
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ГЕРБЕК ЮРИЙ ЭМИЛЬЕВИЧ
ЭФФЕКТ ОТБОРА ПО ПОВЕДЕНИЮ СЕРЫХ КРЫС
НА ЭкспрессиЮ гена рецептора глюкокортикоидов
03.02.07 – генетика
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Новосибирск 2010
Работа выполнена в лаборатории эволюционной генетики Учреждения Российской академии наук Институт цитологии и генетики Сибирского отделения РАН, г. Новосибирск.
Научный руководитель: | кандидат биологических наук, старший научный сотрудник Институт цитологии и генетики СО РАН, г. Новосибирск |
Официальные оппоненты: | доктор биологических наук, профессор Институт цитологии и генетики СО РАН, г. Новосибирск доктор биологических наук, профессор Научный центр клинической и экспериментальной медицины СО РАМН г. Новосибирск |
Ведущее учреждение: | Институт физиологии им. РАН, г. Санкт-Петербург |
Защита диссертации состоится «__» ноября 2010 г. на утреннем заседании диссертационного совета Д 003.011.01 по защите диссертаций на соискание учёной степени доктора наук в Институте цитологии и генетики СО РАН в конференц-зале Института по адресу:
г. Новосибирск, пр-т Ак. Лаврентьева, д. 10, т. (383), , e-mail: *****@
С диссертацией можно ознакомиться в библиотеке Института цитологии и генетики СО РАН.
Автореферат разослан «__» октября 2010 г.
Ученый секретарь
диссертационного совета,
доктор биологических наук
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность проблемы. Стресс – одно из важнейших свойств организма, позволяющее сохранять гомеостаз в экстремальных ситуациях. В то же время длительное действие стрессора может привести к истощению организма, необратимым преобразованиям и смерти (Селье, 1979). Поэтому при продолжительном сохранении изменившихся условий пониженная стресс-реактивность, вероятно, является одним из основных критериев приспособленности (Беляев, Бородин, 1982). В этом ключе, представляет особенный интерес эксперимент по доместикации животных, который проводится в Институте цитологии и генетики СО РАН. По мнению , при историческом одомашнивании решающим фактором являлся отбор по поведению по отношению к человеку, который, по-видимому, включал в себя отбор на стресс-реактивность (Науменко, Беляев, 1981).
Показано, что отбор на доместикацию снижает функциональную активность гипоталамо-гипофизарно-надпочечниковой системы (ГГНС) (Дыгало и др., 1985; Naumenko et al., 1987; Оськина, Плюснина, 2000), являющейся ключевой в развитии стресс-ответа. У серых крыс, селекционируемых на элиминацию и усиление агрессивного поведения по отношению к человеку («ручные» и «агрес-сивные», соответственно), выявлены различия в динамике стресс-реакции, а также в базальном и стресс-индуцированном уровнях глюкокортикоидов (конечного звена ГГНС) в крови. Установлено, что у ручных крыс уровень глюкокортикоидов в плазме крови ниже, чем у агрессивных (Дыгало и др. 1985; Naumenko et al., 1989; Оськина, Плюснина, 2000). В регуляции стресс-ответа ведущее место занимают рецепторы глюкокортикоидов (ГР) гиппокампа. Они участвуют в отрицательной обратной связи ГГНС, купируя стресс-реакцию (Pfaff et al., 2004). Установлено, что у крыс, селекционируемых на контрастное поведение по отношению к человеку, количество ГР в гиппокампе различно (Оськина, Плюснина, 2000).
Наиболее исследованным путём регуляции ГР у крыс, влияющий на ГГНС, является механизм, связанный с альтернативным сплайсингом мРНК ГР (McCormick et al., 2000). Он описан при неонатальных воздействиях, которые вызывают повышение количества ГР в гиппокампе и снижение активности ГГНС (Meaney et al., 1996). Это наводит на мысль о схожести механизмов изменения стресс-реактивности при отборе по поведению и вследствие неонатальных влияний. Показано, что уровень транскрипции гена ГР под контролем промотора альтернативного экзона 17 повышается в гиппокампе под влиянием раннего опыта (Weaver et al., 2004), что связывают с деметилиро-ванием этого промотора. Если же вводить в мозг крысам, подвергнутым таким воздействиям, метионин, то уровень метилирования ДНК повышается, что вызывает снижение экспрессии гена ГР и усиление активность ГГНС (Weaver et al., 2004). Полагают, что подобный механизм задействован и при пренатальных влияниях (Seckl, Holmes, 2007). С другой стороны, содержание крыс во время беременности на малобелковых диетах влияет на уровень мРНК ГР с экзоном 110 в печени их потомков (Lillycrop et al., 2007). Транскрипция под контролем промотора этого экзона составляет более 60% от общей транскрипции гена ГР (McCormick et al., 2000) и регулируется метилированием ДНК (Lillycrop et al., 2007). Однако в пренатальный период развития не было исследовано влияние метионина на экспрессию гена ГР. В то же время известно, что материнская метилобогащённая диета вызывает различные изменения в формировании центральной нервной системы и в профиле метилирования некоторых генов у потомков (Cooney et al., 2002; Van den Veyver, 2002).
Эксперимент по содержанию крыс во время беременности на метилобогащённой диете, проведённый на крысах, селекционируемых на контрастное поведение по отношению к человеку, позволяет оценить влияние метионина в эмбриональный период развития на уровень метилирования промотора гена ГР и экспрессию этого гена во взрослом состоянии. Результаты эксперимента также дают возможность выяснить, являются ли схожими механизмы изменения активности ГГНС при отборе по поведению и перинатальных воздействиях.
Цель и задачи исследования. Целью данной работы является изучение экспрессии гена рецептора глюкокортикоидов у серых крыс, селекционируемых на ручное и агрессивное поведение, и исследование механизмов возможных различий.
Конкретные задачи исследования состояли в следующем:
1. Оценить общую экспрессию гена ГР в гиппокампе и периферических органах у ручных и агрессивных крыс.
2. Исследовать активность периферических звеньев ГГНС и общую экспрессию гена ГР у первого и второго поколений потомков ручных и агрессивных серых крыс, содержавшихся на метилобогащённой диете в период беременности.
3. Проанализировать профиль метилирования промотора экзона 17 гена ГР и количество мРНК ГР с альтернативным экзоном 17 в гиппокампе у потомков крыс, содержащихся на метилобогащенной диете или стандартном питании во время беременности.
4. Определить уровень мРНК ГР с альтернативным экзоном 110 в гиппокампе у крыс, матери которых содержались на стандартном питании во время беременности, а также у первого и второго поколений потомков животных, получавших в период беременности метилсодержащие добавки.
Научная новизна и практическая ценность работы. В данной работе впервые изучено влияние отбора по поведению на экспрессию гена ГР у крыс. Отбор на ручное поведение повышает количество мРНК ГР в гиппокампе, однако не затрагивает мРНК ГР в печени и селезёнке. Впервые исследовано влияние материнской метилобогащённой диеты на активность ГГНС и экспрессию гена ГР у потомков. Материнская диета у животных из ручной популяции приводит к более позднему возвращению уровня кортикостерона в крови к базальным показателям после рестрикционного (лат. стресса и к снижению общего количества мРНК ГР и уровня мРНК ГР с экзоном 110 в гиппокампе, что свидетельствует об ослаблении отрицательной обратной связи ГГНС. В то же время, достоверных влияний пренатальной диеты на динамику стресс-ответа и экспрессию гена ГР у животных из популяции, селекционируемой на усиление агрессивного поведения по отношению к человеку, выявлено не было. Обнаружено отличное от описанного ранее метилирование промотора экзона 17 гена ГР крысы в гиппокампе. Выявлено гипометилирование участка ДНК, считавшегося ранее гиперметилированым у крыс. Данная работа расширяет представления о механизмах изменения активности ГГНС при эволюционных преобразованиях животных вследствие отбора по поведению. Кроме того, освещается вопрос о влиянии пренатальной метилобогащённой диеты на экспрессию гена ГР и о зависимости эффектов диеты от генотипа животных. Полученные данные могут иметь значение в исследованиях патологий раннего развития. Результаты исследования используются в качестве материала для курса лекций «Нейробологические и нейрофизиологические закономерности поведения», читаемого на 4 курсе ФЕН НГУ.
Положения, выносимые на защиту:
1. Отбор серых крыс на доместикационное поведение повышает уровень экспрессии гена ГР в гиппокампе.
2. Выявлены различия в эффекте материнской метилобогащённой диеты на экспрессию гена ГР в гиппокампе у потомков крыс, селекционируемых на элиминацию и усиление агрессивно-оборонительной реакции по отношению к человеку, свидетельствующие о роли метилирования ДНК в эффектах отбора по поведению.
3. Установлен значительный вклад мРНК с экзоном 110 в регуляцию экспрессии гена ГР в гиппокампе и активности ГГНС под влиянием пренатальной метилобогащённой диеты.
4. Выдвинуто предположение, что метилирование промотора экзона 17, на котором основан ранее описанный механизм регуляции экспрессии гена ГР и активности ГГНС, является эктопическим.
Апробация работы. Материалы диссертации обсуждались на XX съезде Физиологического общества им. (Москва, 2007), Международной конференции «Current Evolution Thinking in Biology, Medicine and Sociology International Conference Dedicated to 90 Anniversary of Prof. Dmitry K. Belyaev» (Новосибирск, 2007), IV Международной научной конференции «Факторы экспериментальной эволюции организмов» (Алушта, 2008), VI Сибирском физиологическом съезде (Барнаул, 2008), V съезде Вавиловского общества генетиков и селекционеров (Москва, 2009), международном конгрессе «The 7th International Congress of Neuroendocrinology» (Руан, Франция, 2010) и XXI съезде Физиологического общества им. . Также полученные результаты были представлены и обсуждены на отчётной сессии по программе Президиума РАН «Молекулярная и клеточная биология» (Новосибирск, 2009) и отчётной сессии Института цитологии и генетики СО РАН (Новосибирск, 2010).
Публикации. По материалам диссертации опубликовано 10 работ.
Вклад автора. Основные результаты получены автором самостоятельно. Уровень кортикостерона в крови животных определялся к. б.н. Оськиной и отбор по поведению серых крыс проводились к. б.н.
Структура и объем диссертации. Диссертация содержит следующие разделы: введение, обзор литературы, материалы и методы, результаты исследования, обсуждение, выводы, список цитируемой литературы. Работа изложена на 128 страницах, содержит 17 рисунков, 5 таблиц, список цитируемой литературы включает 241 ссылку.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для экспериментов служили две аутбредные линии серых крыс (Rattus norvegicus), селекционируемых с 70-х годов на элиминацию и усиление агрессивно-оборонительной реакции по отношению к человеку. Животные содержались в стандартных условиях вивария ИЦиГ СО РАН. В опыт были взяты взрослые самцы 64–70 поколений селекции. Также были исследованы дикие серые крысы (самцы), отловленные в окрестностях новосибирского Академгородка и в Новосибирской области.
Матери опытных ручных и агрессивных крыс содержались во время беременности и 5 дней после родов на метилобогащенной диете (Прасолова и др., 2009), а матери контрольных – на стандартном питании. Эксперимент проводили на трехмесячных самцах.
Для оценки активности ГГНС в ответ на стрессорное воздействие исследовали содержание кортикостерона в периферической крови. В качестве стрессорного воздействия использовали ограничение подвижности в течение 30 мин. Пробы крови брали из хвостовой вены до начала эксперимента (базальный уровень) и через разные отрезки времени после окончания стресса. Уровень кортикостерона в плазме крови определяли методом конкурентного белкового связывания (Murphy, 1967; Тинников, Бажан, 1984).
Крыс быстро декапитировали, извлекали печень, селезёнку и головной мозг, от которого отделяли гиппокамп. Ткани замораживали и хранили в азоте. Суммарную РНК, а также ДНК выделяли с использованием «TRI Reagent» («Sigma», США) в соответствии с протоколом фирмы изготовителя. Раствор РНК обрабатывали ДНКазой I («Fermentas», Литва). Затем синтезировали кДНК с помощью рекомбинантной обратной транскриптазы RevertAid M-MuLV («Fermentas», Литва).
Для оценки количества мРНК ГР в образцах использовали метод полуколичественной ОТ-ПЦР (Horikoshi, Sakakibara, 2000). Условия стандарти-зовали путём побора концентрации кДНК и количества циклов ПЦР, при которых реакция находилась в экспоненциальной фазе роста количества продукта, для исследуемого гена и внутреннего контроля. Также использовали метод ОТ-ПЦР в реальном времени (Pfaffl, 2004), который позволяет регистрировать накопление продукта амплификации исследуемого гена и внутреннего контроля в каждом цикле реакции. Для этого использовали комплект реагентов для проведения ПЦР-РВ в присутствии SYBR Green I и референсного красителя ROX (Синтол, Россия) и амплификаторы ABI PRISM 7000 и 7900HT (Applied Biosystems, США).
Для определения метилирования промотора экзона 17 гена ГР использовали метод бисульфитного секвенирования (Clark et al., 1994). После обработки ДНК бисульфитом натрия, при которой все неметилированные цитозины превращались в урацил, проводилась ПЦР с «вложенными» праймерами (Weaver et al., 2006). После чего полученный продукт клонировали в химически-модифицированных бактериях с помощью набора pGEM®-T Easy Vector System II (Promega, США), амплифицировали с использованием универсальных праймеров M13 и секвенировали.
Определение нуклеотидной последовательности проводили согласно протоколу с помощью набора BigDye™ Terminator Ready Reaction Cycle Sequencing Kit (Applied Biosystems, США). Электрофоретический анализ проводился в центре коллективного пользования «Секвенирование ДНК» СО РАН на автоматическом секвенаторе ABI PRISM 3130xl.
Статистический анализ результатов определения содержания кортикостерона в крови проводили с использованием двухфакторного дисперсионного анализа с последующим сравнением межгрупповых различий по критерию Ньюмена-Кейлса, учитывая повторные измерения (ANOVA-MANOVA; Statistica 6.0). Для сравнения результатов определения количества мРНК использовался t-критерий Стьюдента (для ОТ-ПЦР и ОТ-ПЦР в реальном времени) или рандомизированный критерий с двухточечным пере-распределением и алгоритм Тейлора (только для ОТ-ПЦР в реальном времени; REST 2008–2009). При анализе данных по метилированию ДНК применяли U-критерий Манна-Уитни (Statistica 6.0).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Уровень мРНК рецептора глюкокортикоидов у серых крыс, контрастных по поведению
Известно, что у ручных крыс количество ГР в гиппокампе достоверно выше, чем у агрессивных (Оськина, Плюснина, 2000), что, по всей видимости, усиливает отрицательную обратную связь ГГНС и снижает активность системы. Методом полуколичественного ОТ-ПЦР (рис. 1А, Б) и ОТ-ПЦР в реальном времени (рис. 1В) показано, что уровень мРНК ГР в гиппокампе ручных крыс почти в 2 раза выше, чем у агрессивных. Таким образом, можно заключить, что появившиеся в результате отбора по поведению различия в количестве цитозольных ГР в гиппокампе у агрессивных и ручных крыс (Оськина, Плюснина, 2000) в значительной степени связаны с уровнем мРНК ГР в этой же структуре мозга.
При исследовании влияния отбора по поведению на активность ГГНС важно выяснить, изменяется ли количество ГР в периферических органах, основных мишенях глюкокортикоидов. Метод ОТ-ПЦР в реальном времени не выявил достоверных отличий в содержании мРНК ГР ни в печени, ни в селезёнке между ручными и агрессивными крысами (REST 2009). Таким образом, можно предполагать, что наблюдаемые в результате отбора по поведению изменения коснулись, главным образом, именно ГР в гиппокампе, участвующих в отрицательной обратной связи ГГНС.
Влияние пренатальной метилобогащённой диеты на активность ГГНС у крыс, селекционируемых по поведению
Из литературы известно, что метилирование CpG-динуклеотидов регуляторной зоны гена ГР может играть немалую роль в регуляции количества ГР в гиппокампе, а, следовательно, и активности ГГНС через усиление или ослабление глюкокортикоидной отрицательной обратной связи (Weaver et al., 2004). Показано также, что введение метионина в мозг крыс взрослым животным изменяет профиль метилирования ДНК и уровень экспрессии ряда генов, в том числе и гена ГР (Weaver et al., 2004a, 2004b, 2005, Colvis et al., 2005). Единственным доступным методом повышения уровня метионина при пренатальном развитии является введение компонентов цепи метаболизма S-аденозилметионина в организм матери. Известно, что материнская метилсодержащая диета приводит к изменениям в формировании ЦНС и в профиле метилирования некоторых генов (см. Van den Veyver, 2002; Cooney et al., 2002). Для исследования связи метилирования ДНК с различиями между ручными и агрессивными крысами и его влияния на пренатальное «программирование» активности ГГНС был поставлен эксперимент по пренатальному содержанию крыс на метилобогащённой диете. Исследовались четыре группы животных: 1) потомки ручных крыс, содержавшихся во время беременности на метилобогащённой диете (ручные опытные); 2) потомки агрессивных крыс, содержавшихся во время беременности на метилобогащённой диете (агрессивные опытные); 3) потомки ручных крыс, получавшие во время беременности стандартное питание (ручные контрольные); 4) потомки агрессивных крыс, получавшие во время беременности стандартное питание (агрессивные контрольные).
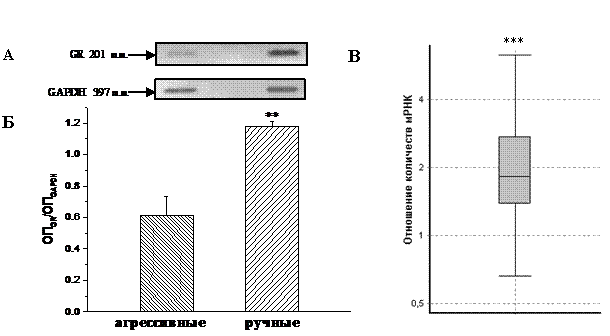
Рис. 1. Количество мРНК ГР в гиппокампе агрессивных и ручных серых крыс.
А – электрофореграмма амплификацированного с помощью полу-количественной ОТ-ПЦР участка мРНК ГР (201 п. н.) и GAPDH (397 п. н.; внутреннего контроля); Б – относительная оптическая плотность амплифи-цированного в полуколичественной ОТ-ПЦР участка мРНК ГР (n = 8 животных в каждой группе); В – отношение количеств мРНК ГР, определённое с помощью ОТ-ПЦР в реальном времени (n = 4 животных в каждой группе) – верхняя и нижняя границы прямоугольника охватывают 50% наблюдений, медиана соответствует значению отношения количеств мРНК ручных крыс по сравнению с агрессивными (REST 2009); **P<0,01, ***P<0,001 у ручных крыс больше по сравнению с агрессивными.
Влияние пренатальной метилобогащённой диеты на базальный и стрессорный уровень кортикостерона в крови у взрослых крыс
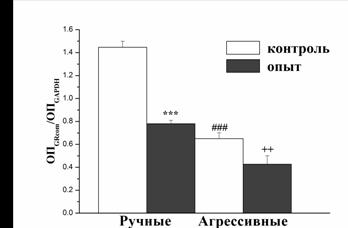
У четырёх групп крыс была изучена временная динамика ответа ГГНС на 30-минутный рестрикционный стресс (рис. 3). Были повторены результаты, полученные при сравнении контрольных ручных и агрессивных крыс (Дыгало и др., 1985; Оськина, Плюснина, 2000). Уровень кортикостерона в крови у ручных животных ниже, чем у агрессивных, а также различна динамика ответа ГГНС на рестрикционный стресс (F1,20 = 20,2, P<0,001). Двухфакторный дисперсионный анализ также выявил отличия в динамике ответа и между опытными и контрольными ручными крысами (F(1,14) = 18,4, P<0,001), тогда как у агрессивных животных различий в опыте и контроле не выявлено. Уровень кортикостерона через час и 2 часа после окончания стресса у ручных опытных крыс достоверно выше, чем у их контроля (P<0,001), тогда как у агрессивных животных достоверные различия в этих временных точках не выявлены. Базальный же уровень кортикостерона не отличался между опытом и контролем ни у ручных, ни у агрессивных. Таким образом, по-видимому, можно заключить, что метилобогащённая диета ослабляет глюкокортикоидную отрицательную обратную связь.
Экспрессии гена рецептора глюкокортикоидов в гиппокампе серых крыс, матери которых содержались во время беременности на метилобогащённой диете
Метод полуколичественной ОТ-ПЦР показал разницу в уровне мРНК ГР в гиппокампе между ручной контрольной и ручной опытной группами (рис. 4). Достоверных различий между агрессивной опытной и агрессивной контрольной группами выявлено не было (рис. 4). Это хорошо согласуется с данными о временной динамике уровня кортикостерона в ответ на стресс (рис. 3) и является дополнительным подтверждением предположения о влиянии материнской метилобогащенной диеты на отрицательную обратную связь у ручных крыс.
Поскольку регуляция количества ГР и, соответственно, активности ГГНС, по-видимому, во многом происходит за счёт изменения уровня мРНК ГР с разными альтернативными экзонами (McCormick, 2000), важно было исследовать их содержание в гиппокампе у ручных и агрессивных крыс. Кроме того, известно, что их регуляция может происходить с помощью метилирования ДНК (Weaver et al., 2005; Lillycrop et al., 2008), поэтому необходимо также было исследовать количества сплайс-вариантов мРНК ГР у потомков крыс, содержащихся на метилобогащённой диете во время беременности. Специфичным для гиппокампа является вариант мРНК ГР с экзоном 17, тогда как мРНК с экзоном 110 преобладает во всех тканях.
Метод ОТ-ПЦР в реальном времени не выявил различий в уровне мРНК ГР с экзоном 17 в гиппокампе ни у одной из четырёх групп крыс (контрольные ручные – 0,54±0,03, контрольные агрессивные – 0,44±0,20, опытные ручных – 0,63±0,25, опытные агрессивные – 0,44±0,19; P>0,05; n = 3 животных в каждой группе). Исследования количества мРНК ГР с экзоном 110 в гиппокампе показали достоверные различия между ручной контрольной группой крыс и ручной и агрессивной опытными группами животных (рис. 5). Достоверного

Рис. 3. Уровень кортикостерона в крови после 30-мин. рестрикционного стресса (n = 12 животных в каждой группе) у потомков агрессивных (А) и ручных (Б) крыс, содержавшихся во время беременности на метилобогащённой диете или стандартном питании. **P<0,01 агрессивные опытные по сравнению со своим контролем; ***P<0,001 ручные опытные по сравнению со своим контролем; +P<0,05, +++P<0,001 ручные опытные по сравнению с агрессивными контрольными.
![]()
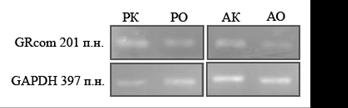
Рис. 4. Влияние материнской метилобогащённой диеты на количество мРНК ГР в гиппокампе серых крыс (n = 8 животных в каждой группе);
|
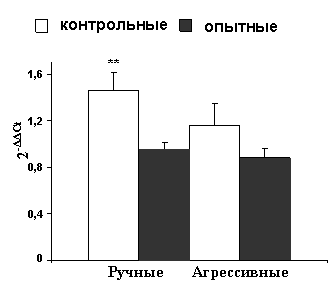
Рис. 5. Влияние материнской метилобогащённой диеты на количество мРНК ГР, содер-жащей экзон 110, в гиппокампе серых крыс (n = 3 животных в каждой группе). Количественный анализ амплифицированного участка экзона 110 транскрипта гена ГР; **P<0,01 ручные конт-рольные по сравнению с ручными и агрессивными опытными.
различия между агрессивной контрольной группой и другими группами обнаружено не было (1,16±0,19; P>0,05).
Таким образом, экзон 110 достоверно снижается у ручных крыс под влиянием метилобогащённой диеты, тогда как его роль в изменении количества мРНК ГР при отборе по поведению не установлена.
Исследование метилирования промотора экзона 17 гена рецептора глюкокортикоидов в гиппокампе
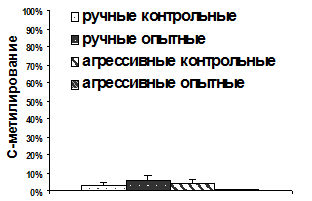
Чтобы установить, влияет ли материнская метилобогащённая диета на профиль метилирования промотора экзона 17 гена ГР в гиппокампе крыс, контрастных по поведению, а также выяснить, вызвал ли отбор по поведению изменение этого профиля, был использован метод бисульфитного секвенирова-ния ДНК. Промотор экзона 17 гена ГР во всех группах оказался гипо-метилирован и уровень метилирования ДНК промотора достоверно не различался между группами (рис. 6А). Таким образом, уровень метилирования промотора экзона 17 гена ГР в гиппокампе не изменялся под влиянием материнской метилобогащённой диеты как у ручных, так и у агрессивных крыс.
16 и 17-ый CpG-динуклеотиды исследованного участка являются 5'- и 3'-CpG в консенсусной последовательности фактора транскрипции NGFI-A, соответственно. По мнению Вивера с соавт., деметилирование именно 5'-CpG этого участка при неонатальных воздействиях повышает общее количество мРНК ГР и уровень мРНК ГР с экзоном 17, а также снижает активность ГГНС. При этом 3'-CpG остаётся гиперметилирован независимо от раннего опыта (Weaver et al., 2004). Анализ показал низкий уровень метилирования как 16-го, так и 17-го CpG в гиппокампе и у ручных, и у агрессивных крыс (рис. 6Б). Материнская метилобогащённая диета также достоверно не изменила уровень метилирования этого участка промотора экзона 17 (рис. 6Б).
Таким образом, профиль метилирования экзона 17 гена ГР в гиппокампе не изменялся ни под влиянием материнской метилобогащённой диеты, ни при отборе по поведению.
|
|

Полученные результаты гипометилирования промотора экзона 17 гена ГР у ручных и агрессивных крыс подняли вопрос о том, не является ли метилирование этого участка ДНК характерным только для определённой линии крыс. Вместе с тем, считается, что в популяциях диких серых крыс также существуют различия в поведении и связанных с ним физиологических реакциях. Различный уровень стресс-ответа считается адаптивным для животных, живущих в местах, отличающихся по комфорту и обогащённости ресурсами. Предполагается, что этот уровень связан с материнской средой и устанавливается с помощью тех же механизмов, что и обнаруженные при хэндлинге и различном уровне материнской заботы (Meaney et al., 1996). Исследования, проведённые на диких серых крысах, отловленных в окрестностях Академгородка и в Новосибирской области дали результаты, аналогичные полученным на крысах, селекционируемых по поведению. Промотор экзона 17 гена ГР в гиппокампе диких крыс также оказался гипометилирован (0,54±0,33%; n=5), при этом 5' и 3' CpG-динуклеотиды в консенсусной последовательности фактора транскрипции NGFI-A были вообще неметилированы.
Изучение последствий материнской метилобогащённой диеты на активность ГГНС у второго поколения потомков серых крыс, селекционируемых по поведению
Из литературы известно, что резкое изменение гормональных систем регуляции путём введения некоторых веществ в организм крысы во время беременности могут сказываться на потомстве в ряду поколений (Anway et al., 2006; Skinner et al., 2008). Поэтому небезынтересным казалось исследовать и второе поколение (F2) потомков крыс, получавших во время беременности метилобогащённую диету. Матери (F1) этих животных содержались на стандартном питании.
Исследование уровня кортикостерона в крови показало, что эффекты материнской метилобогащённой диеты сохраняется и во втором поколении потомков. Двухфакторный дисперсионный анализ выявил отличия в динамике ответа ГГНС на рестрикционный стресс между опытными и контрольными ручными крысами из поколения F2 (рис. 7; F1,11=7,64, P<0,05), тогда как у агрессивных животных различий в опыте и контроле не выявлено. Уровень кортикостерона через час после окончания стресса у ручных опытных крыс достоверно выше, чем у их контроля (P<0,01), однако уже через 2 часа, в отличие от животных из поколения F1, возвращается к базальным показателям. У агрессивных крыс достоверные различия в этих временных точках не выявлены. Базальный же уровень кортикостерона не отличается между опытом и контролем ни у ручных, ни у агрессивных крыс.
Оценка уровня мРНК ГР в гиппокампе также выявила влияние материнской диеты на второе поколение потомков. Метод ОТ-ПЦР в реальном времени показал, что в отличие от крыс из материнского поколения (P) количество мРНК ГР в гиппокампе у ручных крыс из поколения F2 ниже, чем у агрессивных животных из того же поколения (рис. 8А). Близкие результаты были получены в исследовании количества мРНК ГР с экзоном 110. Анализ полученных данных

Рис. 7. Уровень кортикостерона в крови после 30-минутного рестрикционного стресса (n=10 животных в каждой группе) у потомков из F2 агрессивных (А) и ручных (Б) крыс, содержавшихся во время беременности на метилобогащённой диете или стандартном питании. **P<0,01 агрессивные опытные по сравнению со своим контролем; ***P<0,001 ручные опытные по сравнению со своим контролем; ++P<0,01 ручные опытные по сравнению с агрессивными контроль-ными.
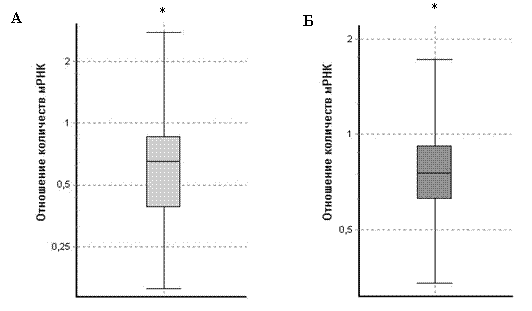 |
Рис. 8. Количество мРНК ГР в гиппокампе агрессивных и ручных серых крыс из поколения F2 (n=4 животных в каждой группе), определённое с помощью ОТ-ПЦР в реальном времени; описание графика представлено в подписи к рис. 1В.
А – отношение общих количеств мРНК ГР в гиппокампе, Б – отношение количеств мРНК ГР с экзоном 110 в гиппокампе; *P<0,05 у ручных крыс меньше по сравнению с агрессивными.
также выявил, что уровень мРНК ГР с экзоном 110 в гиппокампе ручных крыс из поколения F2 ниже, чем у агрессивных из того же поколения (рис. 8Б).
Таким образом, исходя из полученных данных, можно заключить, что метилобогащённая диета влияет на активность ГГНС у потомков как первого, так и второго поколения. При этом изменение экспрессии гена ГР, по-видимому, происходит за счёт уровня транскрипции под контролем промотора экзона 110. Возможно, это связано с изменением уровня его метилирования. В то же время, наблюдаемая различная чувствительность к повышению концентрации пред-шественников S-аденозилметионина у ручных и агрессивных крыс, может быть связана с метилированием неисследованных нами участков генома.
С другой стороны, отличия в экспрессии гена ГР у ручных и агрессивных крыс, по-видимому, не обусловлены изменением количества мРНК с экзоном 110. Вероятно, отбор по поведению вообще не вызывает специфических изменений в уровне транскрипции под контролем альтернативных промоторов гена ГР, а приводит к общему снижению количества мРНК ГР независимо от альтернативных экзонов 1, что и даёт достоверные отличия. Такой вариант регуляции гена рецептора был обнаружен при хроническом стрессе у крыс (Freeman et al., 2004). Было установлено значительное снижение количества всех исследованных вариантов мРНК ГР, однако механизм этого снижения не установлен. В то же время, можно предположить, что пренатальные манипуляции, как и пренатальная метилобогащённая диета затрагивают экспрессию гена ГР именно под контролем промотора экзона 110.
ВЫВОДЫ
1. Сравнительный анализ уровня мРНК рецептора глюкокортикоидов (ГР) выявил более высокий уровень экспрессии гена ГР в гиппокампе «ручных» серых крыс – селекционируемых на элиминацию агрессивно-оборонительной реакции по отношению к человеку, чем у «агрессивных» животных – селекционируемых на усиление этой реакции. В то же время различий в количестве мРНК ГР в печени и селезёнке между ручными и агрессивными животными не обнаружено, что свидетельствует о том, что мишенями отбора служат ГР в гиппокампе, являющиеся существенным звеном глюкокортикоидной отрицательной обратной связи.
2. Не выявлено достоверных различий экспрессии гена ГР в гиппокампе под контролем промоторов альтернативных экзонов 17 и 110 между серыми крысами, селекционируемыми на ручное и агрессивное поведение.
3. Содержание крыс во время беременности на метилобогащённой диете приводит к более позднему возвращению базальных значений стресс-индуцированного уровня кортикостерона в крови у первого и второго поколения потомков ручных матерей, тогда как у потомков агрессивных крыс эффекта диеты на активность гипотоламо-гипофизарно-надпочечниковой системы (ГГНС) не выявлено.
4. Изменение активности ГГНС под влиянием материнской метилобогащённой диеты в значительной степени связано с количеством мРНК ГР, содержащей экзон 110, в гиппокампе. У первого поколения потомков ручных крыс материнская диета снижает уровень мРНК ГР с экзоном 110 и общее количество мРНК ГР почти до уровня агрессивных животных, у которых эти показатели не изменяются под влиянием диеты. Во втором поколении общее количество мРНК ГР и уровень мРНК ГР с экзоном 110 в гиппокампе у ручных крыс ниже, чем у агрессивных. Полученные факты могут свидетельствовать, что пренатальная метилобогащённая диета ослабляет у ручных животных глюкокортикоидную отрицательную обратную связь.
5. Анализ профиля метилирования промотора экзона 17 гена ГР в гиппокампе не выявил достоверных различий между ручными и агрессивными крысами как в условиях стандартной, так и материнской метилобогащённой диеты и показал гипометилирование этого участка ДНК.
6. Совокупность полученных в работе данных даёт основание считать, что отбор по поведению в отличие от материнской метилобогащённой диеты изменяет количество всех вариантов мРНК ГР; материнская же метилобогащённая диета, эффект которой наблюдается только у ручных потомков, вероятно, по-разному меняет профиль метилирования ДНК у животных контрастного поведения.
СПИСОК ПУБЛИКАЦИЙ
1. , , , Гулевич гипоталамо-гипофизарно-надпочечниковой и иммунной систем при отборе животных на доместикационное поведение // Информ. вестник ВОГиС. — 2008. — Т. 12. — № 1/2. — С. 39—49. (перечень ВАК)
2. Herbeck Yu. E., Gulevich R. G., Amelkina O. A., Plyusnina I. Z., Oskina I. N. Conserved methylation of the glucocorticoid receptor gene exon 17 promoter in rats subjected to a maternal methyl-supplemented diet // Int. J. Dev. NeurosciV. 28. - N 1. - P. 9-12. (перечень ВАК)
3. , , Плюснина материнской метилобогащённой диеты на экспрессию гена рецептора глюкокортикоидов в гиппокампе у крыс селектируемых по поведению // Цитол. генет. — 2010. — Т. 44. — № 2. — C. 45—52. (перечень ВАК)
4. , , , Гулевич гена глюкокортикоидных рецепторов у серых крыс при отборе по поведению // XX съезд Физиологического общества им. . Тезисы докладов — М., 2007. — С. 364.
5. , , Оськина рецепторы мозга и изменение их экспрессии при экспериментальной доместикации серых крыс // Тез. междунар. конф. «Развитие эволюционной идеи в биологии, социологии и медицине». — Новосибирск, 2007. — С. 25.
6. , , Взаимодействие генотипа и материнских факторов в детерминации поведения и гормонального ответа на стресс у серых крыс // Тез. докл. VI Сибирского физиологического съезда. — Т. 2. — Барнаул, 2008. — С. 7.
7. , , Плюснина отбора по поведению и метилобогащенной диеты на экспрессию гена рецептора глюкокортикоидов в гиппокампе крыс // Сб. научн. труд. «Фактории експериментальноi еволюцii органiзмiв». — Т. 4. — К., 2008. — С. 246—250.
8. , , Плюснина материнской диеты на экспрессию сплайс-вариантов гена рецептора глюкокортикоидов у потомков крыс, селекционируемых на доместикацию // Тез. докл. V cъезда Вавиловского общества генетиков и селекционеров. Москва, 21—28 июня 2009 г. — Ч. 2. — C. 111.
9. Herbeck Y. E., Plyusnina I. Z., Oskina I. N. Is the methylation of the glucocorticoid receptor exon 17 promoter universal in rats? // The 7th International Congress of Neuroendocrinology, Rouen, France, 11-15 July 2010 - P. 122.
10. , Оськина отбора по поведению и пренатальной метилобогащённой диеты на экспрессию гена рецептора глюкокортикоидов в гиппокампе серых крыс // XXI съезда Физиологического общества им. . Тез. докл. — М. — Калуга, 2010. — С. 141.
Подписано к печати 05.10.2010 г.
Формат бумаги 60 х 90 1/16. Печ. 1 л. Уч. Изд. 0,7
Тираж 110 экз. Заказ 85.
 Ротапринт Института цитологии и генетики СО РАН
Ротапринт Института цитологии и генетики СО РАН
Новосибирск, пр. ак. Лаврентьева, 10.
