGR). В полученных трансформированных клетках было продемонстрировано подавление экспрессии GR. И наоборот, в случае клеток Jeko с изначально низкой экспрессией GR мы получили линию трансформированных клеток, экспрессирующих экзогенный GR (рис. 4).

Рис.4. Уровень экспрессии GR после трансдукции в клетки лентивирусных векторов. Уровень экспрессии GR в клетках СЕМ, K562, NCEB и Granta после трансдукции shGR - и контрольным вектора, не содержащими shGR, и в клетках Jeko после трансдукции вектора для экспрессии экзогенного GR и контрольным вектором определяли методом иммуноблоттинга
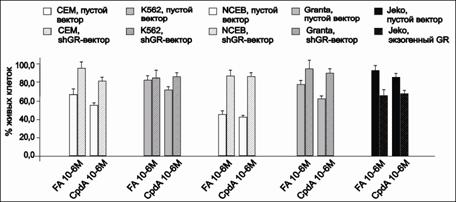
Эксперименты по определению антипролиферативного эффекта FA на исследуемые клетки выявили четкую корреляцию между содержанием рецептора в клетках и цитостатическим эффектом GC. Наиболее сильный антипролиферативный эффект после обработки FA мы наблюдали в линиях СЕМ и NCEB (рис. 5).
|
|
Рис.5. Антипролиферативный эффект FA на трансформированные клетки CEM, K562, NCEB, Granta и Jeko с измененной экспрессией GR. Количество живых клеток представлено в % от контроля, обработанного диметилсульфоксидом (ДМСО) |
Таким образом, нами была разработана модельная система гемобластозов человека для изучения эффектов CpdA в сравнении с GC, а именно, продемонстрирована различная функциональная активность структурно неизмененного GR и получены пары клеточных линий, отличающихся лишь уровнем экспрессии рецептора. Поскольку GR представляет собой как афферентное, так и эфферентное звено целого ряда сигнальных путей в клетке, данные о его структуре и функциональной активности важны не только для выполнения представленного исследования, но и могут быть использованы при анализе различных новых препаратов и подходов к терапии гемобластозов.
2. Оценка генотоксичности CpdA
Для полной характеристики препарата, рассматриваемого в качестве потенциального терапевтического средства, необходим анализ его генотоксичности – способности вызывать повреждения в структуре ДНК. В связи с этим мы оценили генотоксичность СpdA двумя различными методами. Первый метод, бактериальный тест на частоту реверсных мутаций позволял охарактеризовать способность CpdA вызывать мутации по типу сдвига рамки считывания и замены пар оснований. Второй метод, определение влияния вещества на активность топоизомеразы I, оценивал способность СpdA индуцировать соматическую рекомбинацию и, как следствие, вызывать потерю гетерозиготности.
Тест Эймса, наиболее широко распространенный метод определения мутагенной активности соединений, позволяет выявлять как мутагены прямого действия, так и латентные их формы (промутагены) путем активации последних микросомными монооксигеназами из печени крыс. Эксперименты по индукции реверсий у сальмонеллы в основной модификации теста Эймса на штаммах ТА98 и ТА100 в диапазоне концентраций CpdA от 10-9М до 10-4М показали, что данное соединение не обладает мутагенной активностью, в то время как позитивные контроли для штамма ТА98, 2-аминофлуорен и 2,7-диамино-4,9-диокси-5,10-диоксо-4,5,9,10-тетрагидро-4,9-диазапирен (ДДДТДП), вызывали увеличение числа колоний в 46-48 раз, контроли для штамма ТА100, бенз(а)пирен и N-метил-N-нитро-N-нитрозогуанидин, - в 10-20 раз.
Рекомбинационные события в геноме должны быть связаны с нарушениями правильной работы целого ряда ферментов и образованием однонитевых разрывов. В частности, повышение рекомбинационных событий наблюдается под действием красителей Хехст 33258 и 33342, а также Камптотецина, ингибирующих активность топоизомеразы I. Для того, чтобы судить о возможном влиянии CpdA на процесс рекомбинации вышеупомянутым способом, мы оценили его влияние на активность топоизомеразы I. Определение активности данного фермента в присутствии CpdA проводили в широком диапазоне концентраций (1М). Эффективность работы теста на ингибирование активности топоизомеразы I оценивали в сравнении с действием Камптотецина. Как видно из рис. 6, СpdA не нарушает работу топоизомеразы I, и его применение не должно вызывать появление одноцепочечных разрывов в ДНК и, соответственно, повышение частоты рекомбинационных событий.
|
|
Рис.6. Влияние CpdA на активность эукариотической топоизомеразы I. 1 – для контроля влияния условий реакции инкубацию проводили без добавления соединения и топоизомеразы, 2 - для контроля корректной работы топоизомеразы I инкубацию проводили в отсутствие соединения, 3 - в качестве известного ингибитора топоизомеразы I использовали препарат Камптотецин в концентрации 10-6М, 4-9 - для определения способности модулировать активность топоизомеразы I CpdA инкубировали с суперскрученной плазмидной ДНК рНOT в концентрациях М в присутствии топоизомеразы I |

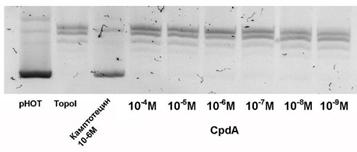
Таким образом, мы продемонстрировали, что CpdA не является мутагенным и рекомбиногенным соединением по результатам теста Эймса и теста на ингибирование топоизомеразы I соответственно.
3. Анализ биологических эффектов CpdA на клетки лейкозов и лимфом
3.1. Антипролиферативный и проапоптотический эффекты CpdA
GC обладают специфической цитотоксичностью в отношении клеток системы кроветворения. Такая тканеспецифичность связана с различиями в активности определенных сигнальных путей и другими особенностями клеток крови. При терапии солидных опухолей GC применяются, в основном, как противовоспалительное средство, однако выраженный противоопухолевый эффект отсутствует. Более того, было показано, что GC увеличивают метастатический потенциал солидных опухолей за счет снижения адгезионных свойств клетки. Однако при терапии гемобластозов увеличение способности клеток к адгезии приводит к рецидивам [Mahlknecht and Schonbein, 2008], и снижение адгезии бластных клеток при лейкозах и лимфомах (в том числе с помощью GC) способствует благоприятному исходу лечения.
Поскольку применение GC в химиотерапии гемобластозов также вызывает быстрое появление целого ряда очень тяжелых побочных эффектов по механизмам транс-активации, поиск препаратов, вызывающих избирательную GR-опосредованную транс-репрессию представляется перспективным современным направлением таргетной терапии.
CpdA является новым нестероидным лигандом GR, свойства которого были изучены на клетках рака предстательной железы PC3 и LNCaP. Однако экстраполировать эффекты CpdA, продемонстрированные на монослойных культурах эпителиальных опухолевых клеток на клеточные линии гемобластозов не представляется возможным из-за тканеспецифических особенностей клеток. В представленной работе было впервые изучено влияние CpdA на экспрессию и функциональную активность рецептора в клетках гематобластозов лимфоидного и миелоидного ряда.
Как уже было отмечено, обработка клеток GC приводит по механизму отрицательной обратной связи к уменьшению количества рецептора. Это в свою очередь уменьшает чувствительность клеток к обработке GC [de Bosscher et al, 2010]. В данной работе мы продемонстрировали, что длительная инкубация клеток всех исследуемых линий с CpdA не вызывает, в отличие от FA, уменьшения экспрессии GR (рис. 7). Эти данные согласуются с результатами исследования де Босшер и соавт., которые показали на мышиной модели ревматоидного артрита, что CpdA не влияет на экспрессию GR [de Bosscher et al, 2010], Различия в эффектах СpdA и GC на экспрессию GR должны обеспечить более медленное развитие резистентности при применении CpdA, и указывает на важное свойство CpdA сохранять свои противовоспалительные, цитостатические и проапоптотические эффекты даже в случае пролонгированного применения.

Рис.7. Влияние CpdA на уровень экспрессии GR в клетках СЕМ, К562, NCEB, Granta и Jeko. Уровень экспрессии GR определяли методом иммуноблоттинга
Нам удалось показать, что так же, как и в клетках рака предстательной железы [Yemelyanov 2007, Yemelyanov 2008] CpdA обладает низкой способностью вызывать перемещение GR в ядро клеток гемобластозов (рис. 8).
Необходимо отметить, что основным процессом, происходящим в ядре при действии глюкокортикоидов, является связывание гомодимера активированного рецептора с ДНК и запуск транскрипции пропролиферативных, антиапоптотических и воспалительных генов, то есть процесс транс-активации, обуславливающий развитие побочных эффектов. Белок-белковые взаимодействия GR с транскрипционными факторами, приводящие к негативной регуляции их транскрипционной активности, происходят как в ядре, так и в цитоплазме.
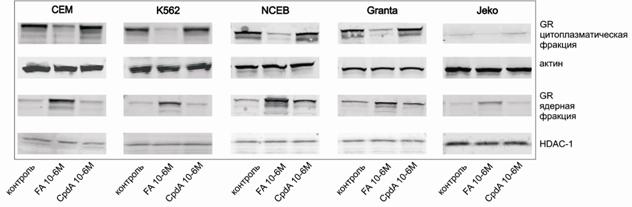
Рис.8. Влияние CpdA на транслокацию GR в ядро в клетках СЕМ, К562, NCEB, Granta и Jeko. Уровень экспрессии GR определяли методом иммуноблоттинга
В отличие от GC, CpdA не вызывает переноса GR в ядро, что делает невозможным запуск транс-активационных механизмов, так как их индукция может происходить только в ядре. Несмотря на это, под действием CpdA мы получили выраженный цитостатический эффект, сопоставимый с эффектом глюкокортикоида. Возможно, это связано с тем, что большая часть антипролиферативных эффектов CpdA вызывается активированным GR, находящимся в цитоплазме по транс-репрессорному механизму. Аналогичное предположение может быть сделано также из данных других исследователей, полученных на культурах клеток простаты [Yemelyanov et al, 2008]
Антипролиферативные эффекты FA и CpdA на клетки всех исследуемых линий соответствовали уровню экспрессии GR. При сравнении эффектов FA на парах клеточных линий NCEB и NCEB-shGR, CEM и CEM-shGR, Jeko и Jeko-GR соответственно различающихся лишь уровнем экспрессии GR, была продемонстрирована четкая взаимосвязь между количеством рецептора и снижением количества жизнеспособных клеток (рис. 9). При исследовании аналогичного эффекта CpdA также было показано, что в клетках с высоким содержанием GR данное соединение проявляет значительное цитостатическое действие, в отличие от линий трансформированных клеток, где экспрессия GR была подавлена.
|
|
Рис. 9. Влияние FA и CpdA на кинетику пролиферации клеток CEM, K562, NCEB, Granta и Jeko. Количество живых клеток представлено в процентах от контроля, обработанного ДМСО |
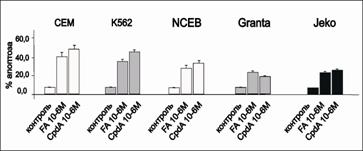
С помощью проточной цитофлуориметрии было продемонстрировано, что в клетках всех исследуемых линий инкубация с CpdA или с FA вызывала запуск апоптоза Проапоптотический эффект данных соединений на клетки лимфом и лейкозов в свою очередь соответствал уровню экспрессии GR: наибольшее количество клеток в пре-G1-фазе мы наблюдали в случае клеточных линий СЕМ, K562 и NCEB (рис. 10).
|
|
Рис.10. Уровень апоптоза в клетках CEM, K562, NCEB, Granta и Jeko после обработки FA и CpdA. Количество апоптотических клеток в процентах от контроля, обработанного растворителем, определяли методом проточной цитофлуориметрии с использованием окраски пропидий иодидом |
Таким образом, было продемонстрировано, что CpdA обладает антипролиферативным действием на клетки лейкозов CEM и K562 и лимфом NCEB, Granta и Jekо. Была показана зависимость данного эффекта от уровня экспрессии рецептора. Кроме того, было установлено, что CpdA не вызывает уменьшения экспрессии GR и, следовательно, развитие резистентности к СpdA должно происходить менее активно, чем при введении GC.
Поскольку чувствительность клеток к GC коррелирует с уровнем экспрессии рецептора, для дальнейших исследований мы выбрали клеточные линии NCEB и СЕМ, наиболее чувствительные к действию FA и CpdA. На данных клеточных линиях мы проводили оценку транс-репрессорного и транс-активационного потенциала CpdA, а также исследование его совместных эффектов с Бортезомибом.
3.2. Оценка транс-активационного и транс-репрессорного потенциала CpdA
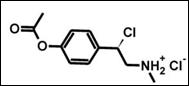
Суммарный эффект глюкокортикоидов на клетки или ткани определяется тем, в какой степени реализуется активация транс-репрессорных и транс-активационных механизмов в клетке. В случае активации GR стандартными GC эти процессы представлены примерно в равной степени. Модификация стероидов и синтез новых нестероидных лигандов GR позволили увеличить долю транс-репрессии. Структура CpdA значительно отличается от большинства известных антагонистов GR, а также от его селективных агонистов [Schacke et al, 2002, Schake et al, 2007, Yemelyanov et al, 2008, Swart et al, 2003, Honer et al, 2003]. CpdA представляет собой низкомолекулярное соединение, содержащее в своей структуре только одно бензольное кольцо (рис.11).
|
|
Рис.11. Химическая структура CpdA |
Ранее было показано, что CpdA может взаимодействовать с лиганд-связывающим доменом GR, конкурируя за связывание с глюкокортикоидом Дексаметазоном [de Bosscher et al, 2005, Yemeyanov et al, 2008]. В отличие от Дексаметазона, CpdA не вызывает фосфорилирование GR по остатку серина Ser211, что является необходимым условием для транслокации рецептора в ядро, образования гомодимера и запуска транс-активационного механизма [de Bosscher et al, 2005]. Емельяновым и соавт. в 2008 г было показано, что данное соединение подавляет связывание GR с ДНК, а также транскрипционную активность рецептора в клетках рака простаты [Yemeyanov et al, 2008].
Запуск транс-активационного механизма GR в клетке происходит при связывании гомодимера рецептора с определенными последовательностями нуклеотидов в ДНК – респонсивными элементами GC (GRE). Это приводит к ацетилированию гистонов, уменьшению плотности хроматина, что увеличивает доступ к ДНК фермента РНК-полимеразы II, катализирующего синтез мРНК [Adcock, 2000]. Таким образом, в клетке активируются процессы транскрипции генов и образования белков, обладающих про-пролиферативным и антиапоптотическим действием.
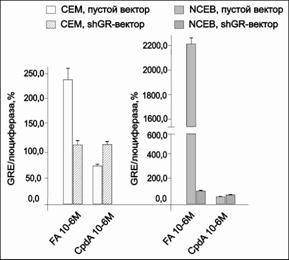
Для оценки влияния CpdA на GR-зависимую транс-активацию в лейкозных клетках СЕМ и клетках лимфомы NCEB были получены клетки данных линий, экспрессирующие ген люциферазы под контролем промотора с множественными GRE. Было обнаружено, что данное соединение не способно индуцировать транс-активационный механизм, в то время как глюкокортикоид FA сильно увеличивал транс-активацию в клетках СЕМ и NCEB. В отличие от FA, CpdA обладал меньшей способностью GR вызывать транс-активацию (рис. 12).
|
|
Рис.12. Влияние CpdA и FA на транскрипционную активность GR в клетках СЕМ и NCEB. Клетки СЕМ и NCEB после инфекции лентивирусным вектором pGF1-GRE инкубировали в течение 8 ч с FA или CpdA, затем определяли активность люциферазы. Данные по активности люциферазы представлены в процентах от контроля, обработанного ДМСО |
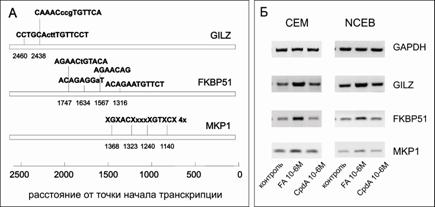
Кроме того, был использован еще один подход анализа транс-активационного потенциала CpdA: был проведен анализ экспрессии ряда генов, которые содержат в своих промоторах GRE, и экспрессия которых увеличивается после обработки клеток GC (рис. 13А). Мы наблюдали этот эффект для генов FKBP51, MKP-1 и GILZ после воздействия FA. Однако, в отличие от FA, CpdA не вызывает увеличение их экспрессии (рис. 13Б). Помимо того, что данные гены являются дополнительными маркерами транс-активационной активности GR, каждый из них обладает собственной функциональной активностью, которая вносит определенный вклад в развитие тех или иных эффектов GC. Ген FKBP51 кодирует белок, входящий в состав шаперонового комплекса GR в цитоплазме. Он принадлежит к семейству иммунофилинов, обладающих способностью связывать FK506 - иммунодепрессант, который блокирует пролиферацию Т-клеток посредством подавления экспрессии отдельных цитокинов [Li et al, 2011].
|
|
Рис.13. Влияние FA и CpdA на экспрессию генов GILZ, FKBP51 и МКР-1. А – схематичное изображение сайтов связывания GR в промоторной области FKBP51, MKP1 и GILZ. Б – Анализ экспрессии FKBP51, MKP1 и GILZ методом ОТ-ПЦР. Количество ПЦР-продуктов оценивали и нормализовали по количеству ПЦР-продукта глицеральдегид-3-фосфатдегидрогеназы (GAPDH) |
По данным литературы, роль FKBP51 при онкологических заболеваниях весьма спорна: в зависимости от гистогенеза опухоли, может наблюдаться как гиперэкспрессия данного гена, так и ее подавление. По данным базы данных Oncomine, содержащей информацию о профилях экспрессии более чем 3200 генов, наблюдается гиперэкспрессия FKBP51 при опухолях мозга, предстательной железы, меланоме и лимфомах [Li et al, 2011, Pei et al, 2009, https://www. oncomine. org]. Было отмечено, что увеличение экспрессии FKBP51 может уменьшать чувствительность клеток острого лимфомбластного лейкоза к антрациклиновым антибиотикам посредством активации IKK-киназ и запуска NF-kB-зависимого сигнального пути [Romano et al, 2004]. Также были получены данные о резистентности к глюкокортикоидам у обезьян саймири, связанной с высоким уровнем FKBP51 у этих животных [Denny et al, 2000, Reynolds et al, 1999].
Ген MKP1 (DUSP-1) соответствует фосфатазе, которая специфически инактивирует МАР-киназы через одновременное дефосфорилирование остатков треонина и тирозина [Wu et al, 2007]. При онкологических заболеваниях экспрессия этого белка может изменяться различным образом в зависимости от заболевания и условий его протекания. В литературе описаны данные о резистентности пациентов с раком яичников, молочной железы и легкого к химиотерапии с гиперэкспрессией МКР1 [Wang etal, 2009, Cortes-Sempere et al, 2009, Haagenson et al, 2010]. В исследованиях Мелхем и соавт. было показано, что применение GC влечет за собой гиперэкспрессию данного гена и является причиной снижения эффективности лечения [Melhem A et al, 2009].
Ген GILZ являлся первым геном, в котором были найдены респонсивные элементы глюкокортикоидов [D’Adamio et al. 1997]. Среди всех белков, регулируемых GR, GILZ является одним из наиболее перспективных потенциальных медиаторов иммуносупрессивного и противовоспалительного действия глюкокортикоидов. Однако в литературе также описано антиапоптотическое действие гиперэкспрессированного GILZ в Т-клетках [Ayroldi and Riccardi, 2009]. Более того, было показано, что при терапии глюкокортикоидами увеличение количества GILZ в остеокластах стимулирует их активацию, что в свою очередь ведет к подавлению пролиферации, дифференциации и функциональных возможностей остеобластов, усилению их апоптоза и, как следствие, развитию стероидного остепороза [Lekva et al, 2010].
Таким образом, увеличение экспрессии всех рассмотренных выше генов связано либо с развитием резистентности (FKBP51 и МКР-1), либо с развитием побочных эффектов (GILZ) при терапии стероидами. Мы показали, что CpdA не вызывал увеличения экспрессии данных генов, и, следовательно, можно ожидать, что его применение не будет приводить к развитию резистентности и уменьшит риск возникновения осложнений при лечении. Это характеризует CpdA как перспективный заместитель GC в лечении гематологических новообразований.
GR-зависимая транс-репрессия играет ведущую роль в противоопухолевом эффекте глюкокортикоидов [Chebotaev et al, 2007, Yemelyanov et al, 2007, Yemelyanov et al, 2008]. На клетках ряда монослойных культур ранее было показано, что CpdA обладает способностью запускать GR-опосредованную транс-репрессию, хорошо воспроизводя действие глюкокортикоидов. Так, на клетках рака простаты было показано, что CpdA, также как и глюкокортикоид FA, ингибирует функциональную активность ряда транскрипционных факторов, включая NF-kB, AP-1, Ets-1, Elk-1, SRF, ATF/CRE и NFATc [Yemelyanov et al, 2008]. Де Босшер и соавт. было продемонстрировано, что CpdA подавляет экспрессию NF-kB-зависимых генов в эмбриональных клетках почки [de Bosscher et al, 2005]. В нашем исследовании при анализе влияния FA и CpdA на активность всех перечисленных выше транскрипционных факторов на клетках NCEB было обнаружено, что ингибирующие эффекты CpdA и FA наиболее ярко выражены в отношении транскрипционных факторов NF-kB и АР-1.
NF-kB, получивший свое название в связи с участием в регуляции экспрессии гена легкой цепи иммуноглобулина κ в В-клетках, представляет собой гетеродимер, состоящий из субъединиц размером в 50 кДа (р50, NFKB1) и 65 кДа (p65, RelA) [Senand and Baltimore, 1986, Lenardo et al, 1989, Scheinman et al, 1995]. Субъединицы NF-kB в неактивном состоянии находятся в цитоплазме в комплексе с ингибитором NF-kB, IkB. Активация данного белка и перемещение его в ядро может быть обусловлена различными причинами, в том числе действием митогенов, цитокинов, липополисахаридов, или вирусной инфекцией. NF-kB является важной частью многих сигнальных путей, которые играют критическую роль в быстрой индукции цитокинов и других генов иммунной системы [Scheinman et al, 1995]. Ранее было показано, что GR может физически взаимодействовать с субъединицами NF-kB и блокировать их способность связываться с ДНК и запускать транскрипцию генов. Также свой вклад в подавление активности NF-kB может вносить негативная регуляция этого фактора транскрипции через МАР-киназный сигнальный путь, приводящая к ингибированию IKK-киназ, играющих ведущую роль в активации NF-kB. [Barnes et al, 2009, Hayashi et al, 2004]. Также глюкокортикоиды вызывают повышение экспрессии главного ингибитора NF-kB, белка IkBα, что приводит к его стабилизации и препятствует транслокации NF-kB в ядро [Hermoso and Cidlowski, 2003]. Таким образом, общее действие GC на NF-kB представляет собой суммарный эффект активации и/или ингибирования в результате изменения активности большого количества сигнальных путей.
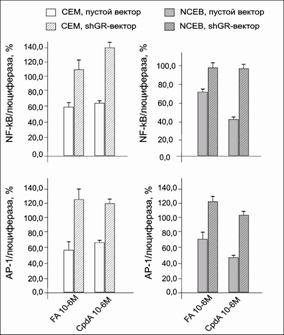
Как было продемонстрировано в нашей работе, GR в клетках лимфомы NCEB и клетках лейкоза СЕМ также способен ингибировать активность NF-kB как при обработке CpdA, так и при обработке FA, причем эффект CpdA был несколько сильнее, чем эффект глюкокортикоида (рис. 14). Полученные в этой работе результаты согласуются с данными, полученными ранее на клетках рака предстательной железы [Yemelyanov et al, 2007, Yemelyanov et al, 2008, de Bosscher et al, 2005].
|
|
Рис.14. Влияние CpdA и FA на транскрипционную активность NF-kB и AP-1 в клетках СЕМ и NCEB. Клетки СЕМ и NCEB после инфекции лентивирусным вектором pGF1-NF-kB и АР-1 инкубировали в течение 8 ч с FA или CpdA, затем определяли активность люциферазы. Данные по изменению активности люциферазы представлены в процентах от контроля, обработанного ДМСО |
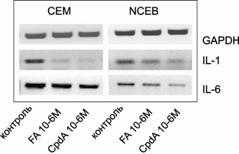
Активность NF-kB после обработки FA и CpdA также исследовали с помощью оценки экспрессии хорошо известных NF-kB-зависимых генов, – цитокинов IL-1 и IL-6, которые стимулируют дифференцировку и пролиферацию Т - и В-клеток и являются медиаторами острого и хронического воспаления [Oppenheim, 1985, Braquet et al, 1989, Kita et al., 1992, Tripathi and Aggarwal, 2006]. Экспрессия этих генов падала, что свидетельствовало о подавлении активности NF-kB (рис. 15).
|
|
Рис.15. Влияние FA и CpdA на экспрессию генов интерлейкина-1 (IL-1) и интерлейкина-6 (IL-6). Оценка уровня экспрессии IL-1 и IL-6 проводилась методом ОТ-ПЦР. Количество ПЦР-продуктов оценивали и нормализовали по количеству ПЦР-продукта глицеральдегид-3-фосфатдегидрогеназы (GAPDH) |
Транскрипционный фактор АР-1 был открыт в клетках млекопитающих благодаря его способности специфически связываться с энхансером вируса обезьян SV40 и промоторами позвоночных, которые содержат АР-1-сайт. Как известно, АР-1 представляет собой гетеродимер, состоящий из белков Fos - (c-Fos, FosB, Fra-1 и Fra-2) и Jun-семейств (c-Jun, JunB и JunD) [Lee et al., 1987, Chen et al, 1998]. АР-1, как и NF-kB, является фактором транскрипции и участвует в поддержании базального уровня экспрессии многих генов. Он является одной из главных мишеней для соединений, вызывающих клеточную пролиферацию или дифференцировку. GR способен ингибировать активность АР-1 либо за счет белок-белкового взаимодействия (собственно транс-репрессии), либо подавлять его транскрипцию, связываясь с негативными GRE в промоторе генов субъединиц АР-1 [Nissen and Yamamoto, 2000, Herrlich et al, 2001]. Также GR, активированный глюкокортикоидами, может взаимодействовать с МАР-киназами и ингибировать их активность, что в свою очередь приводит к подавлению АР-1 [Milleretal, 2005, Gossye et al, 2009]. Кроме того, GR может связываться с GRE в промоторной области генов Jun или Fos, представленных там в большом количестве, и запускать, таким образом, синтез AP-I. В то же время при изучении CpdA было показано, что данное соединение практически не влияет на активность МАР-киназ [Gossye et al, 2009] и образованный им комплекс с GR не способен связываться с ДНК. Таким образом, преобладающим механизмом в случае эффекта СpdA на активность AP-I является непосредственно транс-репрессия.
Как было показано в представленной работе, FA и CpdA в обеих клеточных линиях СЕМ и NCEB подавляют активность транскрипционного фактора АР-1 не менее, чем на 27-30% (рис. 14).
Суммируя полученные на данном этапе работы результаты, можно заключить, что CpdA представляет собой селективный модулятор GR, способный избирательно запускать процесс транс-репрессии, не индуцируя транс-активацию.
4. Анализ эффектов совместного применения CpdA и протеасомного ингибитора Бортезомиба
Комбинированная химиотерапия гемобластозов позволяет получить наилучший терапевтический результат. Наиболее эффективные протоколы, помимо стандартных препаратов-цитостатиков, могут включать фотодинамическую терапию, современные таргетные химиопрепараты и иммунотерапию [Ravandi et al, 2003, Ihle et al, 1998, Scheijen et al, 2002, Ravandi et al, 2003, Fisher et al, 2004, Yu et al, 2003, Cusack et al, 2001, Elliot et al, 2003]. Среди таргетных препаратов одним из самых эффективных считается Бортезомиб (BZ), ингибитор протеасом, введенный в клиническую практику в гг и успешно применяющийся в терапии лимфом, миелом и лейкозов [Orlowski et al, 2002, McConkey and Zhu, 2008, Romaguera et al, 2010, Messinger et al, 2010]. В ряде исследований было продемонстрировано, что пациенты со множественной миеломой и с лимфомой мантийной зоны, а также рядом других гемобластозов, более чувствительны к BZ, и он более специфично оказывает цитотоксическое действие на опухолевые, но не на нормальные клетки [Richardson et al, 2005, McKonkey and Zhu, 2008]. Более того, для данного препарата было показано синергическое действие в комбинации с GC при терапии гемобластозов различного происхождения [Horton et al, 2006]. Данный эффект может быть объяснен рядом причин. BZ подавляет протеасомное расщепление GR, вызывая его накопление в клетке и усиливая его функцию [Elliott et al, 2003]. Глюкокортикоиды, в свою очередь, усиливают экспрессию генов, вовлеченных в сигнальные пути, запускаемые при подавлении активности протеасомы, в частности, генов, принимающих участие в развитии ЭПР-стресса, таких как шапероны ЭПР стресса Grp78 (BiP) и Grp94 [Kraus et al, 2008, Hernandez-Espinosa et al, 2009] и др. Более того, в литературе описана способность как глюкокортикоидов, так и протеасомных ингибиторов подавлять активность NF-kB и антиапоптотических белков из семейства Bcl-2 [McConkey and Zhu 2008, Cusack et al, 2001].
Поскольку CpdA, с одной стороны, является аналогом GC, его действие является GR-зависимым, а с другой стороны, он селективно направлено на запуск транс-репрессии, исследование эффектов от его совместного применения с BZ представлялось нам перспективным. Протеасомы в клетке выполняют функцию элиминирования поврежденных белков, а также отвечают за модулирование активности многих регуляторных белков, которые влияют на воспалительные процессы, клеточную пролиферацию и дифференцировку [Elliott et al, 2003]. Поскольку GR подвергается протеасомной деградации, то, по нашему предположению, BZ, ингибируя работу протеасом, вызывает стабилизацию и дальнейшее накопление GR в клетке. Поскольку CpdA избирательно запускает процесс транс-репрессии, то аккумулированный с помощью BZ рецептор активирует только этот механизм, и в данном случае наблюдаются синергические антипролиферативные и проапоптотические эффекты на клетки лейкозов и лимфом.
Нами был проведен ряд экспериментов по определению оптимальных концентраций CpdA, BZ и FA. В данных экспериментах был обнаружен синергический антипролиферативный эффект на клетки обеих линий при обработке их BZ в концентрации 3×10-8М для СЕМ и 3×10-9М для NCEB, и CpdA в концентрации 3×10-6М для СЕМ и 10-6М для NCEB (рис. 16).
|
|
Рис.16. Влияние FA, CpdA и BZ на кинетику пролиферации клеток CEM и NCEB. Количество живых клеток представлено в процентах от контроля, обработанного ДМСО |
|
|
|
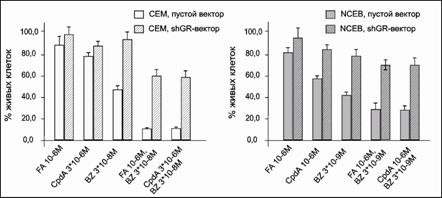
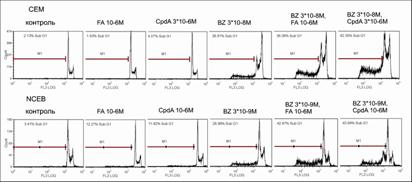
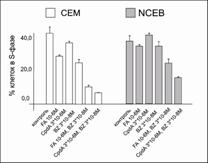
Рис.17. Уровень апоптоза в клетках CEM и NCEB после обработки FA, CpdA и BZ. Количество апоптотических клеток в % от контроля определяли методом проточной цитофлуориметрии с использованием окраски пропидий иодидом
С помощью проточной цитофлуориметрии было показано также, что и CpdA, и FA оказывают потенцирующее действие на проапоптотические эффекты BZ при обработке клеток СЕМ и NCEB в данных концентрациях. Было продемонстрировано увеличение числа апоптотических клеток, которое мы определяли как пре-G1-пик на ДНК-гистограмме. Одновременно с этим наблюдалось уменьшение количества клеток в фазе синтеза, о чем судили по изменению S-пика (рис. 17). Таким образом, было продемонстрировано синергическое антипролиферативное действие CpdA и BZ, обусловленное развитием апоптоза.
Поскольку, согласно полученным результатам, эффект не только лигандов GR, но и BZ коррелировал с количеством рецептора в клетке, это дает основания предполагать, что значительная часть эффектов, проявляемых ингибитором протеасом, реализуются через GR-зависимые сигнальные пути. В литературе описано влияние на GR МАР-киназ, сигнальных путей mTOR и PI3K/Akt [Miller et al, 2005, Miller et al, 2007]. BZ также способен влиять на данные сигнальные каскады, активируя тем самым GR. Например, Миллер и соавт. было продемонстрировано, что активация р38 МАР-киназы способствует сайт-специфическому фосфорилированию GR по остатку серина Ser211 [Miller et al, 2005]. В то же время описана индукция р38-опосредованного пути передачи сигнала при использовании BZ [Wang et al, 2011]. Таким образом, BZ может усиливать активацию GR через МАР-киназный сигнальный путь.
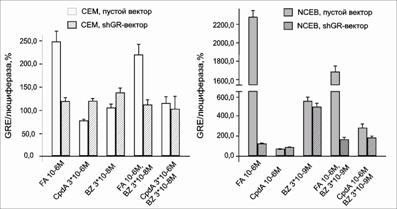
|
|
Рис.18. Влияние CpdA, FA и BZ на транскрипционную активность GR в клетках СЕМ и NCEB. Клетки СЕМ и NCEB после инфекции лентивирусным вектором pGF1-GRE инкубировали в течение 8 ч с FA или CpdA, затем определяли активность люциферазы. Данные по изменению активности люциферазы представлены в процентах от контроля, обработанного ДМСО |
Далее было проанализировано совместное действие CpdA и BZ, FA и BZ на основные механизмы работы GR – транс-активацию и транс-репрессию. Для этого были использованы трансформированные клетки СЕМ и NCEB, несущие ген люциферазы светлячка под контролем промоторов, содержащих либо множественные GRE (для оценки транс-активации), либо сайты связывания NF-kB и АР-1 (для оценки транс-репрессии).
BZ не внес существенного вклада в запуск процесса транс-активации в клетках СЕМ. В клетках NCEB ингибитор протеасом вызывал увеличение активности репортерного гена в 5 раз по сравнению с контролем (рис. 18). Однако при совместной обработке клеток NCEB BZ и FA наблюдалось потенцирующее действие ингибитора протеасом на подавление транс-активационного процесса, а при комбинированном применении CpdA и BZ на данные клетки CpdA способствовал снижению активности люциферазы, индуцированной BZ.
Влияние BZ на GR-опосредованную транс-репрессию определяли по эффектам данного соединения на транскрипционную активность NF-kB и АР-1.
Протеасомы вносят вклад в активацию NF-kB посредством двух основных механизмов, один из которых заключается в протеасомной деградации физиологического ингибитора NF-kB, IkB. Это ведет к чрезмерной активации NF-kB, его ядерной транслокации и запуску транскрипции провоспалительных генов. Второй механизм представляет собой протеолитическое расщепление р100, который является предшественником р52, играющего центральную роль в запуске неканонического пути активации NF-kB [Dejardin et al, 2006]. Было продемонстрировано, что ингибиторы протеасом, в том числе и BZ, способны блокировать активацию NF-kB, наблюдающуюся при применении определенных препаратов-цитостатиков или ионизирующей радиации [Adams et al, 2004, Cusack et al, 2001, Garber et al, 2002, Russo et al, 2001]. Рассмотренные выше пути активации GR при действии BZ через сигнальные пути МАР-киназ, mTOR и другие, дают основание предположить, что механизм подавления транскрипционной активности NF-kB может также осуществляться посредством GR-опосредованной транс-репрессии.
Было показано, что при инкубации клеток обеих линий только с BZ активность NF-kB падала незначительно. Однако при обработке клеток комбинациями FA с BZ и CpdA с BZ наблюдалось потенцирующее действие протеасомного ингибитора на эффекты обоих соединений (рис. 19, 20). Более того, при инкубации с данными соединениями клеток СЕМ-shGR и NCEB-shGR было установлено, что такие клетки гораздо менее чувствительны не только к действию непосредственных лигандов GR, CpdA и FA, но и BZ (рис. 19, 20). Таким образом, механизм совместного действия BZ и CpdA, а также BZ и глюкокортикоидов заключается в подавлении активности NF-kB и в большой степени является GR-зависимым.
Полученные нами данные хорошо согласуются с результатами изучения эффектов BZ в комбинации с GC при лечении при лечении множественной миеломы, лимфом и лейкозов [McKonkey and Zhu, 2008] и демонстрируют перспективность данного препарата перед другими химиотерапевтическими агентами, активирующими NF-kB, что ограничивает их цитотоксические эффекты [McKonkey and Zhu, 2003, Wang et al, 1996, Wu et al, 1996].
Транскрипционный фактор АР-1 является еще одним белком, регулируемым GR, чья активность может быть повышена BZ через сигнальные пути МАР-киназ, mTOR, PI3K/Akt. Как и в случае с NF-kB, мы использовали модельную систему клеток СЕМ и NCEB, с пониженной или нормальной экспрессией GR, и в то же время экспрессирующих люциферазу светлячка под контролем промотора АР-1. Мы наблюдали GR-зависимый потенцирующий эффект BZна подавление активности АР-1 как FA, так и CpdA. Наиболее четко этот эффект был выражен в клетках СЕМ.
Таким образом, при использовании CpdA и BZ в определенных концентрациях при их комбинированном воздействии было продемонстрировано потенцирующее действие BZ на эффект CpdA в клетках СЕМ и NCEB. Обработка клеток комбинациями CpdA (или FA) и BZ, вызывала значительное подавление пролиферации клеток, а также влияла на GR-зависимые процессы транс-активации и транс-репрессии.
|
|
Рис.19.Влияние CpdA, FA и BZ на транскрипционную активность NF-kB и AP-1 в клетках СЕМ. Клетки СЕМ после инфекции лентивирусным вектором pGF1-NF-kB и АР-1 инкубировали в течение 8 ч с FA или CpdA, затем определяли активность люциферазы. Данные по изменению активности люциферазы представлены в процентах от контроля, обработанного ДМСО |
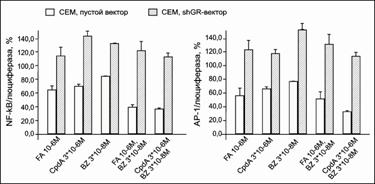
|
|
Рис. 20. Влияние CpdA, FA и BZ на транскрипционную активность NF-kB и AP-1 в клетках NCEB. Клетки NCEB после инфекции лентивирусным вектором pGF1-NF-kB и АР-1 инкубировали в течение 8 ч с FA или CpdA, затем определяли активность люциферазы. Данные по изменению активности люциферазы представлены в процентах от контроля, обработанного ДМСО |

ЗАКЛЮЧЕНИЕ
В представленном исследовании проведено изучение нового нестероидного лиганда GR, 2-(4-ацетоксифенил)-2-хлор-N-метилэтиламмоний хлорида, или CpdA. Исследования проводились на двух клеточных линиях лейкозов СЕМ и К562, и трех линиях лимфом NCEB, Granta и Jeko.
На первом этапе была разработана модельная система гемобластозов человека для изучения эффектов лигандов GR. Мы показали, что клеточные линии лейкозов СЕМ и К562, и лимфом NCEB, Granta и Jeko, не несут в гене GR мутаций, способных нарушать такие его функции, как связывание с лигандом и перемещение в ядро. Во всех пяти линиях клеток большую часть экспрессируемого белка составляла изоформа GR-α, которая отвечает за все основные функции рецептора. Для анализа роли GR в наблюдаемых эффектах были получены трансформированные клетки с измененным статусом рецептора; было проведено подавление экспрессии GR с помощью shRNA в клетках СЕМ, К562, NCEB и Granta, и введение экзогенного GR в клетках Jeko.
На данной модельной системе мы впервые продемонстрировали GR-зависимые антипролиферативный и проапоптотический эффекты CpdA на клетки лейкозов и лимфом, сравнимые с действием GC. Мы показали, что CpdA, в отличие от GC, не вызывает уменьшения экспрессии рецептора, что является одним из механизмов сохранения чувствительности клеток к его действию. Эти данные согласовывались с отсутствием увеличения экспрессии генов FKBP51 и МКР1, что играет существенную роль в развитии резистентности к GC.
Способность CpdA активировать механизм GR-зависимой транс-репрессии также была нами показана в случае транскрипционных факторов NF-kB и АР-1, а также NF-kB и АР-1-зависимых генов IL-1 и IL-6. В то же время было продемонстрировано отсутствие запуска транс-активационного механизма при инкубации клеток с CpdA: данное соединение не вызывало ни увеличения активности репортерного гена, находящегося под GRE-содержащим промотором, ни увеличения экспрессии генов, имеющих в промоторах множественные GRE.
В работе проведена оценка совместного действия CpdA с ингибитором протеасом Бортезомибом на клетки лейкозов и лимфом. BZ является перспективным препаратом в отношении терапии гематологических новообразований. Нами было показано синергическое антипролиферативное и проапоптотическое действие данных соединений на клетки лейкозов и лимфом. В то же время было продемонстрировано, что BZ не подавляет «диссоциированные» свойства CpdA, а наоборот, оказывает потенцирующее действие на ингибирование активности транскрипционных факторов NF-kB и АР-1 при совместной обработке клеток CpdA и BZ.
Таким образом, в данной работе впервые на клетках лейкозов и лимфом были охарактеризованы эффекты нового селективного лиганда GR, CpdA, и продемонстрировано синергическое действие CpdA и Бортезомиба при совместной обработке клеток их определенными концентрациями.
ВЫВОДЫ:
1. Клеточные линии лейкозов СЕМ и К562, и лимфом NCEB, Granta и Jeko, являются адекватной модельной системой для оценки эффектов новых лигандов GR, поскольку они экспрессируют функционально активный рецептор, не несущий в себе полиморфизмов и мутаций.
2. Нестероидный аналог глюкокортикоидов, 2-(4-ацетоксифенил)-2-хлор-N-метилэтиламмоний хлорида, или CpdA, вызывает в клетках лейкозов СЕМ и К562, и лимфом NCEB, Granta и Jeko значительные антипролиферативные и проапоптотические эффекты, сравнимые с действием глюкокортикоидов.
3. CpdA не является генотоксическим агентом: он не индуцирует появление точечных мутаций, и не активирует рекомбинационную репарацию путем ингибирования топоизомеразы I.
4. CpdA по биологическим эффектам на клетки лейкозов и лимфом представляет собой селективный агонист GR: он активирует транс-репрессию NF-kB, АР-1 и связанных с ними генов, но не запускает процесс транс-активации.
5. В отличие от GC, CpdA не вызывает развития резистентности: обработка клеток данным нестероидным лигандом GR не приводит к уменьшению экспрессии рецептора и не вызывает увеличения экспрессии генов FKPB51 и МКР1, ассоциированных с потерей чувствительности к действию GC.
6. При совместном применении ингибитора протеасом Бортезомиба и CpdA наблюдается синергизм антипролиферативного и проапоптотического эффектов данных соединений.
7. Бортезомиб оказывает потенцирующее действие на способность CpdA селективно модулировать активность GR, причем этот эффект зависит от уровня экспрессии рецептора в клетках.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Лесовая, действие нестероидного лиганда глюкокортикоидного рецептора, CpdA, на лейкозные клетки линий СЕМ и К562/, , и др. // Биохимия (Москва)– Т. 76, № 11. – С. .
2. Kirsanov, K. I. SYBR Gold and SYBR Green II are not mutagenic in Ames test/K. I., Kirsanov, E. A. Lesovaya, M. G. Yakubovskaya, G. A. Belitsky // Mutation Research. – 2010. – V. 699. – P. 1–4.
3. Сусова, димерные бисбензимидазолы ингибируют in vitro связывание с ДНК эукариотической ДНК-топоизомеразы I/, , Руис, и др. // Биохимия (Москва). – 2010. – Т. 75, № 6. – С. 781-788.
4. Лесовая, раковых заболеваний при помощи направленной активации комплемента/, // Российский Биотерапевтический журнал. – 2008. – Т. 3. – С. 13-19.
5. Lesovaya, E. A. Selective activation of glucocorticoid receptor transrepression as a new approach for acute leukemia targeted therapy/E. A. Lesovaya, M. G. Yakubovskaya, I. V. Budunova // Spetses Summer School on Nuclear Receptor signaling. – Spetses, Greece, Aug. 23-28, 2009. - P.30
6. Lesovaya, E. A. Anti-cancer effect of Compound A, a novel modulator of the glucocorticoid receptor, in acute leukemia cells/E. A. Lesovaya, K. I. Kirsanov, I. Budunova, M. G. Yakubovskaya // Proceedings of the 101st Annual Meeting of the American Association for Cancer Research. – Washington, DC, USA, Apr 17-21, 2010. - Abstract 214.
7. Lesovaya E. A. Anti-cancer effects of novel non-steroidal glucocorticoid receptor ligand, Compound A, in T-cell leukemia lines/E. A Lesovaya., A. Yemelyanov, K. I. Kirsanov et al // Proceedings of the 102nd Annual Meeting of the American Association for Cancer Research. - Orlando, FL, USA, Apr 2-6, 2011. - Abstract 669.
