


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
Влияние экологических факторов
на рост растений лесообразующих видов
в природных комплексах северной Карелии
03.00.16 – экология
03.00.05 – ботаника
Автореферат диссертации на соискание ученой степени кандидата
биологических наук
Петрозаводск – 2007
Работа выполнена в ГОУ ВПО «Петрозаводский государственный университет»
Научный руководитель доктор биологических наук, профессор
Официальные оппоненты: доктор биологических наук, старший научный сотрудник
Институт биологии Карельского научного центра РАН
доктор биологических наук, доцент
Институт леса Карельского
научного центра РАН
Ведущая организация Карельский государственный
педагогический университет
Защита состоится 17 октября 2007 г. в 14.00 часов на заседании диссертационного совета Д 212.190.01 при Петрозаводском государствен-
ном университете Республика Карелия, г. Петрозаводск, пр. Ленина, 33, эколого-биологический факультет, ауд. 326 теоретического корпуса.
С диссертацией можно ознакомиться в библиотеке Петрозаводского государственного университета.
Автореферат разослан «17» сентября 2007 года
![]() Ученый секретарь
Ученый секретарь
![]()
диссертационного совета Крупень И. М.
ВВЕДЕНИЕ
Актуальность темы. Picea abies (L.) Karst является одним из главных лесообразующих видов в северной Карелии, занимая 24 % лесопокрытой площади республики (Государственный доклад…, 2006). Еловые леса играют важную средообразующую роль и являются источником ценной древесины. Между тем площади этих лесов неуклонно сокращаются в результате интенсивных рубок главного пользования. В связи с этим решение проблемы повышения продуктивности, устойчивости и рационального использования еловых лесов становится все более актуальным. Между тем разработка эффективных мероприятий по повышению продуктивности лесов возможна только на основе глубоких знаний о ходе формирования прироста древесины стволов и фитомассы других вегетативных органов (Сукачёв, 1950; Молчанов, 1961; Елагин, 1980; Острошенко, 1991).
Берёзовые, осиновые и сероольшаниковые леса являются производными. С экономической точки зрения они не представляют большого интереса и потому должны сменяться на хозяйственно более ценные коренные ельники и сосняки. Успешность решения этой проблемы
во многом зависит от знания процессов формирования органического вещества в древостоях.
Важнейшими процессами, характеризующими жизненность растений лесообразующих видов, являются рост и развитие (Булыгин, 1979; Встовская, 1983; Трулевич, 1991; Шкутко, 1991). Именно зависимость ростовых процессов растения от постоянно изменяющихся экологических факторов и приводит его к определённому состоянию адаптации. В связи с этим изучение эколого-биологических особенностей роста растений лесообразующих видов имеет чрезвычайно важное как теоретическое, так и практическое значение. Раскрытие таких закономерностей является одной из центральных и вместе с тем слабо изученных проблем современной биологии.
Цель и задачи исследования. Целью данного исследования является изучение особенностей сезонной ритмики роста растений основных лесообразующих видов в природных комплексах на севере Карелии для раскрытия особенностей биопродукционных процессов в естественных древостоях.
В задачи исследования входило:
1. Выявить зависимость роста растений Picea abies, Betula pubescens Ehrn., Populus tremula L. и Alnus incana (L.) Moench. от экологических факторов.
2. Установить особенности роста растений Picea abies и Betula pubescens в разных типах леса.
3. Определить продолжительность и интенсивность роста растений Picea abies и Betula pubescens на разных этапах онтогенеза.
4. Установить закономерности сезонного роста растений Picea abies в зависимости от фитоценотического положения деревьев.
5. Выявить особенности анатомического строения древесины ствола Picea abies.
Научная новизна. Впервые на территории Карелии проведены
исследования сезонного роста побегов, листового аппарата и стволов Picea abies, Betula pubescens, Populus tremula и Alnus incana под влиянием основных экологических факторов. При этом установлены их оптимальные и экстремальные для ростовых процессов значения. Приводится математическое описание обнаруженных связей. Определена интенсивность деятельности меристематических тканей у деревьев в разных типах леса. Выявлена не известная ранее зависимость сезонного роста от возраста дерева и его фитоценотического положения. В результате проведённых исследований установлены важные закономерности в сезонном росте деревьев, позволяющие наметить некоторые пути повышения продуктивности еловых древостоев.
Теоретическая и практическая значимость работы. В результате исследований установлены характер и степень влияния экологических факторов на динамику роста растений основных лесообразующих видов таежной зоны. Выявлены особенности этого процесса у деревьев в различных типах леса и на разных этапах онтогенеза. Кроме того, установлена зависимость ростовых процессов дерева от его фитоценотического положения. Полученные результаты могут служить основой для разработки системы мероприятий по рациональному использованию таежных ельников, повышению их продуктивности, устойчивости и средообразующей роли. Вместе с тем они могут быть использованы также для организации мониторинга и производства лесокультурных работ.
Результаты исследований включены в программы дисциплин «Экология», «Ботаника» и «Лесоведение» (Петрозаводский государственный университет), используются при чтении соответствующих лекционных курсов, проведении лабораторных работ по экологии и полевых практик по ботанике на эколого-биологическом и лесоинженерном факультетах Петрозаводского государственного университета.
Организация исследований и личный вклад автора. Исследования проводились на 10 постоянных пробных площадях, заложенных на территории Муезерского района Республики Карелия. Диссертационная работа выполнялась в 2002–2007 гг. Вначале исследования проводились в рамках дипломной работы. Начиная с 2005 г. исследования выполнялись в качестве раздела плановой научно-исследовательской темы кафедры ботаники и физиологии растений ПетрГУ «Рост и развитие аборигенных и интродуцированных древесных растений в Карелии». Климатические факторы регистрировались на Ребольской мете-
орологической станции (Карельская гидрометеорологическая обсерватория). Автором лично проведены все полевые исследования. Диссертантом выполнены камеральная и статистическая обработка экспериментальных материалов, а также анализ полученных результатов.
Достоверность и обоснованность результатов исследования.
В процессе исследований использовались современные научные методы, применяемые в экологии и ботанике. Обоснованность и достоверность результатов исследований обеспечивались применением методик планирования исследований, необходимым объемом выборок по каждому признаку с учетом их изменчивости, соответствующих рекомендаций и наставлений. Оценка результатов осуществлялась на 5%-м уровне значимости. Полученные данные обработаны с помощью методов вариационной статистики. Для установления зависимостей между изучаемыми признаками и параметрами среды применены корреляционный, дисперсионный и регрессионный анализы. Материалы обработаны на персональном компьютере с помощью соответствующих пакетов программ.
Апробация работы. Основные положения и результаты исследований представлялись на международной конференции «Структурно-функциональные особенности биосистем севера (особи, популяции, сообщества)» (Петрозаводск, 2005); межрегиональной общественно-научной конференции с международным участием «Запад России и ближнее Зарубежье: устойчивость социально-культурных и эколого-хозяйственных систем (Псков, 2005); X Пущинской школе - конференции «Школа-конференция молодых учёных» (Пущино, 2006); IV Всероссийской научно-технической конференции (Вологда, 2006); международной конференции «Современные экологические проблемы севера» (Апатиты, 2006); международной конференции «Устойчивость экосистем и проблема сохранения биоразнообразия на севере» (Кировск, 2006); VII международной молодёжной научной конференции «Севергеоэкотех–2006» (Ухта, 2006); IV Российской конференции «Флора и растительность Сибири и Дальнего Востока» (Красноярск, 2006).
Публикации. По материалам исследования опубликовано 11 научных работ, в т. ч. 1 статья в рецензируемом издании, включённом
в требуемый список ВАК.
Структура и объём диссертации. Диссертация изложена на 203 страницах и состоит из введения, общей характеристики работы, шести глав, заключения и 13 приложений. Список литературных источников включает 161 наименований, из которых 25 на иностранных языках. Текст иллюстрирован 27 таблицами и 53 рисунками.
Благодарности. Автор выражает искреннюю благодарность научному руководителю доктору биологических наук, профессору И. Т. Кищенко, сотрудникам кафедры ботаники и физиологии растений ПетрГУ, а также сотруднику Карельского научного центра РАН Е. Н. Спектор. Особую признательность за помощь в работе на всех этапах ее выполнения выражаю моим родителям.
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ
ПО ТЕМЕ ИССЛЕДОВАНИЙ
Изучению сезонного роста растений, в том числе и древесных
видов, уделяется большое внимание и в России, и за рубежом. Объектами исследований служат аборигенные виды, как хвойные, так
и лиственные.
В главе приводится обзор специальной литературы по сезонному росту растений лесообразующих видов преимущественно таёжной зоны (Мелехова, 1954; Харитонович, 1961, 1962; Молчанов, 1961; Смирнов, 1964; Елагин, 1976, 1980; Елагина, 1969; Мамаев, 1970; Фролова, 1979; Верзунов, 1982; Абатуров, 1985; Фёдоров, 1987; Острошенко, 1991; Забуга, 1992; Лёвкина, 1996; Тимофеев, 1996; Micola, 1950; Ladefoged, 1952; Kozlowski, 1963; Leicola, 1969; Odin, 1972; Worral, 1973; Serre, 1976; Yunttila, Heide, 1981; Кищенко, 2000).
Анализ литературных источников показал, что интенсивность биопродукционных процессов древостоев определяется в конечном итоге продолжительностью и скоростью роста побегов, листового аппарата и стволов деревьев, на которые оказывает существенное влияние ряд экологических факторов.
При этом многие стороны формирования побегов, хвои, листьев и стволов растений лесообразующих видов в течение вегетации изучены слабо и нуждаются в дальнейшем уточнении. В частности, отсутствуют сведения об экстремальных и оптимальных значениях факторов среды для роста побегов, ассимиляционного аппарата и стволов древесных растений. Нет достаточной ясности и в вопросе о зависимости этого процесса от фитоценотического положения дерева и его возраста. Исследования сезонного роста растений Picea abies в Карелии ранее носили эпизодический характер. Сведения по формированию побегов, листьев и стволов Betula pubescens, Populus tremula и Alnus incana в специальной литературе не обнаружены. Все это, а также почти полное отсутствие подобных фактических данных для Карелии объясняет выбор направления исследований и темы настоящей работы.
ГЛАВА 2. КРАТКИЙ ОЧЕРК ПРИРОДНЫХ УСЛОВИЙ КАРЕЛИИ, ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
2.1. Физико-географические условия района исследований
В диссертации даётся характеристика рельефа и климата Карелии. Приводятся среднемноголетние данные по радиационному режиму
и продолжительности периода с температурой воздуха выше +5, +10
и +15 ºС по срокам наступления первых осенних и последних весен-
них заморозков, влажности воздуха, облачности и количеству осадков. Перечислены типы почв, на которых формируются еловые, берёзовые, осиновые и ольховые древостои.
Лесная растительность Карелии представлена еловыми, сосновыми и березовыми лесами. Еловые леса являются коренными и занимают 24% лесопокрытой площади. Берёзовые леса вторичного происхождения занимают 9%, осинники – 0.5%, ольшаники – 0.2% покрытой лесом площади (Государственный доклад…, 2006).
Суровые климатические условия и бедность песчаных почв обусловливают низкую производительность еловых лесов. Средний класс бонитета их равен IV.0, текущий прирост – 1.3 м3/га (Государственный доклад…, 2006).
2.2. Объекты исследований и их характеристика
Исследования проводили в 2002–2007 гг. в северной Карелии (подзона северной тайги). Объектами исследований служили еловые, берёзовые, осиновые и сероольшаниковые древостои.
Рост растений Picea abies изучали в чистых по составу (в отдельных случаях с небольшой примесью Betula pubescens и Pinus sylvestris) приспевающих древостоях четырёх типов леса: ельник черничный, ельник брусничный, ельник болотно-травяной, ельник сфагновый. Кроме того, заложены две пробные площади в молодом и спелом древостоях ельника черничного.
Рост растений Betula pubescens изучали в чистых по составу (с небольшой примесью Populus tremula) 67-летних и 78-летних древостоях двух типов леса (березняк чернично-разнотравный, березняк злаково-брусничный). Кроме того, в березняке чернично-разнотравном пробные площади заложены в 30-летнем и 67-летнем древостоях.
Рост растений Populus tremula изучали в чистом по составу древостое (с примесью Betula pubescens) в условиях осинника злаково-черничного.
Рост растений Alnus incana изучали в чистых по составу древостоях (с примесью Betula pubescens) в условиях сероольшаника злаково-разнотравного.
Почва, где произрастают изученные древостои, относится к типу иллювиально-железисто-гумусового подзола средней мощности. Почва под ельником сфагновым характеризуется мощным слоем торфа, большой оглеенностью и более глубоким залеганием минерального грунта, подстилающего торфяник. В диссертации приводится геобо-
таническая характеристика фитоценозов.
При закладке пробных площадей и геоботаническом описании фитоценозов использованы соответствующие методические рекомендации (Полевая геоботаника, 1972; Программа…, 1974). Всего заложено 10 пробных площадей.
Формирование стволов, побегов второго порядка и фотосинтезирующего аппарата исследовали у деревьев II–V классов роста (по Крафту). Измерения проводили с мая по август через каждые 3–5 суток. Изменение длины стеблевой части побегов регистрировали линейкой (Молчанов, Смирнов, 1967).
Рост хвои (по длине) и листьев (по площади) изучали по методу, предложенному В. В. Смирновым (1961). Анализировали листовой аппарат в средней части этих же побегов преимущественно с юго-западной стороны.
Формирование радиального прироста у стволов изучали методом высечек (Молчанов, Смирнов, 1967). Образцы древесины вместе с фло-
эмой вырезали с западной стороны стволов на высоте 1.3 м по спирали снизу вверх и слева направо, каждый раз отступая от места предыдущей высечки на 0.5 см. Препараты древесины для просмотра готовили с помощью микротома (Яценко-Хмелевский, 1954). На этих же препаратах измеряли толщину клеточных оболочек трахеид в радиальном направлении в трёх местах.
Климатические факторы (продолжительность солнечного сияния; атмосферные осадки; среднесуточная температура и относительная влажность воздуха) регистрировались на Ребольской метеорологической станции (Карельская гидрометеорологическая обсерватория), расположенной в 3 км к северо-востоку от объектов исследований.
По результатам наблюдений за ростом, а также за динамикой климатических факторов сформировали банк данных, обработанный с помощью корреляционного, регрессионного и дисперсионного анализов.
ГЛАВА 3. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ РОСТА PICEA ABIES (L.) KARST.
3. 1. Влияние климатических факторов на рост растений
Изучение растений Picea abies позволило обнаружить, что рост их побегов, хвои и стволов в разных условиях местопроизрастания происходит в определённой последовательности. Первыми (конец мая) трогаются в рост побеги второго порядка, спустя 6–16 суток появляется молодая хвоя и начинается деление клеток камбия ствола. Кульминация прироста побегов наблюдается в начале июля, стволов – в середине июля, хвои – в конце июля. Рост побегов заканчивается во второй половине июля, ствола – в августе, хвои – в конце августа. Наибольшая продолжительность роста характерна для хвои – 60–87 суток, у стволов она составляет 57–59 суток, а у побегов – 45–65 суток.
Исследованиями установлено, что рост растений Picea abies в основном связан с динамикой температуры воздуха. Деятельность камбия ствола начинается при среднесуточной температуре воздуха +7.2–+9.4 ºС, хвои – при +7.4–+8.6 °С, стволов – при +11.9–+20.6 °С. На сроки начала роста побегов, хвои и стволов дерева существенное влияние оказывает и температурный режим периодов, предшествующих этим фенофазам. Температура воздуха оказывает влияние на интенсивность роста побегов, хвои и стволов в течение всего периода их формирования. Однако наиболее заметно ее воздействие на процессы роста в период времени, предшествующий кульминации прироста, что согласуется с мнением многих исследователей (Смирнов, 1961; Lanner, 1964; Мамаев, 1970; Молчанов, 1970; Кайрюкштис, Юодвалькис, 1972; Junttila, Heide, 1981; Забуга, Забуга, 1992). Кульминация прироста побегов происходит при температуре воздуха выше +17 ºС, хвои – выше +16 ºС, стволов – выше +14 ºС. Резкое снижение темпов роста побегов наблюдается при падении температуры ниже +12 ºС, хвои – ниже +13 ºС, стволов – ниже +7 ºС (рис. 1–3). Прекращение роста побегов происходит при снижении среднесуточной температуры воздуха до +15 ºС, хвои – до +13 ºС, стволов – до +13 ºС.
А
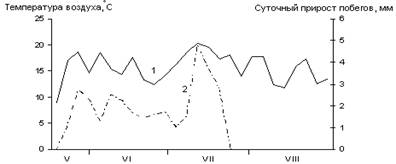
Б
В
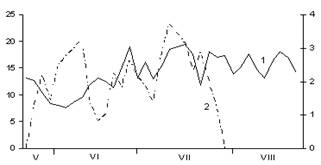
Рис. 1. Сезонная динамика температуры воздуха
и суточного прироста побегов Picea abies.
Годы наблюдений: А – 2002; Б – 2003; В – 2005;
1 – температура воздуха, °С; 2 – суточный прирост побегов, мм
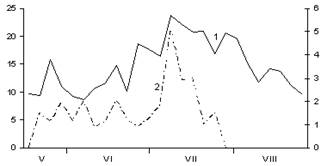
А
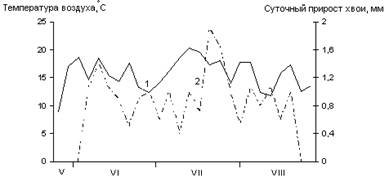
Б
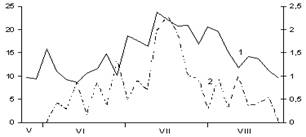
В
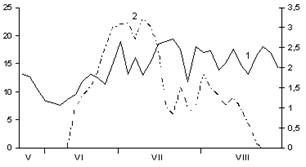
Рис. 2. Сезонная динамика температуры воздуха и суточного прироста хвои Picea abies.
Годы наблюдений: А – 2002; Б – 2003; В – 2005;
1 – температура воздуха, °С; 2 – суточный прирост хвои, мм
А
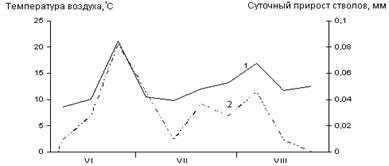
Б
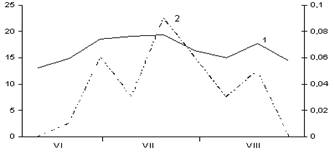
В
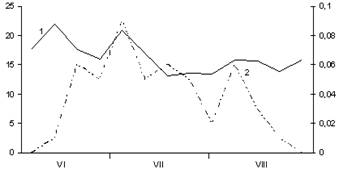
Рис. 3. Сезонная динамика температуры воздуха
и суточного прироста стволов Picea abies.
Годы наблюдений: А – 2003; Б – 2005; В – 2006;
1 – температура воздуха, °С; 2 – суточный радиальный прирост стволов, мм
Количество атмосферных осадков существенно не отражается на сроках начала и окончания роста побегов и хвои. Однако деление клеток камбия ствола в древостоях на избыточно увлажненных почвах
в годы с незначительным количеством летних осадков заканчивается на 4−8 суток позднее, чем в годы, характеризующиеся их обилием. Рост древесины на умеренно увлажненной почве в годы с количеством летних осадков, близких к норме, продолжается на 4−7 суток дольше, чем в годы с их недостатком или избытком.
Среднесуточная влажность воздуха не оказывает заметного влияния на сроки начала и окончания роста побегов, хвои и стволов Picea abies. Однако на интенсивность ростовых процессов этот фактор оказывает некоторое слабое отрицательное воздействие.
Продолжительность солнечного сияния отражается на деятельности меристематических тканей: с её возрастанием она усиливается.
В результате исследований установлено, что зависимость интенсивности роста вегетативных органов дерева (У) от климатических факторов описывается уравнением параболы второго порядка. Для деревьев ельника черничного получены следующие модели роста:
для побегов (мм)
у = 0.046х12 − 0.067х2 − 0.167х3 + 0.045х4 + 0.585х5;
для хвои (мм)
у = 0.059х12 − 0.004х2 − 0.069х3 + 0.039х4 + 0.299х5;
для стволов (мкм)
у = 0.318х12 − 0.283х2 − 0.022х3 + 0.020х4 + 0.006х5,
где Х1 – интервал времени с момента начала роста, сут.; Х2 – темпе-
ратура воздуха, °С; Х3 – относительная влажность воздуха, %; Х4 – количество осадков, мм; Х5 – продолжительность солнечного сияния, час.
3.2. Формирование текущего прироста в различных условиях
местопроизрастания
Тип леса является интегральным показателем, отражающим влияние климата и почвенно-грунтовых условий на интенсивность биопродукционных процессов в лесных фитоценозах. Выяснилось, что различия в условиях местопроизрастания отражаются на особенностях сезонного роста растений Picea abies. В более продуктивных типах леса рост деревьев этого вида начинается на 1–12 суток раньше, а заканчивается на 1–16 суток позже. В целом продолжительность формирования прироста в более благоприятных почвенно-грунтовых условиях возрастает на 7–16 суток. Сравнение сезонного хода роста растений Picea abies в разных типах леса позволило обнаружить существенные различия в величине их годичного прироста, возникающие в результате неодинаковой интенсивности и продолжительности ростовых процессов (табл. 1). Это согласуется с данными и других исследователей (Вомперский, 1968; Ахмеров, 1969; Leicola, 1969; Молчанов, 1970; Орлов и др., 1972).
Таблица 1
Некоторые характеристики прироста побегов *, хвои и стволов Picea abies
Годы | Максимальный | Годичный прирост, мм | Продолжительность | ||||||
побеги | хвоя | стволы | побеги | хвоя | стволы | побеги | хвоя | стволы | |
2002 | 4.8 | 1.9 | − | 70 | 26 | − | 53 | 79 | − |
2003 | 5.4 | 1.5 | 0.04 | 61 | 20 | 3.3 | 56 | 74 | 72 |
2005 | 3.6 | 1.3 | 0.05 | 55 | 17 | 3.0 | 65 | 83 | 77 |
2006 | − | − | 0.03 | − | − | 3.1 | − | − | 78 |
Примечание: * − здесь и далее рассматриваются побеги второго порядка.
На основе данных наблюдений построены графики сезонной динамики роста побегов, хвои и стволов Picea abies в различных типах леса (рис. 4).
Изучение сезонного роста деревьев, занимающих разное фитоценотическое положение, позволило обнаружить, что у деревьев господствующей части древостоя формирование древесины стволов начинается раньше и заканчивается позже. У этих деревьев отмечается также и более интенсивное деление клеток камбия. По этим причинам величина годичного радиального прироста ствола у деревьев господствующей части оказывается в среднем на 95% больше, чем
у деревьев подчинённой части древостоя. Подобная тенденция наблюдалась другими исследователями (Kozlowski, Peterson, 1962; Borowski, 1974; Елагин, 1975).
Продолжительность роста оболочек ранних трахеид составляет 30–40 суток, что на 10–20 суток меньше, чем поздних трахеид. Повышенная интенсивность и большая продолжительность роста оболочек поздних трахеид приводят к увеличению их толщины (5.3–7.9 мкм) по сравнению с ранними трахеидами (1.2–3.5 мкм) в 1.5–3.0 раза.
А
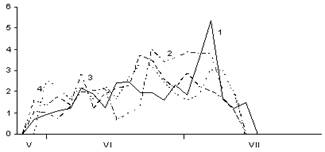
Б
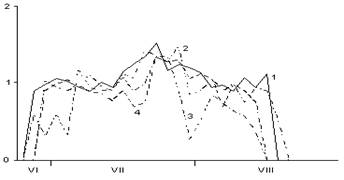
В
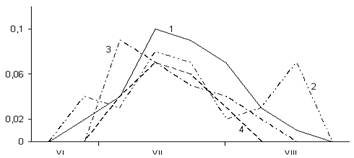
Рис. 4. Сезонная динамика суточного прироста побегов, хвои и стволов Picea abies в разных типах леса (по данным 2005 г.):
А – суточный прирост побегов; Б – хвои; В – стволов. По оси абсцисс – календарные месяцы. По оси ординат – суточный прирост в ельнике черничном (1), е. брусничном (2), е. сфагновом (3) и е. болотно-травяном (4), мм
Формирование оболочек трахеид в лучших почвенно-грунтовых условиях происходит интенсивнее и длится дольше. Поэтому в ельниках зеленомошной группы типов леса оболочки трахеид на 10–70% толще, а доля поздней древесины на 15–20% больше, чем в ельнике сфагновом и ельнике болотно-травяном.
Качество древесины стволов у деревьев подчинённой части древостоя намного ниже, чем у деревьев господствующей части. Это выражается в снижении доли участия поздней древесины в формировании годичного кольца на 12%, а также в уменьшении толщины оболочек поздних трахеид на 10%.
3.3. Особенности роста растений на разных этапах онтогенеза
Исследования показали, что динамика сезонного роста побегов, хвои и стволов Picea abies в процессе онтогенеза несколько меняется. Так, деятельность латеральной меристемы у 30-летних деревьев начинается на 1−6 суток раньше, а заканчивается на 2−10 суток позже, чем у 150-летних. Интенсивность роста стволов с возрастом дерева уменьшается, хвои – увеличивается, а у побегов наибольшей величины она достигает в возрасте приспевания.
Различия в годичном приросте побегов и хвои деревьев разного возраста связаны исключительно с изменением интенсивности их роста, а стволов, кроме того, и с разницей в продолжительности их формирования. Годичный прирост ствола в молодняке в среднем составляет 3.0 мм, в приспевающем – 3.2 мм, в спелом – 2.9 мм, хвои – соответственно 23.0, 23.1 и 24.3 мм, побегов – 47.7, 49.1, 47.5 мм. Различия
в росте деревьев на разных этапах онтогенеза обнаружены и другими авторами (Харитонович, 1961; Лобжанидзе, Канделаки, 1971; Острошенко, 1991; Митрухова, 1993).
Анализ фактического материала позволил обнаружить, что несмотря на различия, связанные с почвенно-грунтовыми условиями, а также
с возрастом и положением дерева в пологе леса, последовательность
в сроках начала, окончания и кульминации деятельности апикальной меристемы не меняется. Установлено, что различия в величине годичного прироста органической массы древостоев на 80–90% обусловлены неодинаковой интенсивностью роста и лишь на 10–20% – различной продолжительностью формирования. Однако основные черты кривых, отображающих прирост побегов, хвои и стволов, также характеризуются значительным сходством. Наряду с этим выяснено, что до 80% годичного прироста надземной части растений формируется в первую половину периода их роста. Это указывает на наличие значительного временнόго резерва для увеличения биологической продуктивности еловых древостоев.
Установленные закономерности сезонного роста еловых древостоев позволяют назначать сроки для наиболее эффективного проведения ряда лесохозяйственных мероприятий: рубок ухода, внесения удобрений, борьбы с болезнями и вредителями леса и др.
ГЛАВА 4. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ РОСТА BETULA PUBESCENS EHRN.
4. 1. Влияние климатических факторов на рост растений
Исследования показали, что рост стволов, побегов второго порядка и листьев Betula pubescens происходит в той же последовательности, что и у Picea abies. Кульминация прироста побегов наблюдается в конце июня – начале июля, листьев – в начале – середине июля, ствола – в середине – конце июля. Рост побегов заканчивается во второй половине июля, листьев – в августе, ствола – в конце августа. Наибольшая продолжительность роста характерна для листьев – 71–89 суток, у стволов она составляет 64–73 суток, а у побегов – 54–63 суток.
Из всех изученных климатических факторов температура воздуха также оказывает наибольшее влияние на ростовые процессы растений Betula pubescens. Формирование побегов начинается при среднесуточной температуре около +13 °С, листьев – +11 °С и ствола – +15 °С. Интенсивность роста побегов зависит от температуры воздуха в докульминационный период, а листьев и стволов – на протяжении всего периода роста. Кульминация прироста побегов происходит при температуре воздуха выше +19 °С, листьев – выше +11 °С, стволов – выше +12 °С. Прекращение роста побегов наблюдается при падении температуры ниже +15 °С, листьев – ниже +13 °С, стволов – ниже +14 °С.
Исследованиями установлено, что продолжительность солнечного сияния оказывает слабое положительное влияние на интенсивность ростовых процессов, а атмосферные осадки и влажность воздуха – слабое отрицательное воздействие.
4.2. Формирование прироста деревьев в различных условиях местопроизрастания
Выяснено, что различия в условиях местопроизрастания отражаются на особенностях сезонного роста растений Betula pubescens. В более продуктивных типах леса рост деревьев этого вида начинается на 1–5 суток раньше, а заканчивается на 1–11 суток позже. Сравнение сезонного хода роста деревьев по типам леса показало, что различия в величине их годичного прироста в основном определяются интенсивностью, а не продолжительностью ростовых процессов (табл. 2). В период кульминации роста в березняке чернично-разнотравном суточный прирост побегов в среднем достигает 10.3 мм, листьев – 4.6 мм2 и ствола – 0.9 мм, а в березняке злаково-брусничном – соответственно 8.8 мм, 4.4 мм2 и 0.7 мм.
Таблица 2
Некоторые характеристики прироста
побегов, листьев и стволов Betula pubescens
Годы | Максимальный | Годичный | Продолжительность | ||||||
побеги | хвоя | стволы | побеги | хвоя | стволы | побеги | хвоя | стволы | |
2005 | 10.3 | 5.7 | 58 | 90.5 | 38.2 | 2.0 | 61 | 87 | 72 |
2006 | 12.4 | 2.8 | 68 | 103.2 | 41.0 | 2.8 | 63 | 71 | 73 |
3.3. Особенности роста растений на разных этапах онтогенеза
Исследования показали, что динамика сезонного роста побегов, листьев и стволов Betula pubescens в процессе онтогенеза также несколько меняется. Образование клеток древесины стволов у 30-летних деревьев начинается на 3−9 суток раньше, а заканчивается на 1−5суток позже, чем у 67-летних. Интенсивность роста побегов листьев и стволов у 67-летних деревьев достигает максимальной величины. Неодинаковая интенсивность в процессах роста обуславливает и различия в годичном приросте побегов, листьев и стволов у деревьев на разных этапах онтогенеза. Величина годичного прироста побегов, листьев и стволов у молодых деревьев (30 лет) больше, чем у сравнительно старых (67 лет), соответственно в среднем на 88, 83 и 75%.
ГЛАВА 5. ВЛИЯНИЕ КЛИМАТИЧЕСКИХ ФАКТОРОВ
НА РОСТ POPULUS TREMULA L.
Изучение растений Populus tremula позволило установить, что первыми (в начале июня) начинают рост побеги второго порядка, спустя 2–4 суток появляются листья, а через 10–13 суток начинается деление камбиальных клеток ствола. Кульминация прироста побегов наблюдается в середине июня – начале июля, листьев – в начале-середине июля, ствола – в середине июля. Рост побегов заканчивается в конце июля, листьев – в середине августа, ствола – в конце августа. Наибольшая продолжительность роста характерна для листьев – 65–75 сут., у стволов она составляет 65–67 суток, у побегов – 38–49 суток.
Температурный режим воздуха заметно влияет на рост побегов, листьев и стволов Populus tremula. Рост побегов начинается при среднесуточной температуре воздуха +12.5–+13.7 °С, листьев – при +12.8–+17.2 °С, стволов – при +12.2–+13.3 °С. Температура воздуха оказывает воздействие на интенсивность роста побегов, листьев и стволов в течение всего периода их формирования. Кульминация прироста побегов происходит при температуре воздуха выше +15 °С, листьев – выше +18 °С, стволов – выше +19 °С. Прекращение роста побегов отмечается при падении температуры ниже +10 °С, листьев – ниже 15 °С, стволов – ниже 16 °С.
В результате исследований установлено, что продолжительность солнечного сияния оказывает слабое положительное влияние на интенсивность ростовых процессов, а атмосферные осадки и влажность воздуха – слабое отрицательное воздействие.
ГЛАВА 6. ВЛИЯНИЕ КЛИМАТИЧЕСКИХ ФАКТОРОВ
НА РОСТ ALNUS INCANA (L.) MOENCH.
Как показали исследования растений Alnus incana, первыми (в конце мая) начинают рост побеги второго порядка, спустя 1–2 суток появляются листья, а через 19–20 суток начинается формирование древесины ствола. Кульминация прироста побегов наблюдается в конце июня – начале июля, листьев – в конце июня – середине июля, ствола – в середине июля. Окончание роста побегов отмечается во второй половине июля, листьев и ствола – в середине августа. Наибольшая продолжительность роста характерна для листьев – 79–80 суток, у стволов она составляет 57–63 суток, у побегов – 50–52 суток.
Исследованиями установлено, что особенности роста деревьев этого вида в основном связаны с динамикой температуры воздуха. Рост побегов начинается при повышении среднесуточной температуры воздуха до +8.0–+9.1 °С, листьев – до +9.4–+9.8 °С, стволов – до +14.6–+16.8 °С. Температура воздуха оказывает влияние на интенсивность роста побегов и стволов в течение всего периода их формирования,
а на рост листьев – только в период кульминации прироста. Прирост побегов кульминирует при температуре воздуха выше +19 °С, листьев – выше +17 °С, стволов – выше +18 °С. Прекращение роста побегов наблюдается при падении температуры ниже +18 °С, хвои – ниже 12 °С, стволов – ниже 13 °С.
Продолжительность солнечного сияния оказывает слабое положительное влияние на интенсивность ростовых процессов, а атмосфер-
ные осадки и влажность воздуха – слабое отрицательное воздействие.
ЗАКЛЮЧЕНИЕ
Результаты исследований, обработанные с помощью математических методов позволили получить представление о закономерностях роста растений некоторых лесообразующих видов в условиях Карелии (подзона северной тайги), а также установить зависимость этих процессов от главнейших экологических факторов.
Изучение сезонного роста растений Picea abies, Betula pubescens, Populus tremula и Alnus incana показало, что его особенности определяются биологией вида, динамикой экологических факторов в течение вегетационного периода, различиями почвенно-грунтовых условий, возрастом деревьев и их положением в пологе древостоя. Эти факторы
оказывают влияние на сроки начала, кульминации и окончания роста,
а также на интенсивность и продолжительность формирования стволов, побегов второго порядка и фотосинтезирующего аппарата. Выявлены формы зависимости между интенсивностью продукционных процессов и факторами внешней среды, построены графики, отражающие связь сезонного роста побегов, хвои, листьев и стволов деревьев с динамикой экологических факторов.
В ходе формирования фитомассы древостоев обнаружена определённая последовательность в сроках начала, кульминации и окончания роста побегов, листового аппарата и стволов. Первыми в рост трогаются побеги, затем появляется молодая хвоя и начинается деление клеток камбия стволов. Наибольшая продолжительность роста характерна для хвои, листьев и стволов (70–90 суток), а наименьшая – для побегов (45–60 суток).
Установлено, что 70–80% годичного прироста побегов, хвои, листьев и стволов формируется в первую половину периода их роста. Различия в величине годичного прироста органической массы древостоев, вызванные влиянием экологических факторов, на 65–80% связаны с неодинаковой интенсивностью роста и лишь на 20–35% − с разницей в продолжительности их формирования.
Главными причинами сравнительно медленного роста растений лесообразующих видов в районе исследований являются довольно прохладный и короткий вегетационный период, недостаток минерального питания и застойный режим увлажнения.
Обнаружено, что интенсивность и продолжительность роста побегов, ассимиляционного аппарата, и стволов растений исследуемых видов в процессе онтогенеза, как правило, снижается, что соответствующим образом отражается и на величине их годичного прироста.
Установлено, что с усилением степени угнетенности дерева и ухудшением почвенно-грунтовых условий продолжительность деятельности меристематических тканей уменьшается, а ее интенсивность снижается. Это приводит к соответствующему уменьшению величины годичного прироста побегов, хвои, листьев и стволов и ухудшению качества древесины ствола.
Выяснилось, что основные особенности роста растений определяются не только биологическими особенностями вида, но и погодными условиями вегетационного периода. Среди изучаемых экологических факторов, оказывающих влияние на рост растений, главную роль играет температура воздуха. Определены значения среднесуточной температуры воздуха и суммы положительных температур, необходимых для начала роста и кульминации прироста побегов, ассимиляционного аппарата и стволов Picea abies, Betula pubescens, Populus tremula и Alnus incana.
На основе проведённых исследований и анализе данных можно сделать следующие выводы:
1. Рост побегов, хвои, листьев и стволов у Picea abies, Betula pubescens, Populus tremula и Alnus incana происходит в определённой последовательности. Первыми трогаются в рост побеги, затем появляются хвоя и листья, после чего начинается деление клеток камбия ствола. Кульминация и прекращение роста происходит в следующем порядке: побеги, стволы, хвоя и листья. Наибольшая продолжительность роста характерна для листового аппарата, а наименьшая – для побегов.
2. Из всех изученных климатических факторов температурный режим воздуха оказывает наибольшее влияние на процессы роста растений лесообразующих видов. Сроки начала, кульминации и окончания роста побегов, листового аппарата и стволов растений того или иного вида связаны с особыми температурными условиями.
3. Темпы формирования побегов, хвои, листьев и стволов растений изученных видов в основном определяются погодными условиями докульминационного периода. Падение температуры воздуха ниже +12–13 °С отрицательно сказывается на интенсивности деятельности меристематических тканей.
4. Рост побегов, фотосинтезирующего аппарата и стволов деревьев в лучших почвенно-грунтовых условиях начинается и заканчивается позже. Поэтому продолжительность формирования побегов, хвои и стволов в ельнике черничном больше, чем в ельнике сфагновом, в среднем соответственно на 11, 19 и 12 суток.
5. В лучших условиях местопроизрастания интенсивность ростовых процессов заметно усиливается. В период кульминации суточный прирост побегов в ельнике черничном в среднем достигает 1.8, хвои – 1.1 и стволов – 3.3 мм, а в ельнике сфагновом – соответственно 1.0, 0.6 и 2.9 мм.
6. Более интенсивный и продолжительный рост дерева в лучших условиях местопроизрастания приводит к увеличению его годичного прироста. В ельнике черничном по сравнению с ельником сфагновым длина побегов в среднем больше на 89 %, хвои – на 85 %, ширина годичного кольца стволов – на 45 %.
7. С улучшением положения дерева в пологе леса ростовые процессы идут продолжительнее и эффективнее. Продолжительность формирования древесины у деревьев II–III классов роста по сравнению с деревьями IV–V классов увеличивается в среднем на 7 суток. Величина годичного радиального прироста у деревьев II–III классов по сравнению с деревьями IV–V классов оказалась больше в среднем на 95%. В связи с этим при проведении рубок ухода целесообразно убирать отставшие в росте деревья как неперспективные, оставляя на корню толстые, более продуктивные деревья.
8. Динамика сезонного роста побегов, листового аппарата и стволов в процессе онтогенеза несколько меняется. Интенсивность роста стволов с возрастом дерева уменьшается, хвои – увеличивается, а у побегов наибольшей величины она достигает в приспевающем древостое. Годичный прирост ствола в молодняке в среднем составляет 3.0 мм, в приспевающем – 3.2 мм, в спелом – 2.9 мм, хвои – соответственно 23.0, 23.1, и 24.3 мм, побегов – 47.7, 49.1, 47.5 мм.
9. Влияние исследуемых экологических факторов на величину годичного прироста побегов, хвои, листьев и стволов реализуется в основном через изменение интенсивности, а не продолжительности их формирования.
10. Атмосферные осадки и относительная влажность воздуха оказывают слабое отрицательное, а продолжительность солнечного сияния – положительное воздействие на динамику прироста побегов, ассимиляционного аппарата и стволов растений изучаемых видов.
11. Наиболее качественная древесина у растений Picea abies формируется в ельнике брусничном и ельнике черничном. По сравнению с ельником сфагновым и ельником болотно-травяным доля поздней древесины в этих типах леса в среднем на 15–20% выше, а толщина оболочек поздних трахеид – на 23–45 % больше.
12. Качество древесины растений Picea abies у господствующей части древостоя выше, чем у деревьев угнетенных. Это выражается в увеличении доли участия поздней древесины в формировании годичного кольца на 12%, а также возрастании толщины оболочек трахеид на 10–15%.
Установленные закономерности в сезонном росте растений лесообразующих видов, произрастающих в северной Карелии, позволяют дать ряд практических рекомендаций:
1. В еловых древостоях должны проводиться лесохозяйственные мероприятия, направленные на потепление фитоклимата (рубки ухода) в вегетационный период, что активизирует деятельность меристематических тканей.
2. Лесохозяйственные мероприятия по повышению продуктивности еловых лесов эффективнее проводить в весенний период и в первую очередь в молодых древостоях в связи с тем, чтобы использовать их наивысшую энергию роста в наиболее благоприятное для этого время.
3. С понижением класса роста и развития деревьев интенсивность
и длительность роста вегетативных органов лесообразующих видов существенно снижаются, что следует учитывать при проведении рубок ухода.
СПИСОК ОПУБЛИКОВАННЫХ РАБОТ
ПО ТЕМЕ ДИССЕРТАЦИИ
1. Кищенко И. Т., Вантенкова И. В. Сезонный рост Picea abies (L.) Karst. в северной Карелии // Структурно-функциональные особенности биосистем севера (особи, популяции, сообщества): Мат. конференции, посвящённой 65-летию Петрозаводского государственного университета и эколого-биологического факультета (26–30 сентября 2005 года). Ч. II. – Петрозаводск, 2005. – С. 219–222.
2. Вантенкова И. В., Кищенко И. Т. Сезонный рост вегетативных органов ели европейской в разных типах леса северной Карелии // Экологические проблемы севера: Межвузовский сборник научных трудов. Вып. 8. – Архангельск, 2005. – С. 144–146.
3. Вантенкова И. В., Кищенко И. Т. Сезонный рост Picea abies (L.) Karst. в древостоях разного возраста северной Карелии // Экологи-
ческие проблемы севера: Межвузовский сборник научных трудов. Вып. 8. – Архангельск, 2005. – С. 151−153.
4. Вантенкова И. В. Влияние экологических факторов на сезонный рост Picea abies (L.) Karst. в природных комплексах северной Карелии // Вузовская школа – региону: Мат. IV Всероссийской научно-технической конференции (21 февраля 2006 года). Т. I. – Вологда, 2006. – С. 393–395.
5. Вантенкова И. В. Влияние экологических факторов на сезонный рост лесообразующих видов в северной Карелии // Севергеоэкотех–2006: Мат. VII международной молодёжной научной конференции (22–24 марта 2006 года). Ч II. – Ухта, 2006. – С. 294–297.
6. Вантенкова И. В. Влияние факторов окружающей среды на сезонный рост Picea abies (L.) Karst. в северной Карелии // Биология − наука XXI века: Мат. X Пущинской школы-конференции молодых учёных, посвящённой 50-летию Пущинского научного центра РАН (17–21 апреля 2006 года). – Пущино, 2006. – С. 261.
7. Вантенкова И. В. Рост вегетативных органов Picea abies (L.) Karst. в северной Карелии // Флора и растительность Сибири и Дальнего Востока: Мат. IV Российской конференции (18–20 апреля 2006 года). – Красноярск, 2006. – С. 116–120.
8. Кищенко И. Т., Вантенкова И. В. Сезонный рост Picea abies (L.) Karst. в северной Карелии // Устойчивость экосистем и проблема сохранения биоразнообразия на севере: Мат. Международной конференции (26–30 августа, 2006). Т. I. – Кировск, 2006. – С. 91–95.
9. Вантенкова И. В., Кищенко И. Т. Формирование годичного кольца Picea abies L. (Karst.) в северной Карелии // Экологические проблемы севера: Межвузовский сборник научных трудов. Вып. 9. – Архангельск, 2006. – С. 20–22.
10. Кищенко И. Т., Вантенкова И. В. Влияние экологических факторов на сезонный рост Picea abies (L.) Karst. в северной Карелии // Экология. 2007. – № 2. – С. 111–117.
Подписано к печати 10.09.07. Формат 60х84 1/16.
Бумага офсетная. Уч.-изд. л. 1.
Тираж 100 экз. Изд. № 212
Государственное образовательное учреждение
высшего профессионального образования
Петрозаводский государственный университет
Отпечатано в типографии Издательства ПетрГУ
![]()
185910 Петрозаводск, пр. Ленина, 33
