

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

На правах рукописи
БОГОСЛОВСКИЙ ВАСИЛИЙ ВАСИЛЬЕВИЧ
ПРОГНОЗИРОВАНИЕ ПРОДУКТИВНОСТИ, АДАПТАЦИОННЫХ СПОСОБНОСТЕЙ ПОРОД И ГИБРИДОВ ТУТОВОГО ШЕЛКОПРЯДА ПО ФЕРМЕНТАМ
И БЕЛКОВЫМ СПЕКТРАМ
06.02.01 – разведение, селекция, генетика и воспроизводство сельскохозяйственных животных
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Ставрополь – 2009
![]() Диссертация выполнена в ГНУ Республиканская научно-исследовательская станция шелководства Россельхозакадемии
Диссертация выполнена в ГНУ Республиканская научно-исследовательская станция шелководства Россельхозакадемии
Научный руководитель: доктор биологических наук
Официальные оппоненты: доктор биологических наук, профессор
член-корр. РАСХН
доктор сельскохозяйственных наук,
профессор
Ведущая организация: ГНУ Научно-исследовательский институт
пчеловодства Россельхозакадемии
Защита диссертации состоится «3» июля 2009 года в 900 часов на заседании диссертационного совета Д 006.078.01 при Ставропольском научно-исследовательском институте животноводства
и кормопроизводства Ставрополь, пер. Зоотехнический, 15, тел./
С диссертацией можно ознакомиться в библиотеке ГНУ Ставропольского научно – исследовательского института животноводства и кормопроизводства.
Автореферат диссертации разослан «___» июня 2009 года и размещен на официальном сайте института – http:www. ***** «___» июня 2009 года.
Ученый секретарь
диссертационного совета
Общая характеристика работы
1.1 Актуальность темы. Одной из основных задач современной генетики и селекции тутового шелкопряда является создание высокопродуктивных пород и гибридов. Традиционные методы оценки эффективности различных гибридных комбинаций предполагают проведение длительных по времени и трудозатратных в исполнении экспериментальных выкормок. В связи с этим разработка надежных и быстрых в исполнении тестов с использованием современных достижений в области биохимии, молекулярной генетики для раннего прогнозирования продуктивности гибридов и отбор наиболее эффективных для дальнейшего селекционного процесса является актуальной (, , 2003; , 2006; и соавт., 2007).
В ряде исследований доказана возможность использования активности различных ферментов – амилаз, фосфатаз, аминотрансаминаз, эстераз в тканях и органах куколок, личинок тутового шелкопряда для прогнозирования эффективных вариантов и отбора наиболее гетерозисных комбинаций ( и соавт., 1974; 1987; , , 1979; , 1983; , 1991; , 2005). В то же время известно, что развитие зародыша определяется интенсивностью процесса расщепления запасных белков грены (яйца), что обеспечивается специфическими протеолитическими ферментами (Liu X. D. et al., 1996). Однако исследований по установлению связи между их активностью и хозяйственно полезными показателями у гибридов тутового шелкопряда для раннего прогнозирования продуктивности выполнено недостаточно.
Не менее важной проблемой в шелководстве является повышение устойчивости пород и гибридов к неблагоприятным условиям среды и заболеваниям. Метаболизм тутового шелкопряда, являющегося пойкилотермным организмом, в значительной степени зависит от факторов внешней среды, особенно температуры, которая во многом определяет скорость протекания биохимических процессов (, 1975). Поэтому воздействие близких к пороговым значениям температур может вызывать изменение внутриклеточных механизмов и биолого-технологических показателей ( и соавт., 1995; Суханов, С. В., 1999). В этом аспекте определенный интерес представляет уровень ферментативной активности, поскольку он может служить критерием оценки физиологического состояния организма (и соавт., 1995; и соавт., 1996). Однако возможность использования активности рибонуклеаз и дезоксирибонуклеаз для прогнозирования устойчивости тутового шелкопряда к тепловым воздействиям и вирусным инфекциям до сих пор изучена недостаточно.
В связи с этим поиск биохимических тестов прогнозирования продуктивности и резистентности у тутового шелкопряда является актуальным и практически востребованным.
1.2 Цель и задачи исследований. Целью работы являлась разработка методов прогнозирования продуктивности гибридов, адаптационных качеств и идентификации пород тутового шелкопряда по ферментативной активности и спектрам растворимых белков.
Для достижения цели были поставлены следующие задачи:
1. Изучить сопряженность активности протеолитических ферментов грены с хозяйственно ценными признаками у тутового шелкопряда и разработать биохимический метод тестирования продуктивности гибридного поколения на эмбриональной стадии развития.
2. Изучить влияние стрессов различной природы на продуктивные показатели тутового шелкопряда, белковый профиль, активность рибонуклеаз гемолимфы для прогнозирования адаптационных возможностей тутового шелкопряда.
3. Изучить степень гетерогенности растворимых белков грены и на ее основе разработать метод идентификации морфологически схожих пород тутового шелкопряда.
1.3 Научная новизна исследований. На основе комплексного исследования протеолитических ферментов грены тутового шелкопряда выявлена сопряженность их активности с рядом хозяйственно ценных признаков и впервые разработан биохимический метод отбора перспективных хозяйственно полезныминых гибридов на ранней стадии развития.
Впервые изучено влияние пороговых температурных факторов и инфекционного стресса на количественно-качественное изменение электрофоретической подвижности растворимых белковых фракций гемолимфы и активность рибонуклеаз. Показано, что их активность может служить показателем прогнозирования адаптационных качеств у тутового шелкопряда.
Впервые изучена возможность использования растворимых белков грены для идентификации морфологически трудноразличимых пород тутового шелкопряда.
1.4 Практическая значимость и реализация результатов исследований.
Для практического использования разработаны:
- биохимический метод прогнозирования продуктивности гибридов тутового шелкопряда на ранней стадии эмбриогенеза;
- метод тестирования адаптационных способностей пород тутового шелкопряда на основе сопоставления спектров растворимых белков и активности рибонуклеаз гемолимфы;
- метод идентификации трудноразличимых пород тутового шелкопряда по наборам растворимых белков грены.
Использование методов прогнозирования продуктивности гибридов и отбора наиболее перспективного селекционного материала позволило сократить затраты одного выкормочного сезона на 470,0 тыс. в год.
Ежегодный экономический эффект от использования метода тестирования адаптационных способностей тутового шелкопряда составил 442,5 тыс. руб. за один экспериментальный сезон.
1.5 Апробация работы. Основные положения диссертации докладывались на ежегодных заседаниях Ученого Совета ГНУ Республиканской научно-исследовательской станция шелководства Россельхозакадемии (РНИСШ, г. Железноводск), выездных сессиях Отделения зоотехнии Россельхозакадемии в 2000 – 2006 гг., международной научно-практической конференции (Оренбург, 2008), опубликованы в открытой научной печати.
1.6 Связь темы с планом научных исследований. Диссертационная работа выполнена в рамках Программы фундаментальных и приоритетных прикладных исследований по научному обеспечению развития агропромышленного комплекса РФ ( гг.; гг.; гг.) и плана НИОКР ГНУ РНИСШ по заданиям: 01.02.01 «Разработать на основе биохимических тестов метод ранней довыкормочной диагностики гетерозисной силы исходного селекционного материала на ранних стадиях эмбриогенеза тутового шелкопряда для выявления гибридных сочетаний с высокой комбинационной ценностью»; 01.01.01 «Усовершенствовать метод оценки адаптационных возможностей пород тутового шелкопряда на основе сопоставления белковых спектров и активности ферментов»; 01.01.02 «Разработать универсальные биохимические тесты для породной идентификации тутового шелкопряда».
1.7 Основные положения, выносимые на защиту:
1. Различия в активности протеолитических ферментов у исходных пород тутового шелкопряда и их гибридов могут служить биохимическим тестом прогнозирования гетерозисных комбинаций.
2. Воздействие пороговых температурных факторов и инфекционного стресса значительно снижает показатели продуктивности, изменяет электрофоретическую подвижность растворимых белковых фракций и активность рибонуклеаз гемолимфы гусениц у тутового шелкопряда.
3. Активность рибонуклеаз может служить дополнительным критерием оценки адаптационных способностей тутового шелкопряда на стадии гусеницы.
4. Метод изоэлектрофокусирования растворимых белков грены позволяет получить уникальные спектры для идентификации морфологически трудноразличимых пород тутового шелкопряда.
1.7 Публикация результатов исследований. По материалам диссертационной работы опубликовано 9 научных работ, в том числе 2 статьи в научных изданиях из перечня ВАК.
1.8 Объем и структура диссертации. Диссертация изложена на 123 страницах компьютерного текста, содержит 18 таблиц и 11 рисунков, включает введение, обзор литературы, методическую часть, результаты исследований, выводы и предложения производству, список литературы, насчитывающий 206 источников, в т. ч. 61 зарубежных.
2 МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
Исследования проводились в лаборатории шелководства РНИИСШ в период 2000 – 2008 гг. согласно схеме, представленной на рисунке 1.
В опыте по изучению активности протеолитических ферментов ( гг.) материалом для исследований служила грена (яйца) четырех пород: Кавказ 1 (К-1), Белококонная 1 (Б-1), Шелковистая 4 (Ш-4), Мзиури 2 (Мз-2) и их гибридов: трех прямых – К-1хШ-4; К-1хМз-2; Б-1хШ-4 и трех обратных – Ш-4хК-1; Мз-2хК-1; Ш-4хБ-1.
Протеолитическую активность протеиназ в 4 и 9 суточной грене постдиапаузного развития (Е1, бластокинез и Е3, стадия почернения гусеницы) и их гетерогенность определяли по методу Ансона (Anson, 1939) в модификации и соавт. (1987) в трехкратной повторности на кафедре биохимии Московского педагогического государственного университета.
Активность трипсино - и химотрипсиноподобных протеиназ определяли спектрофотометрическим методом, используя в качестве субстратов растворы N-бензоил–DL-аргинин–п–нитроанилида (BAPNA) и сукцинил-D, L-фенилаланин-п–нитроанилида (SUPHEPA) из расчета 5 мг/мл. Диапазон рН от 2,2 до 11,0 единиц обеспечивался различной растворимостью при определенных значениях pH двух природных субстратов – гемоглобина и казеина.
Подклассовую принадлежность ферментов определяли по соответствующим ингибиторам: для сериновых протеиназ – фенилметилсульфонилфторид; цистеиновых – n-хлормеркурибензоат.
В эксперимент по изучению адаптационных способностей было отобрано шесть пород трех контрастных групп: российской – Кавказ 13 (К-13), ПС-5; китайской – Белококонная 2 (Б-2), Сычуань; японской – Японская 107 (Я-107), Даидзо. На гусениц V возраста воздействовали стрессовыми факторами: индуцированным вирусом ядерного полиэдроза (ВЯП), повышенной сублетальной (+40ºС), пониженной (+4ºС … +5ºС) температурами.
Индуцирование вирусом гусениц вызывали путем скармливания сброженного листа шелковицы. Температурные стрессы вызывали путем помещения гусениц в термостат и холодильную камеру при указанных температурах соответственно на два и шесть часов. Через пять минут после описанных воздействий от каждой партии отбирали по 25 гусениц консервации гемолимфы. Контролем служили гусеницы тех же пород, выращенные в оптимальных условиях.
Фракционный состав растворимых белков определяли методом электрофореза в полиакриламидном геле (7,5% ПААГ). Количественное содержание белковых фракций – денситометрированием гелевых пластин с использованием соответствующей компьютерной программы.
Определение РНКазной и ДНКазной активности проводили спектрометрически по модифицированной методике Кога с соавт. (1987).
|
|


|
|
|

|
|
|
|
|
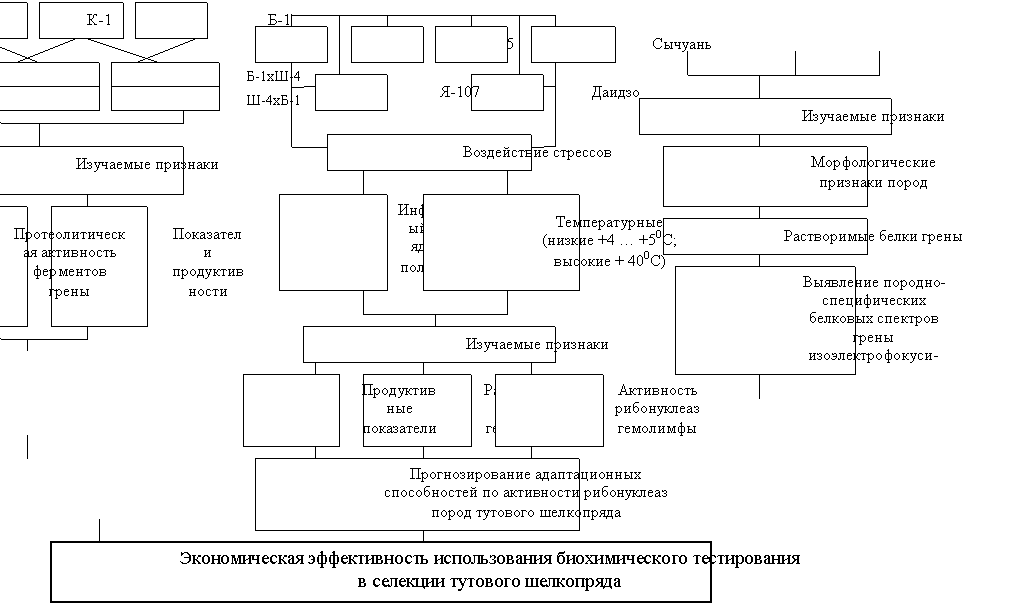

Объектом исследования служила грена трех пород – Пятигорская станция (ПС-5), Белококонная 2 (Б-2), HSC-90 второго дня постдиапаузного развития. Инкубацию грены осуществляли в стандартных условиях (температура 23-240С, относительная влажность 70%).
Для получения растворимых белков навеску грены (1,0 г) и гемолимфы (0,2 г) гомогенизировали при охлаждении (0 + 40С), белки экстрагировали 2 мл 0,15М раствора NaCl в течение 15 мин., центрифугировали при 2 000 об/мин в течение 30 мин.
К полученной надосадочной жидкости добавляли 5% додецилсульфата натрия (SDS) и 5% β-меркаптоэтанола и выдерживали на водяной бане при 900С в течение 5 минут.
Проведение экспериментальных выкормок и определение биолого-хозяйственных и технологических показателей пород и гибридов тутового шелкопряда осуществляли в соответствии с «Методикой проведения экспериментальных выкормок тутового шелкопряда» (Росшелкстанция, 1990). Учитывали: биолого-хозяйственные (жизнеспособность гусениц, средняя масса кокона, выход коконов с 1 г гусениц однодневного возраста, шелковая оболочка сырого кокона) и технологические (шелконосность сухих коконов, разматываемость оболочки сухих коконов, выход шелка-сырца сухих коконов, длина непрерывно разматываемой нити, расчетный выход шелка-сырца) признаки.
Статистическую обработку экспериментально полученных данных проводили, используя руководство (1983).
3 РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
3.1 Протеолитическая активность протеиназ в грене
родительских пород и гибридов тутового шелкопряда
Исследование активности протеиназ позволило установить, что в состав растворимых белков грены исследованных пород и гибридов входит сложный комплекс протеолитических ферментов, функционирующих при различных значениях рН.
Исследование их подклассовой принадлежности по специфичности к модельным субстратам позволило определить цистеиновые (рН 3,0; 3,6; 6,2 и 8,6) и сериновые (рН 6,2; 7,2; 8,6 и 9,0) протеиназы. При этом цистеиновые протеиназы с оптимумом рН 3,0; 3,6; 6,2 и 8,6 являлись папаиноподобными, сериновые с оптимумом рН 7,2 – химотрипсиноподобными, а с оптимумом рН 9,0 – трипсиноподобными протеиназами.
Сопоставление уровня их активности у исходных пород и гибридов выявило, что у гибридов в сравнении с исходными родителями при одних значения рН отмечалось повышение энзиматической активности, при других – понижение (табл. 1, 2).
Таблица 1 - Разница в активности папаиноподобных протеиназ в грене родительских пород
и гибридов тутового шелкопряда, %
Породы, гибриды | Протеолитическая активность, усл. ед/мг белка | |||||||||||
Папаиноподобные | ||||||||||||
рН 3,0 | рН 3,6 | рН 6,2 | рН 8,6 | |||||||||
I | II | III | I | II | III | I | II | III | I | II | III | |
К-1, Ш-4 | 60,0 ±3,47 | 21,0 ±0,75 | 10,0 ±0,02 | 28,5 ±0,09 | ||||||||
К-1хШ-4 | 142,3*** ±7,48 | 57,8 | 30,0*** ±0,09 | 30,0 | 28,8*** ±2,02 | 65,2 | 18,6 ±1,74 | -34,7 | ||||
Ш4хК-1 | 134,0*** ±8,34 | 55,2 | 60,0*** ±5,12 | 65,0 | 14,1**±1,17 | 29,4 | 39,0*** ±2,37 | 26,9 | ||||
К-1, Мз-2 | 35,5 ±2,01 | 9,0 ±0,06 | 6,0 ±0,01 | 16,5 ±1,12 | ||||||||
К-1хМз-2 | 55,0** ±4,04 | 35,4 | 20,0*** ±1,78 | 55,0 | 0 | -100 | 45,8*** ±3,56 | 63,9 | ||||
Мз-2хК1 | 46,0* ±2,98 | 22,8 | 22,0*** ±1,46 | 59,1 | 0 | -100 | 0 | -100 | ||||
Б-1, Ш-4 | 65,5 ±4,45 | 20,5 ±0,05 | 30,0 ±0,16 | 12,0 ±0,05 | ||||||||
Б-1хШ-4 | 87,0* ±4,35 | 24,7 | 83,0*** ±5,16 | 75,3 | 13,0 ±0,08 | -56,6 | 39,0*** ±3,07 | 69,2 | ||||
Ш-4хБ-1 | 88,0* ±3,98 | 25,5 | 84,0***± 4,16 | 75,5 | 15,0 ±1,17 | -50,0 | 41,0*** ±2,46 | 70,7 |
Так, активность папаиноподобных протеиназ с оптимумом рН 3,0 и 3,6 у всех изученных гибридов была достоверно выше среднеродительского показателя. Разница колебалась от 22,8% (Р<0,05) до 75,5% (Р<0,001).
Превышение активности пептидогидролаз с оптимумом рН 8,6 отмечено у прямого и обратного гибридов Б-1 и Ш-4, а также у прямого гибрида К-1 и Мз-2 и обратного – Ш-4 и К-1 (от 26,9 до 70,7%, Р<0,001).
В активности протеиназ с оптимумом рН 6,2 положительная разница выявлена только у прямого и обратного гибридов К-1 и Ш-4 на 29,4% (Р<0,01) и 65,2% (Р<0,001).
Таблицы 2 – Разница в активности химо - и трипсиноподобных протеиназ в грене родительских пород и гибридов тутового шелкопряда, %
Породы, гибриды | Протеолитическая активность, усл. ед/мг белка | |||||
Химотрипсиноподобные | Трипсиноподобные | |||||
рН 9,0 | рН 7,2 | |||||
I | II | III | I | II | III | |
К-1; Ш-4 | 812,5±38,7 | 29,0±0,13 | ||||
К-1 х Ш-4 | 53,6±4,12 | -93,4 | 0 | -100 | ||
Ш-4 х К-1 | 487,5±18,9 | -40,0 | 0 | -100 | ||
К-1; Мз-2 | 619,0±31,6 | 29,5±0,15 | ||||
К-1 х Мз-2 | 313,6±18,9 | -49,3 | 41,6 ±2,18*** | 41,0 | ||
Мз-2 х К-1 | 613,2±29,4 | -0,93 | 139,0 ±5,36*** | 371,3 | ||
Б-1; Ш 4 | 575,0±28,5 | 97,0±7,11 | ||||
Б-1 х Ш-4 | 76,5±4,16 | -86,6 | 0 | -100 | ||
Ш-4 х Б-1 | 143,7±6,23 | -75,0 | 0 | -100 |
Примечание: I – среднеродительский показатель; II – гибрид;
III – разница между показателем протеолитической активности
у гибрида и среднеродительским показателем, %
В то же время у всех полученных гибридов отмечено снижение активности химотрипсиноподобных ферментов с оптимумом рН 9,0 (от 0,9% до 93,4%, Р<0,001), у гибридов К-1 и Мз-2, Б-1 и Ш-4 - папаиноподобных протеиназ с оптимумом рН 6,2 (на 50,0-100,0%, Р<0,001), у гибрида Мз-2 хК1 - с оптимумом рН 8,6 и у гибридов К-1 и Ш-1, Б-1 и Ш-4 - с оптимумом рН 7,2 (на 100,0%, Р<0,001).
Таким образом, по превышению в активности протеолитических ферментов выделялись гибриды Ш-4хК-1 и К-1хМз-2, что позволяет выделить их как наиболее перспективные гибридные сочетания. Наибольшей информативностью в плане прогнозирования гетерозиса у гибридов, на наш взгляд, являются папаиноподобные протеиназы с оптимумом рН 3,0, 3,6 и 8,6, поскольку у всех гибридов выявлено превышение над среднеродительскими показателями.
Таблица 3 – Сравнение биолого-хозяйственных показателей родительских пород и гибридов тутового шелкопряда, %
Породы, гибриды | Биолого-хозяйственные показатели | |||||||||||
Жизнеспособность гусениц, % | Средняя масса кокона, г | Выход коконов с 1 г гусениц однодневного возраста, кг | Шелковая оболочка сырых коконов, % | |||||||||
I | II | III | I | II | III | I | II | III | I | II | III | |
К-1; Ш-4 | 92,2 ±0,72 | 2,0 ±0,01 | 3,71 ±0,02 | 20,7 ±0,49 | ||||||||
К-1 х Ш-4 | 95,5* ±0,72 | 3,4 | 2,1*** ±0,01 | 5,0 | 3,99*** ±0,04 | 7,5 | 21,1 ±0,73 | 1,9 | ||||
Ш-4 х К-1 | 94,1 ±0,81 | 1,9 | 2,2*** ±0,01 | 10,0 | 3,87** ±0,05 | 4,3 | 21,1 ±0,49 | 1,9 | ||||
К-1; Мз-2 | 92,2 ±0,54 | 2,1 ±0,01 | 3,75 ±0,02 | 19,9 ±0,42 | ||||||||
К-1 х Мз-2 | 93,7 ±0,92 | 1,6 | 2,1 ±0,01 | 0 | 4,18*** ±0,05 | 11,4 | 20,5 ±0,62 | 3,0 | ||||
Мз-2 х К-1 | 93,8 ±0,64 | 1,7 | 2,0 ±0,01 | 5,0 | 4,01*** ±0,04 | 6,9 | 20,0 ±0,56 | 0,5 | ||||
Б-1; Ш 4 | 91,2 ±0,63 | 2,1 ±0,01 | 3,57 ±0,02 | 19,9 ±0,50 | ||||||||
Б-1 х Ш-4 | 90,0 ±0,77 | 1,8 | 2,1 ±0,01 | 0 | 3,89*** ±0,02 | 8,9 | 19,9 ±0,51 | 0 | ||||
Ш-4 х Б-1 | 91,0 ±0,59 | 2,9 | 2,0 ±0,01 | 5,0 | 3,91*** ±0,03 | 9,5 | 20,6 ±0,49 | 3,5 |
Таблица 4 – Сравнение технологических показателей родительских пород и гибридов тутового шелкопряда, %
Породы, гибриды | Технологические показатели | ||||||||||||||
Шелконосность сухих коконов, % | Разматываемость оболочки сухих коконов, % | Выход шелка-сырца сухих коконов, % | Длина непрерывно разматываемой нити, м | Расчетный выход шелка-сырца, кг | |||||||||||
I | II | III | I | II | III | I | II | III | I | II | III | I | II | III | |
К-1; Ш-4 | 42,1 ±0,68 | 87,9 ±0,42 | 37,8 ±0,49 | 1150 ±5,40 | 0,560 ±0,17 | ||||||||||
К-1 х Ш-4 | 48,8*** ±0,82 | 15,9 | 89,7* ±0,34 | 2,0 | 40,0* ±0,27 | 5,8 | 1159 ±6,17 | 0,7 | 0,638* ±0,02 | 13,9 | |||||
Ш-4 х К-1 | 48,9*** ±0,83 | 16,1 | 89,4* ±0,43 | 1,7 | 40,4* ±0,32 | 6,8 | 1147 ±5,96 | -0,3 | 0,630* ±0,02 | 12,5 | |||||
К-1; Мз-2 | 41,4 ±0,53 | 88,75 ±0,43 | 37,6 ±0,48 | 1108 ± | 0,561 ±0,18 | ||||||||||
К-1 х Мз-2 | 44,8** ±0,64 | 8,2 | 91,6** ±0,51 | 3,2 | 39,8* ±0,26 | 5,8 | 1089 ±4,83 | -1,7 | 0,688** ±0,01 | 22,6 | |||||
Мз-2 х К-1 | 43,6* ±0,71 | 5,3 | 90,6* ±0,48 | 2,1 | 38,9 ±0,41 | 3,4 | 1078 ±4,64 | -2,7 | 0,623* ±0,02 | 11,0 | |||||
Б-1; Ш-4 | 42,1 ±0,68 | 87,0 ±0,41 | 36,1 ±0,47 | 1072 ±5,10 | 0,530 ±0,12 | ||||||||||
Б-1 х Ш-4 | 42,2 ±0,64 | 0,2 | 90,5** ±0,47 | 4,0 | 40,2*** ±0,28 | 11,3 | 1123*** ±5,03 | 4,7 | 0,632*** ±0,01 | 19,2 | |||||
Ш-4 х Б-1 | 41,0 ±0,59 | -2,6 | 89,2* ±0,36 | 2,5 | 39,4*** ±0,31 | 9,1 | 1142*** ±4,48 | 6,5 | 0,618** ±0,02 | 16,6 |
Примечание: * – Р<0,05; ** – Р<0,01;*** – Р<0,001;
I – среднеродительский показатель; II – показатели гибридов;
III – разница между гибридом и среднеродительским показателем, в % ;
С целью подтверждения сделанного предположения, а также выявления возможной связи между протеолитической активностью ферментов и продуктивными качествами тутового шелкопряда были изучены биолого-хозяйственные и технологические показатели гибридов и проведено их сопоставление с показателями родительских пород (табл. 3, 4).
В шести гибридных комбинациях выявлена высокодостоверная разница (от 4,4 до 11,4%) по выходу кокона с 1 г гусениц, выходу шелка-сырца сухих коконов (от 3,4 до 4,3%) и его расчетному выходу (12,5 – 22,6%). Несколько меньшая, но во всех случаях положительная разница установлена по жизнеспособности гусениц (на 1,7 – 3,4%), проценту шелковой оболочки (на 0,5 – 3,5%), разматываемости оболочки сухих коконов (на 1,7 – 4,0%).
По таким показателям, как шелконосность сухих коконов и длина непрерывно разматываемой нити у одних гибридов установлено превышение над родителями, у других – понижение. Так, у гибридных сочетаний К-1 хШ-4, Ш-4хК-1 и К-1хМз-2, Мз-2 хК-1 шелконосность сухих коконов была выше на 5,3 – 16,1%, в то время как у Б-1хШ-4 и Ш-4хБ-1 отмечалось снижение этого показателя на 2,6%.
Сопоставление данных по величине относительного превосходства гибридов над среднеродительскими показателями выявило, что по жизнеспособности гусениц, массе кокона, проценту шелковой оболочки и шелконосности сухих коконов лидирующее положение занимали гибриды, полученные от К-1 и Ш-4.
По разматываемости сухих коконов, выходу шелка-сырца сухих коконов и длине непрерывно разматываемой нити – особи от Б-1 и Ш-4; по выходу коконов с 1 г гусениц и расчетному выходу шелка – гибриды, произошедшие от К-1 и Мз-2.
Таким образом, по активности папаиноподобных ферментов при оптимумах рН 3,0, 3,6, 6,2 и 8,6, их повышению по отношению к исходным родительским формам, а также комплексу биологических и технологических показателей можно выделить группу гибридов, полученных при использовании К-1 и Ш-4, К-1 и Мз-2.
3.2 Сопряженность протеолитической активности протеиназ
с биологическими и технологическими показателями
тутового шелкопряда
С целью прогнозирования уровня хозяйственно полезных признаков по протеолитической активности были рассчитаны коэффициенты корреляции между активностью протеиназ и основными биологическими и технологическими показателями у изученных пород и гибридов тутового шелкопряда.
Корреляционный анализ выявил как положительную, так и отрицательную связь между сопоставляемыми показателями (табл. 5).
Статистически достоверная корреляция установлена между активностью папаиноподобных протеиназ с жизнеспособностью гусениц, выходом коконов с 1 г гусениц, содержанием шелковой оболочки и длиной непрерывно разматываемой нити; между активностью трипсиноподобных протеиназ и средней массой коконов и их разматываемостью, между активностью химотрипсиноподобных протеиназ и жизнеспособностью гусениц и средней массой коконов.
Таблица 5 – Коэффициенты корреляции между активностью протеолитических ферментов в грене и биолого-хозяйственными, технологическими
показателями тутового шелкопряда
Биолого-хозяйственные и технологические показатели | Фермент/ Оптимум | |||||
Папаиноподобная протеиназа | Трипсино-подобная протеиназа | Химотрипсиноподобная протеиназа | ||||
рН 3,0 | рН 3,6 | рН 6,2 | рН 8,6 | рН 9,0 | рН 7,2 | |
Жизнеспособность гусениц | 0,27 | 0,43 | 0,44 | 0,39 | -0,44 | 0,19 |
Средняя масса кокона | 0,21 | 0,07 | -0,75 | -0,34 | 0,40 | 0,20 |
Выход коконов с 1 г гусениц однодневного возраста, кг | 0,19 | 0,23 | -0,20 | 0,27 | -0,26 | -0,24 |
Содержание шелковой оболочки в сыром коконе | 0,32 | 0,06 | 0,05 | 0,39 | -0,01 | -0,21 |
Шелконосность сухих коконов | 0,67 | 0,02 | 0,20 | 0,04 | -0,34 | 0,23 |
Разматываемость оболочки сухих коконов | 0,28 | 0,11 | 0,12 | -0,22 | 0,34 | 0,11 |
Выход шелка-сырца сухих коконов | -0,07 | -0,37 | -0,02 | 0,09 | -0,02 | -0,06 |
Длина непрерывно разматываемой нити | 0,31 | 0,24 | 0,55 | 0,02 | -0,76 | -0,03 |
Расчетный выход шелка-сырца с 1 г гусениц | 0,10 | 0,26 | -0,03 | 0,01 | 0,02 | 0,12 |
Отрицательная статистически достоверная корреляция выявлена между активностью папаиноподобных протеиназ с оптимумом рН 3,0, 3,6 и 6,2 и выходом шелка-сырца; активностью папаиноподобных протеиназ с оптимумом рН 6,2 со средней массой коконов, выходом шелка-сырца сухих коконов и его расчетным выходом с 1 г гусениц; активностью папаиноподобных протеиназ с оптимумом рН 8,6 и средней массой коконов и разматываемостью сухих коконов; активностью трипсиноподобных протеиназ с оптимумом рН 9,0 и жизнеспособностью гусениц, содержанием шелковой оболочки в сыром коконе, выходом шелка, а также активностью химотрипсиноподобных протеиназ с оптимумом рН 7,2 и содержанием шелковой оболочки в сыром коконе и выходом шелка-сырца.
Следовательно, для повышения жизнеспособности гусениц, содержания шелковой оболочки, разматываемости коконов следует отбирать породы тутового шелкопряда с повышенной активностью папаиноподобных протеиназ, для увеличения средней массы кокона - трипсино - и химотрипсиноподобных протеиназ.
Следует особо подчеркнуть, что активность папаиноподобных ферментов с оптимумом рН 3,0 и 3,6 положительно коррелировала со всеми изученными показателями, за исключением выхода шелка-сырца, что следует учитывать как при отборе исходных пород, так и определении наиболее перспективных гибридных сочетаний.
Обобщение полученных результатов свидетельствует о том, что активность папаиноподобных трипсиноподобных и химотрипсиноподобных протеиназ может быть использована в качестве биохимических тестов для раннего прогнозирования потенциальной продуктивности у пород тутового шелкопряда и их гибридов.
3.3 Влияние инфекции и температурных стрессов
на продуктивность тутового шелкопряда
Исследование влияния стрессов различной природы выявило, что они вызывают значительное снижение как биологических, так и технологических показателей у тутового шелкопряда, при этом наибольшие отрицательные воздействия оказывали вирус полиэдроза и повышенная сублетальная температура.
Так, эти факторы снижали жизнеспособность гусениц на 33,9 – 75,3% (Р<0,001), в то время как переохлаждение – на 19,9 – 64,7 (Р<0,01). При этом следует отметить, что инфекционный стресс увеличивал количество больных коконов в 5 – 10 раз, тогда как температурные факторы всего в 1,5 – 2,5 раза.
Выявлена разница по силе воздействия вызванных стрессов по такому показателю как выход коконов с 1 г гусениц. Так, если при инфекционном индуцировании и сублетальной температуре его уровень снижался на 51,5 – 84,0% (Р<0,001), то при низких всего на 19,7 – 62,7% (Р<0,01; Р<0,001) (табл. 6, 7).
Все неблагоприятные факторы высокодостоверно снижали выход коконов с 1 г гусениц. Однако инфекционный стресс и перегрев у большинства пород снижали этот показатель на 72,8 – 84,0% (Р<0,001), в то время как стресс пониженной температуры – 19,7 – 34,4% (Р<0,05; Р<0,01). Аналогичная закономерность наблюдалась и по содержанию шелка в сырых коконах. Индукция ВЯП и высокая температура приводили к уменьшению этого показателя на 9,1 – 14,4%, тогда как при охлаждении – 0,7 -8,6%(Р<0,05).
Следует отметить, что при всех видах стрессовых воздействий наибольшей общей резистентностью обладали К-13 и Даидзо. Снижение уровня хозяйственно полезных показателей (жизнеспособность гусениц, средняя масса кокона и их выход с 1 г гусениц) у них по сравнению с другими породами было наименьшим.
Наименее устойчивой оказалась Б-2, как при инфекционном, так и температурном воздействиях.
Таблица 6 - Влияние различных стрессов на биологические показатели пород тутового шелкопряда
Порода | Жизнеспособность гусениц, % | Разница к контролю (-), % | Средняя масса кокона, г | Разница к контролю (-), % | Двойники, % | Разница к контролю (-), % |
Контроль | ||||||
К-13 | 95,2 ± 0,49 | - | 2,0 ±0,01 | - | 0,4 ± 0,01 | - |
ПС-5 | 94,5 ± 0,54 | - | 1,8 ±0,01 | - | 0,5 ± 0,02 | - |
Б-2 | 93,9 ± 0,62 | - | 2,0 ±0,01 | - | 0,6 ± 0,02 | - |
Сычуань | 92,4 ± 0,48 | - | 1,6 ±0,01 | - | 0,5 ± 0,02 | - |
Я-107 | 95,9 ± 0,69 | - | 1,7 ±0,01 | - | 1,3 ± 0,03 | - |
Даидзо | 94,5 ± 0,56 | - | 1,6 ±0,01 | - | 1,2 ± 0,03 | - |
Инфекционный стресс | ||||||
К-13 | 45,3 ± 0,13*** | 52,4 | 1,7 ±0,01 | 15,0 | 5,8 ±0,03*** | - |
ПС-5 | 27,0 ± 0,17*** | 71,4 | 1,1 ±0,01*** | 38,8 | 10,0 ±0,07*** | 190,0 |
Б-2 | 25,4 ± 0,21*** | 72,9 | 1,2 ±0,01 | 40,0 | 4,2 ±0,02*** | - |
Сычуань | 36,8 ± 0,38*** | 60,1 | 1,2 ±0,01 | 33,3 | 0,8 ± 0,03*** | 60,0 |
Я-107 | 29,4 ± 0,12*** | 69,3 | 1,3 ±0,01 | 30,7 | 1,9 ± 0,02 | +30,7 |
Даидзо | 62,4 ± 0,24*** | 33,9 | 1,1 ±0,01*** | 31,2 | 2,4 ± 0,03*** | 100,0 |
Температурный стресс: +40ºС | ||||||
К-13 | 37,5 ± 0,47*** | 60,6 | 1,7 ±0,01 | 15,0 | 0,5 ± 0,01** | 25,0 |
ПС-5 | 24,2 ± 0,38*** | 71,3 | 1,2 ±0,01 | 33,3 | 0,2 ± 0,01 | +60,0 |
Б-2 | 23,1 ± 0,52** | 75,3 | 1,3 ±0,01 | 35,0 | 0,4 ±0,02*** | - |
Сычуань | 28,7 ± 0,48*** | 68,9 | 1,4 ±0,01 | 12,5 | 0,3 ±0,07*** | - |
Я-107 | 25,3 ± 0,64** | 73,6 | 1,4 ±0,01 | 17,6 | 0,4 ± 0,01 | +69,2 |
Даидзо | 18,1 ± 0,71*** | 49,1 | 1,5 ±0,01 | 6,2 | 0,3 ± 0,01 | +75,0 |
Температурный стресс: +4º … +5ºС | ||||||
К-13 | 70,0 ± 0,43*** | 26,5 | 1,8 ±0,01 | 10,0 | 0,1 ±0,01 | - |
ПС-5 | 39,1 ± 0,42*** | 58,6 | 1,5 ±0,01 | 16,7 | 0,2 ± 0,01 | +60,0 |
Б-2 | 27,6 ± 0,52** | 70,6 | 1,6 ±0,01 | 20,0 | 0,4 ±0,01 | - |
Сычуань | 37,2 ± 0,39*** | 59,7 | 1,3 ±0,01 | 18,7 | 0,2 ± 0,01 | +60,0 |
Я-107 | 31,4 ± 0,46*** | 67,2 | 1,4 ±0,01 | 17,6 | 0,8 ± 0,02 | +38,4 |
Даидзо | 76,0 ± 0,56*** | 19,6 | 1,6 ±0,01 | - | 0,1 ±0,01 |
Таблица 7 - Влияние различных стрессов на технологические показатели пород тутового шелкопряда
Порода | Больные коконы, % | Разница к контролю, (количество раз) | Выход коконов с 1 г гусениц однодневно го возраста, кг | Разница к контролю (-), % | Шелковая оболочка сырых коконов, % | Разница к контролю (-), % |
Контроль | ||||||
К-13 | 2,5 ±0,02 | - | 4,22 ±0,03 | - | 20,4 ±0,51 | - |
ПС-5 | 2,4 ±0,01 | - | 3,89 ±0,03 | - | 16,7 ±0,33 | - |
Б-2 | 2,6 ±0,02 | - | 4,26 ±0,04 | - | 18,5 ±0,64 | - |
Сычуань | 1,7 ±0,01 | - | 3,51 ±0,02 | - | 14,2 ±0,46 | - |
Я-107 | 1,4 ±0,01 | - | 3,89 ±0,03 | - | 18,0 ±0,44 | - |
Даидзо | 2,1 ±0,02 | - | 3,55 ±0,02 | - | 13,1 ±0,50 | - |
Инфекционный стресс | ||||||
К-13 | 10,4 ±0,09*** | 4,2 | 1,71 ±0,02*** | 59,4 | 18,3 ±0,48 | 10,3 |
ПС-5 | 12,0 ±0,10*** | 5,0 | 0,70 ±0,03*** | 82,0 | 14,3 ±0,53*** | 14,4 |
Б-2 | 20,5 ±0,12*** | 7,8 | 0,71 ±0,03*** | 83,5 | 16,3 ±0,47 | 11,9 |
Сычуань | 18,0 ±0,09*** | 10,6 | 1,05 ±0,01*** | 70,0 | 12,4 ±0,26** | 12,7 |
Я-107 | 10,0 ±0,09*** | 7,1 | 0,91 ±0,02*** | 76,6 | 14,3 ±0,16** | 9,4 |
Даидзо | 11,5 ±0,10*** | 5,4 | 1,61 ±0,02*** | 54,6 | 11,3 ±0,14** | 13,7 |
Температурный стресс: +40ºС | ||||||
К-13 | 3,9 ±0,02*** | 1,6 | 1,41 ±0,03*** | 66,5 | 18,7 ±0,49 | 8,3 |
ПС-5 | 3,8 ±0,02*** | 1,6 | 0,67 ±0,02*** | 82,8 | 14,7 ±0,28 | 11,9 |
Б-2 | 4,5 ±0,03*** | 1,7 | 0,68 ±0,02*** | 84,0 | 16,6 ±0,26 | 10,3 |
Сычуань | 2,4 ±0,02*** | 1,4 | 0,95 ±0,03*** | 72,8 | 12,9 ±0,18 | 9,1 |
Я-107 | 2,2 ±0,02*** | 1,6 | 0,85 ±0,02*** | 78,2 | 17,2 ±0,21 | 4,4 |
Даидзо | 2,5 ±0,01*** | 1,2 | 1,72 ±0,03*** | 51,5 | 12,4 ±0,17 | 5,3 |
Температурный стресс: +4º - +5ºС | ||||||
К-13 | 10,3 ±0,09*** | 4,1 | 2,79 ±0,02*** | 33,8 | 18,8 ±0,11 | 7,8 |
ПС-5 | 4,3 ±0,08*** | 1,8 | 1,34 ±0,03*** | 34,4 | 13,2 ±0,14 | 20,9 |
Б-2 | 0,9 ±0,06 | - | 1,01 ±0,03*** | 76,3 | 16,9 ±0,19 | 8,6 |
Сычуань | 2,2 ±0,04 | 1,3 | 1,15 ±0,02*** | 67,2 | 13,4 ±0,17 | 5,6 |
Я-107 | 1,5 ±0,03 | - | 1,06 ±0,02*** | 72,7 | 17,1 ±0,10 | 5,0 |
Даидзо | 2,2±0,04 | - | 2,85 ±0,03*** | 19,7 | 13,0 ±0,14 | 0,7 |
3.4 Влияние инфекции и температурных стрессов на белковый спектр и активность рибонуклеаз гемолимфы
В ходе спектрального анализа белков у исследованных пород тутового шелкопряда выявлено, что неблагоприятные воздействия вызывали значительные изменения как в качественном распределении белковых фракций в зависимости от их электрофоретической подвижности, так и в количественном.
Так, у всех пород (за исключением Даидзо) индуцирование ВЯП вызывало появление белков с высокой молекулярной массой и соответственно низкой подвижностью (Rf = 0,01). У породы Даидзо отмечено появление белков в зоне Rf = 0,22, тогда как у пород ПС-5 - в диапазоне Rf = 0,25 – 0,27 и Б-2 в зоне Rf = 0,27 вирусная инфекция приводила к их исчезновению.
У китайских и японских пород под воздействием этого фактора отмечено значительное увеличение белков со средней молекулярной массой соответственно в диапазонах Rf = 0,35 – 0,55 на 195,6-241,0% и Rf = 0,45 – 0,55 на 150,0 – 225,0%, в то время как у российских пород в диапазоне Rf = 0,45 – 0,55 таких белков вообще не обнаружено. В зоне Rf =0,25 у всех пород (за исключением Б-2) выявлено существенное снижение (на 33,3 – 156,2%) белковых фракций, а при Rf = 0,27 их полное отсутствие.
Воздействие низких и высоких температур, так же как вирусная инфекция, у всех пород, кроме Даидзо, вызывало появление на элекрофореграммах белков с низкой подвижностью (Rf = 0,01).
Переохлаждение практически у всех пород провоцировало снижение содержания средне - и высокоподвижных растворимых белков гемолимфы в диапазоне Rf = 0,25 – 0,45, за исключением Б-2, у которой в зоне Rf = 0,25 – 0,35 отмечено значительное (на 169,5 – 471,4%) их повышение.
У пород Б-2 и Сычуань, как при воздействии низких, так и сублетальных высоких температур, происходило значительное (на 166,6-200,0% и 133,3%) повышение высокомолекулярных белков с низкой подвижностью (в зоне Rf = 0,17), тогда как у К-13 низкие температуры в указанной зоне вызывали снижение содержания белка (на 42,5%), а высокие, наоборот, повышение (на 175,0%).
Таким образом, при значительном разнообразии изменений в белковом спектре гемолимфы гусениц разных пород тутового шелкопряда под воздействием стрессов различной природы общим для всех пород было значительное возрастание количества белков со средней и особенно высокой молекулярной массой. С большей долей вероятности можно предположить, что перестройки в белковом спектре гемолимфы носят приспособительный характер и связаны с синтезом защитных белков (антител).
Другим немаловажным аспектом при изучении адаптационных возможностей тутового шелкопряда является исследование амплитуды пластичности рибонуклеазных ферментов, таких как РНК-азы и ДНК-азы.
Таблица 8 - Влияние стрессовых факторов на активность РНК - и ДНК-аз в гемолимфе различных пород тутового шелкопряда (усл. ед. на 1 мл гемолимфы, 10²)
Контроль, стрессовый фактор | К-13 | ПС-5 | Б-2 | Сычуань | Я-107 | Даидзо |
РНК-аза | ||||||
Контроль | 22,70±1,39 | 22,22±1,71 | 24,95±1,83 | 23,81±0,96 | 16,25±1,62 | 12,50±2,70 |
ВЯП | 94,16±3,38*** | 83,62±4,62*** | 99,82±3,08*** | 92,33±5,12*** | 61,66±3,91*** | 18,33±2,41 |
Разница к контролю (%) | +436,80 | +276,31 | +300,08 | +287,78 | +279,45 | +46,64 |
+4º … +5ºС | 20,80±1,15 | 19,46±2,04 | 21,19±1,97 | 20,56±2,06 | 11,70±1,52 | 12,50±1,87 |
Разница к контролю (%) | -8,37 | -12,40 | -15,08 | -13,65 | -28,00 | 0 |
+40ºС | 25,00±1,30 | 21,38±1,46 | 20,6±0,96 | 23,71±1,46 | 8,75±1,72* | 4,13±0,98* |
Разница к контролю (%) | +10,11 | -3,78 | -17,43 | -0,42 | -46,15 | -66,96 |
ДНК-аза | ||||||
Контроль | 23,04±1,13 | 20,04±0,66 | 22,86±1,02 | 21,02±1,82 | 19,28±0,99 | 22,23±1,77 |
ВЯП | 54,82±2,19*** | 41,42±3,13** | 49,54±2,47*** | 51,83±4,17*** | 41,61±3,12** | 18,16±2,10 |
Разница к контролю (%) | +137,93 | +106,69 | +116,71 | +146,57 | +115,82 | -18,31 |
+4º … +5ºС | 17,17±0,78* | 19,52±1,26 | 24,17±1,49 | 22,24±1,15 | 14,19±0,86* | 21,46±1,48 |
Разница к контролю (%) | -25,48 | -2,59 | +5,70 | +5,81 | -26,40 | -3,46 |
+40ºС | 15,16±1,12** | 14,16±0,96** | 19,86±1,07 | 21,00±2,02 | 12,16±1,87* | 16,18±2,13 |
Разница к контролю (%) | -34,20 | -29,30 | -13,12 | -0,09 | -36,93 | -27,22 |
Выявлено, что индуцирование вируса ядерного полиэдроза у всех пород вызывало значительное повышение активности РНК и ДНК-азы, при этом наибольшая активация этого фермента отмечена у К-13, Б-2, ПС-5, Сычуань и Я-107 – в 3,7- 4,1 раза (Р<0,001).
Температурные стрессы вызывали изменения, обратные тем, которые отмечались при индуцировании вируса. Переохлаждение и воздействие повышенных температур практически у всех пород приводили к снижению активности ферментов на 12,4 - 41,6% (Р<0,01; 0,001) и 3,4 - 34,2% (Р<0,05).
Сопоставление у изученных пород изменений в уровне продуктивных качеств и активности рибонуклеаз под воздействием стрессов показывает, что большую адаптационную пластичность при вирусной инфекции проявляли породы, у которых отмечалось более значительная активация РНК - и ДНК- ферментов, а при температурных воздействиях – породы, у которых снижение в активности рибонуклеаз была наименьшей. По активности рибонуклеаз и большему сохранению продуктивного потенциала при воздействии неблагоприятных факторов выделялись Б-2 и К-13.
Возможность биохимического тестирования адаптационных способностей тутового шелкопряда к стрессовым факторам по активности рибонуклеазных ферментов подтверждают полученные положительные коэффициенты корреляции между активностью этих ферментов и некоторыми продуктивными показателями тутового шелкопряда (табл. 9).
Таблица 9 - Коэффициенты корреляции между активностью рибонуклеаз гемолимфы и биологическими и технологическими показателями тутового шелкопряда
Показатели | Ферменты | |||||||
РНК-аза | ДНК-аза | |||||||
контроль | ВЯП | +4º… +5ºС | +40ºС | контроль | ВЯП | +4º …+5ºС | +40ºС | |
Жизнеспособность гусениц | -0,47 | -0,71 | -0,21 | -0,70 | -0,21 | -0,62 | -0,08 | -0,03 |
Средняя масса кокона | 0,59 | 0,37 | 0,20 | -0,02 | 0,54 | 0,56 | -0,03 | -0,10 |
Выход коконов с 1 г гусениц | 0,49 | -0,42 | -0,17 | -0,40 | 0,39 | -0,25 | -0,10 | -0,06 |
Процент шелковой оболочки сырых коконов | 0,46 | 0,67 | 0,15 | 0,31 | 0,18 | 0,70 | -0,44 | -0,39 |
При нормальных условиях выращивания гусениц активность РНК-азы имела сопряженность со средней массой кокона, выходом коконов с 1 г гусениц и процентом шелковой оболочки. При стрессовых факторах выявленная положительна связь менялась на отрицательную (за исключением коэффициентов корреляции со средней массой кокона и процентом шелковой оболочки при ВЯП и сублетальной температуре).
Активность ДНК-азы положительно коррелировала со средней массой кокона и их выходом с 1 г гусениц в контроле, однако при неблагоприятных условиях коэффициенты корреляции имели отрицательное значение (кроме связи со средней массой кокона и проценту в них шелковой оболочки при вирусной инфекции).
Обращает на себя внимание тот факт, что активность ферментов отрицательно коррелировала с жизнеспособностью гусениц при нормальных условиях, однако при стрессовых факторах (ВЯП и сублетальной температуре) величина этого показателя значительно возрастала.
Таким образом, процесс адаптации и изменение биохимических реакций при воздействии разных стрессов изменяют вектор взаимосвязи между активностью изученных ферментов и жизнеспособностью гусениц.
Полученные данные по изучению РНКазной и ДНКазной активности белков гемолимфы у пород тутового шелкопряда, их связь при различных условиях позволяют получить дополнительные параметры в оценке адаптационных возможностей при воздействии неблагоприятных факторов. Интенсивность изменения активности рибонуклеаз под воздействием стрессов следует учитывать при отборе пород и получении гибридов, оптимально сочетающих высокую устойчивость к неблагоприятным факторам с высокой продуктивностью.
3.5 Идентификация пород
тутового шелкопряда по растворимым белкам грены
Сравнение пород тутового шелкопряда по морфологическим признакам не выявило каких-либо значимых различий по цвету грены, ее форме, зернистости коконов, окраске кожных покровов гусениц, а также биологическим и технологическим показателям – средней массе коконов и их выходу с 1 г однодневных гусениц, содержанию шелковой оболочки в сырых коконах, шелконостности сухих коконов их разматываемости.
В тоже время исследование белковых спектров трех пород тутового шелкопряда позволило выявить у них достаточный широкий белковый полиморфизм. Выделено от 13 до 18 белковых фракций с различным количественным содержанием белка. Так, наиболее полиморфной оказалась НSС-90, у которой было выявлено 18 белковых фракций, затем следовала Б-2, у которой обнаружено 16 белковых фракций и менее полиморфной из изученных пород оказалась ПС-5 с 13 белковыми вариантами (рис. 2).
Сравнение исследованных пород между собой показало, что у Б-2 в зоне рН 5,3 и 5,6 присутствует белок, в то время как у ПС-5, HSC-90 в этой зоне белков нет. В зоне рН 7,5 есть белки у Б-2, HSC –90, в то время как у ПС-5 не выявлено белковых фракций. Белковый полиморфизм в зонах рН 5,7, 6,0, 6,5 и 7,1 характерен только для HSC – 90.
Таким образом, использование метода изоэлектрофокусирования белков и их электрофоретическое разделение позволяет получить белковые профили, информативность которых является достаточной для проведения идентификации пород тутового шелкопряда. Выявление маркерных белков с определенным значением рН позволит осуществлять своего рода биохимическую паспортизацию пород тутового шелкопряда и иметь объективные показатели для подтверждения чистоты живых коллекций шелководческих станций.
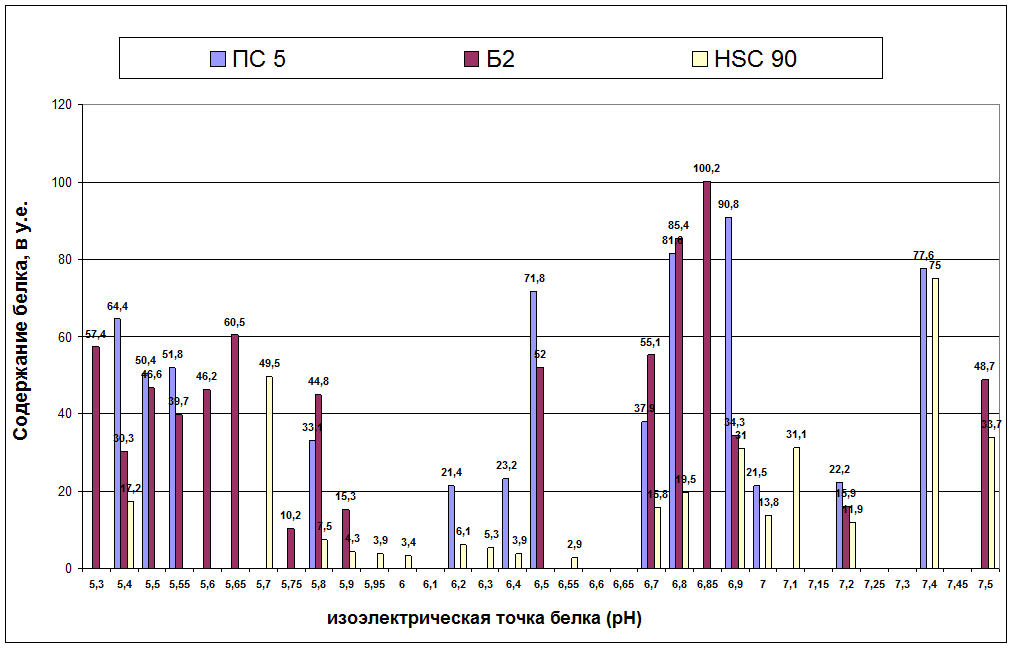
Рис. 2. Изоэлектрофокусограммы растворимых белков грены разных пород тутового шелкопряда
3.6 Экономическая эффективность
Стоимость экспериментальных выкормок для оценки хозяйственно-полезных признаков четырех пород и шести гибридов в соответствии с принятой классической схемой селекции и отбор перспективных гибридов по данным научно-производственной деятельности РНИИСШ составила 504 тыс. рублей. Стоимость проведения биохимического тестирования, позволяющего отобрать гибридные комбинации без проведения экспериментальной выкормки, составляет 34 тыс. рублей. Таким образом, предлагаемый метод биохимического прогнозирования продуктивности гибридов и отбора наиболее ценного исходного селекционного материала на ранних стадиях эмбриогенеза позволяет сэкономить 470 тыс. рублей.
Экономия затрат при использовании метода прогнозирования адаптационных качеств тутового шелкопряда по биохимическим показателям за счет сокращения объема выкормок в 8,2 раза составила 442,5 тыс. рублей.
ВЫВОДЫ
1. В составе растворимых белков грены тутового шелкопряда пород Б-1, Ш-4, Мз-2 и их прямых и обратных гибридов выявлен сложный комплекс протеолитических ферментов, представленный папаино-, трипсино - и химотрипсиноподобными протеиназами с различной активностью в диапазоне рН от 2,2 до 11,0.
2. Большую продуктивность имели гибриды К-1хШ-4 и К-1хМз-2: их превосходство над гибридными сочетаниями Ш-4хК-1, Мз-2хК1, Б-1хШ-4 и Ш-4хБ-1 по жизнеспособности гусениц, выходу коконов, их шелконосности и количеству шелка-сырца составило соответственно 2,4 абс. проц., 4,1%, 2,9 абс. проц. и 6,1%. Кроме того, они имели большую разницу по сравнению с родителями по активности протеолитических ферментов, выходу коконов и шелка-сырца, чем другие гибриды соответственно на 16,3%, 2,1% и 3,5 абс. проц.
3. Установлена сопряженность активности папаиноподобных протеиназ с жизнеспособностью гусениц, выходом коконов с 1 г гусениц, содержанием шелковой оболочки и длиной непрерывно разматываемой нити (r=0,19 …0,67); трипсиноподобных – со средней массой коконов и их разматываемостью (r=0,403; r=0,346), химотрипсиноподобных – с жизнеспособностью гусениц и средней массой коконов (r=0,19), что позволяет рекомендовать активность данных ферментов в качестве биохимических тестов для раннего прогнозирования продуктивности у гибридов тутового шелкопряда.
Наиболее информативными являются папаиноподобные протеиназы с оптимумом рН 3,0, 3,6 и 8,6. Гибриды, у которых их активность выше, чем у исходных родителей, являются перспективными для отбора и последующего использования в селекционном процессе.
4. Воздействие стрессов различной природы снижает как биологические, так и технологические показатели у тутового шелкопряда, при этом наибольшее отрицательное влияние оказывают вирус полиэдроза и сублетальная повышенная температура, которые уменьшают жизнеспособность гусениц, среднюю массу кокона, выход коконов и процент шелковой оболочки в среднем на 63,2; 25,7; 71,8 и 10,1% соответственно, тогда как пониженная температура на 48,0; 13,8; 49,1 и 7,1%.
5. Неблагоприятные факторы вызывают значительные изменения, как в качественном, так и в количественном распределении белковых фракций в гемолимфе гусениц тутового шелкопряда.
5.1. Вирусная инфекция у всех изученных пород (за исключением Даидзо) приводит к появлению белков с высокой молекулярной массой в зоне Rf=0,01; у китайских и японских пород к повышению на 150,0…241,0% белков со средней молекулярной массой в диапазоне Rf=0,35-0,55, у всех пород (за исключением Б-2) снижению белков на 33,3 – 156,2% в зоне Rf=0,25 и полному их отсутствию в зоне Rf=0,27.
5.2. Температурные стрессы снижают содержание белков со средней молекулярной массой в диапазоне Rf=0,25-0,45, а также приводят к исчезновению белковых фракций в зоне Rf=0,04 у Даидзо, Rf=0,35 у К-13, Rf=0,45-0,55 у Б-2 и Rf=0,48 у Я-107.
6. Стрессовые факторы изменяют активность рибонуклеаз: вирусная инфекция приводит к повышению активности РНК - и ДНК-аз соответственно в 3,7-4,1 (Р<0,001) и 2,1-2,5 раз (Р<0,01), воздействие температур, наоборот, к их понижению на 12,4 - 41,6% (Р<0,01; 0,001) и 3,4 - 34,2% (Р<0,05).
7. Большую адаптационную пластичность при вирусной инфекции проявляют породы, у которых отмечается более значительная активация РНК - и ДНК-ферментов, а при температурных воздействиях – породы, у которых снижение в активности рибонуклеаз была наименьшей. По активности рибонуклеаз и большему сохранению продуктивного потенциала при воздействии неблагоприятных факторов выделялись Б-2 и К-13.
8. Выявлены положительные коэффициенты корреляции между активностью РНК-, ДНК-аз и средней массой коконов (r=0,37-0,59), их выходом с 1 г гусениц (r=0,43-0,63), РНК-азы и процентом шелковой оболочки (r=0,39-0,49), что позволяет использовать активность рибонуклеаз в качестве дополнительных критериев при отборе стресс-устойчивых пород тутового шелкопряда.
9. У морфологически неразличимых пород тутового шелкопряда – ПС-5, Б-2 и HSC–90 выявлены специфичные белковые профили: у Б-2 в зоне рН 5,3 и 5,6 имелись белковые фракции, в то время как у ПС-5, HSC-90 их не обнаружено; у Б-2, HSC-90 в зоне рН 7,5 присутствовали белки, в то время как у ПС-5 нет; белковый полиморфизм в зонах рН 5,7, 6,0, 6,5 и 7,1 характерен только для HSC – 90.
ПРАКТИЧЕСКИЕ ПРЕДЛОЖЕНИЯ
1. Для ускорения селекционного процесса и сокращения материальных затрат при использовать метод биохимического прогнозирования продуктивности гибридов и отбора наиболее ценного исходного селекционного материала на ранних стадиях эмбриогенеза.
2. С целью прогнозирования адаптационных способностей пород тутового шелкопряда использовать метод, основанный на сопоставлении белковых спектров гемолимфы и активности рибонуклеаз.
3. При оградительной и поддерживающей селекции коллекционных пород тутового шелкопряда для идентификации морфологически трудно различимых пород использовать спектры растворимых белков грены.
СПИСОК ОПУБЛИКОВАННЫХ РАБОТ
Публикации в изданиях, рекомендованных ВАК Минобразования и науки РФ
1. Богословский тесты прогнозирования продуктивности и гетерозиса у тутового шелкопряда // Вестник РАСХН, 2009. №2. С. 84-85.
2. Богословский пород тутового шелкопряда по растворимым белкам грены // Известия ТСХА, Вып.С. 151-153.
Публикации в других изданиях
3. , , Филиппович локализация ферментов и их белковых ингибиторов в яйцах тутового шелкопряда // Сб. научн. тр. МГПУ. Серия: естеств. науки. М.: Прометей, 2000. – С. 249-252 (0,2 п. л., доля авторского участия 20%).
4. , , Филиппович цистеиновых протеиназ и белковых ингибиторов папаина в грене различных по продуктивности пород тутового шелкопряда // Сб. научн. тр. МГПУ. Серия: естеств. науки. М.: Прометей, 2001. – С. 183 – 186 (0,25 п. л., доля авторского участия 20%).
5. , , Клунова сериновых протеиназ в грене родительских пород и гибридов тутового шелкопряда // Сб. научн. тр. МГПУ. Серия: естеств. науки. М.: Прометей, 2005. – С. 103 – 106 (0,25 п. л., доля авторского участия 20%).
6. Метод биохимической диагностики гетерозисной силы исходного селекционного материала на ранних стадиях эмбриогенеза тутового шелкопряда / / РНИИСШ. Железноводск, 20с.
7. Метод тестирования адаптационных способностей пород тутового шелкопряда на основе сопоставления спектров растворимых белков тканей организма / , , / РНИИСШ. Железноводск, 20с. (0,72 п. л., доля авторского участия 70%).
8. Метод идентификации пород тутового шелкопряда по наборам растворимых белков грены» / , , / РНИИСШ. Железноводск, 20с. (0,50 п. л., доля авторского участия 60%).
9. Богословский генетические признаки тутового шелкопряда // Матер. междунар. научно-практич. конф. Вып. 61. Т. II, Оренбург, ВНИИМС. 2008. С. 18-22.
