Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ЛЕКЦИЯ 20. МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ (незавершенный черновик!)
, ИЦиГ СО РАН и ФЕН НГУ, Новосибирск, 2012 г.
20.1. Классификация и свойства мобильных генетических элементов
20.1.0. Общие замечания.
Способны либо к транспозициям, либо к дупликациям путем инсерций. Кодируют необходимые для этого ферменты, но нуждаются и в ферментной машине «хозяина». Большинство копий дефектны - неавтономны, и либо перемещаются или размножаются с использованием ферментов, кодируемых недефектными – автономными - копиями, либо вообще не способны к перемещению и деградируют за счет мутаций.
Два больших класса и один (или больше) маленьких.
1. Ретротранспозоны.
Через РНК-посредники, ревертазу (русское название, по английски – обратная транскриптаза). Сначала строит цепь ДНК по матрице РНК, потом РНКаза гидролизует РНК и та же самая ревертаза строит вторую цепь ДНК, гидролизуя цепь РНК. В геноме ДНК ретротранспозона всегда остается на месте. За счет этого: а) общая их масса в геноме имеет тенденцию к увеличению и б) индуцированные их инсерциями мутации стабильны.
1.1. LTR-ретротранспозоны
(Long Treminal Repeat) – с длинными (100 – 5000 пар оснований) прямыми терминальными повторами, сходны с ретровирусами, но не образуют вирионов, чем только и отличаются. Встроенные в геном ретротранспозоны ничем не отличаются от провирусов. Обычно имеется два гена. Первый сходен с геном gag ретровинусов, кодирующий белковые компоненты нуклеопротеинового компонента вирионов, второй – с геном pol, ответственным за все активности, связанные с транспозицией: протеазную, ревертазную, РНК-азную и интегразную. Они транслируются как полипротеид, но затем разрезаются протеазами на отдельные ферменты (иногда не до конца). Иногда идет трансляция gag и pol как единого полипротеида. Иногда есть третий ген, позиция которого совпадает с позицией гена ретровирусов env, кодирующего белки капсида, однако гомологии с этим геном не прослеживается. Потеря функционального гена env и отличает LTR ретротранспозоны от ретровирусов.
Транскрибируется как единая мРНК. LTR имеют три участка – U3, R и U5, транскрипция от 5’конца левого R до 3’конца правого R. – обратная транскрипция через кольцевую форму. Обратная транскрипция происходит в цитоплазме, поскольку первоначальной затравкой служит тРНК, к которой «правее» левого LTR имеется комплементарный участок. От нее строится комплементарная цепь ДНК до 5’-конца РНК. Затем РНК в ДНК-РНК дуплексе удаляется, но в месте посадки тРНК находится РНК-РНК дуплекс, который сохраняется. РНК ретротранспозона закольцовывается и одноцепочечная ДНК праймирует построение второй цепи ДНК на оставшемся участке. Это происходит в цитоплазме, но затем двунитевая ДНК ретротранспозона перемещается в ядро и встраивается в геном.
У эукариот три основных класса: Ty1- copia like, Ty-3 gypsy like, Pao BEL (эти только у животных).
1.2. Non-LTR Retrotransposones
1.2.1. LINE (Long Interspersed Elements) – не имеют LTR. На 3’конце – сигнал полиаденилирования и поли-А (в кодирующей цепи). То есть с виду не имеют структур, специализированных для встройки с полным восстановлением собственной структуры, а скорее выглядят как «обычные» мРНК. Однако для обратной транскрипции и встройки с возможностью дальнейшей транскрипции, имеют специальный – внутенний промотор для РНК-полимеразой II (обычно промоторы для нее внешние). Ею и транскрибируются. Терминация транскрипции в районе поли-А, но они и достраиваются. Обратная транскрипция идет не в цитоплазме, а прямо в ядре. Затравкой служит 3’-конец при внесении однонитевого разрыва в ДНК. , промотор к которой у них (напомню, что РНК-полимераза II). В 3’-конце имеют мишень для ревертазы – участок, ею опознаваемый. Имеют ген, подобный gag и ген, ответственный за все функции, необходимые для встраивания – связь с нуклеиновыми кислотами, ревертазную (особое сродство именно к собственной РНК), нуклеазную.
1.2.2. SINE (Short Interspersed Elements) –до 1500 пар, но обычно меньше 500 пар. Если Изначально тРНК, рРНК и прочие малые РНК, транскрибируемые РНК-полимеразой III. Если у них перед поли-А имеется участок, похожий на 3’-нетранскрибируемый район LINE, то ревертаза от LINE ошибочно распознает их как свои и они встраиваются. Чтобы после встройки они оставались способными к транскрипции и дальнейшему размножению, очень желательно, чтобы у них сохранялся промотор, то есть чтобы такая РНК транскрибировалась со внутреннего промотора (то есть чтобы РНК-полимераза транскрибировала тот промотор, с которым связалась). Таким свойством обладают промоторы к РНК-полимеразе III, поэтому SINE обычно происходят от тРНК или 7SL РНК (как у Alu-повтора.), синтезируемой РНК-полимеразой III, и ею же транскрибируются. Размножаются за счет ревертазы и интегразы от LINE.
У позвоночных много, у дрозофилы нет.
Две копии SINE, оказывающиеся рядом друг с другом, при дефекте терминатора транскрипции у левого, превращают любую фланкируемую ими последовательность в составной ретротранспозон. Так много чего.
2. ДНК-транспозоны. Обычно Cut and paste. Без РНК-посредников. Транспозазы, той или иной сайт-специфичности. Кодируют как минимум одну функцию – транспозазную. Можно было бы ожидать, что обратные повторы могли бы образовывать двойную шпильку за счет образования дуплексов каждой из цепей ДНК, но для этого они длинноваты. На самом деле двойная цепь ДНК образуют одинарную шпильку, но не за счет комплементации ДНК, а за счет одинаковой картины белков, опознающих инвертированные повторы. Выстригаются с некомплементарными друг другу (!) висящими концами. Встраиваются по инвертированным коротким повторам, например ATTA, которые надрезают также с висящими 5’-концами, которые компементарны, но сшиваются с висящими концами транспозона не как комплементарные липкие концы (которыми часто называют), а именно как некомплементарные однонитевые ДНК конец-к-концу и двойная брешь потом залатывается. При выстригании остаются двунитевые разрывы с некомплиментарными висячими 5’-концами, которые репарируются либо по гомологу, либо как негомологичный end-joining, во втором случае остается «транспозоновый след» (transposone footprint), в некоторых случаях это делает специальный фермент, который оставляет след в виде тринуклеотида, который не нарушает рамки считывания. Поскольку при встраивании ДНК «хозяина» разрезается с висячими концами, а при выстригании нечто встраивается между ними, а недостающая цепь залечивается, транспозоновый след оказывается обрамленным короткими – 4-8 нуклеотидных пар – дупликациями хозяйской ДНК.
Обычно транспозоны прыгают во время репликации, тогда дыра может зарепарироваться по гомологу, где транспозон не выпрыгнул, тогда у нас получится сохранение копии в геноме плюс одна свободная и может произойти увеличение копийности.
Некоторые все-таки не вырезаются, а дуплицируются, например Helitron у эукариот – по механизму катящегося кольца.
У бактерий есть класс, элементарные представители которого - «инсерционные последовательности», IS - имеют только фермент для перемещения и один регулирующий ген, а также инвертированные повторы по краям; всего пар оснований. Впрочем, mariner-like у эукариот им родственен, и для него как раз характерны случаи горизонтального переноса между неродственными таксонами. На основе IS возникают комплексные транспозоны – они состоят из двух IS по краям и участка с добавочными структурными генами между ними – например, устойчивость к антибиотикам – и перепрыгивать из генома в плазмиды и обратно, делая их чувствительными. Впрочем, у бактерий есть и транспозоны со структурными генами, не основанные на тандемах IS.
3. Элементы с длинными инвертированными концевыми повторами. Небольшой класс, механизм перемещения неизвестен. fold back у дрозофилы.
Все бывают автономными (кодирующими функциональную ревертазу или транспозазу) или неавтономными – перемещающимися за чужой счет.
20.2. Наиболее известные примеры мобильных генетических у эукариот и их открытие.
20.2.1. Открытие транспозонов у кукурузы
Очень легко и удобно говорить, когда есть сиквенсы, а вот найти феноменологию подобного типа классическими методами было очень трудно. Но без них могли бы не обратить внимание.
Барбара Мак-Клинток

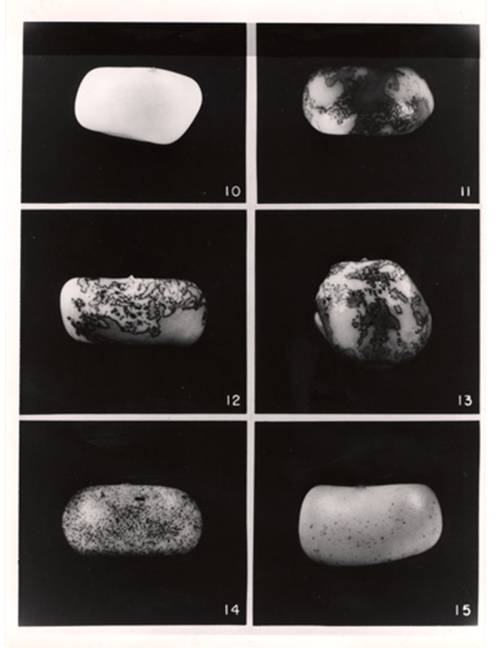
, кукуруза как объект, первая генетическая карта ее, в 1944 г. в национальную академию наук (в возрасте 42 г.), третья женщина за историю, в том же году – президент генетического общества Америки. Изучала разрывы хромосом под действием радиации. Идентифицировала образующиеся кольцевые хромосомы, которые спонтанно теряются в соматических клетках, делая листву пятнистой. На этом основании она впервые постулировала существование теломер. Изучала разрывы хромосом в эндосперме. Оборванные хроматиды, без теломеры, зашивались концами при репликации ДНК, образовывали мосты в анафазе, рвались и снова зашивались, все это сопровождалось мутагенезом. Breakage-fusion-bridge cycle. Иногда то же происходило и спонтанно. В том же 1944 году начала исследование мозаичной окраски алейрона. Нашла два гена – Ds и Aс (диссоциатор-активатор). Первый вызывал разрывы хромосом, инсерции делеции, мутации, но только в присутствии второго, оба доминантны, по сути комплементарны. Зависело от дозы Ac. В 1948 г. обнаружила, что Ds может менять свое положение, что могло сопровождается разрывом хромосомы и приводить к хромосомным перестройкам, но только в присутствии Ac. Ас также оказался подвижным. Она назвала их контролирующими элементами. Присутствие контролирующего элемента вызывала нестабильную мутацию, которая могла ревертировать к дикому типу, а также повышало вероятность возникновения мутации поблизости. Исчезновение контролирующего элемента делало район генома стабильным. В частности, она следила за ненаследуемой пестротой алейрона, она возникала в присутствии Ac, то есть в некоторых клеточных клонах восстанавливалась нормальная окраска (отсюда мозаицизм) – это значило, что Ds уходил из гена антоциановой окраски алейрона. Частота зависила от дозы Ac. На нижеследующем рисунке вверху слева показано зерно без пятен с генотипом ac ac ac), справа вверху и в середине – Ac ac ac (одна доза активатора, появляются крупные окрашенные сектора), внизу слева – Ac Ac ac (две дозы активатора, сектора мелкие), внизу справа – Ac Ac Ac (три дозы активатора, сектора мелкие, окраски мало).
В 1950 г. статья в ПНАСе «Происхождение и поведение мутабильных локусов у кукурузы». Полагала, что это – элементы регуляции генов в онтогенезе, благодаря чему генетически идентичные клетки и оказываются разными. Нашла еще одну систему – Sp m – супрессор-мутатор, с более сложным поведением. В 1953 г. опубликовала свои первичные данные. Скептицизм. После этого на эту тему не печаталась. В южноамериканскую этноботанику кукурузы. Но в 1961 году Жакоб и Моно открыли лактозный оперон и она опубликовала в American Naturalist статью, где проводила параллели.
Транспозоны были открыты у бактерий и дрожжей в конце 60х и 70х молекулярно-генетческими методами. В 1970 г. Ac и Dc были клонированы и оказались транспозонами второго класса. Ac – полный и содержит неповрежденный ген транспозазы.
Нобелевская 1983 г. (в возрасте 81), единственная женщина и единственная неразделенная по биологии. Замуж не выходила, детей не имела.
У кукурузы три системы – Ac-Ds, Spm (супрессор-мутатор) и Dt. Ас – 4563 нуклеотидных пар, инвертированные повторы на концах, один ген, кодирующий транспозазу, с 5 экзонами. Ds – это неполные, с внутренними делециями, копии.
20.2.2. Гибридный дисгенез у дрозофилы.
У F1 от скрещивание самцов, взятых из природы, с давними лабораторными линиями. Температурозависимая стерильность (прямая связь), включающая сильнейшее недоразвитие гонад, высокая мутабильность в клетках зародышевого пути, повышенный уровень рекомбинации, рекомбинация у самцов. Назвали P-M (от paternal и maternal). Есть и Q-цитотип – не индуцирует и не индуцируется. В реципрокных скрещиваниях ничего не происходит, потому что у самок Р-цитотипа в цитоплазме накоплен репрессор.
Существует аналогичная система I-R (индуктор-репрессор), независимая от системы P-M, но имеющая точно такую же феноменологию, за тем исключением, что в дисгенном направлении не наблюдается редукции яичников.
Анализ мутаций, возникающий при гибридном дисгенезе выявил встройки мобильных элементов 2го типа, то есть ДНК-транспозонов. Тот, что P-M –систему был назван P-элемент – самый знаменитый и широко используемый. Полноценный 2907 н. п., терминальные повторы 31 н. п. Транскрипт 2,7 килобаз. Способен перемещаться только в клетках зародышевого пути. От скрещивания самцов c автономным Р-элементом (Р-цитотип) с самками, лишенными автономного Р-элемента (М-цитотипа).
Надо сказать, что среди возникших мутаций найдены и встройки других мобильных элементов, например copia, которая вообще ретротранспозон. Либо перемещение Р-элемента провоцирует перестройки, либо при скрещивании происходит активация и других мобильных элементов, не связанная с Р-М системой, но остающаяся незамеченной, так как не дает такого многостороннего синдрома.
Гибридный дисгенез способна вызывать любая хромосома, пришедшая в линию с М-цитотипом от линии с Р-цитотипом (а это, как вы помните, можно устроить с использованием балансеров), поскольку его вызывают именно P-элементы, которых автономных в геноме Р-цитотипа штук 30-50.
20.3. Защита от мобильных элементов
20.3.1. Вводные замечания
Сама история открытия мобильных элементов говорит о них прежде всего как об эндогенных мутагенных факторах. Мутации бывают вредные, полезные и нейтральные. Хорошо известно и интуитивно понятно в силу принципа «ломать – не строить», что вредных мутаций бывает гораздо больше, чем полезных (тогда как относительно доли и даже самого существования нейтральных мутаций еще 2-3 десятка лет назад велись ожесточенные баталии). Это означает, что перемещение мобильных элементов гораздо чаще снижает, чем повышает приспособленность носителей – это высказывание эквивалентно утверждению о том, что против перемещения мобильных элементов существует естественный отбор. Под действием этого отбора были выработаны механизмы защиты от подвижности мобильных элементов.
Но систем защиты от самого присутствия транспозонов в геноме большинство эукариот не выработало, благодаря чему их общее количество проявляет впечатляющую тенденцию к накоплению. Исключением здесь служат только сумчатые грибы, которые научились бороться с повторами, и инфузории, которые научились очищать от ненужной ДНК хотя бы свой соматический геном – геном макронуклеуса. Оба эти случая будут рассмотрены ниже, в разделах 20.3.4 и 20.6.5. Большинство же эукариот, подобно примитивной медицине, борются с симптомами болезни – встройками транспозонов, а не с ее нозологической причиной. С их присутствием в геноме, как и с присутствием любой «мусорной ДНК», большинство организмов предпочитают мириться по всей видимости в связи с гораздо более низкой «ценой» репликации лишней ДНК по сравнению с ценой поддержания специальных и с необходимостью сложных механизмов, которые были бы способна отличать нужную ДНК от ненужной в ситуации многоклеточности, когда многие гены в большинстве клеток не работают. Такую систему могли себе позволить лишь, например, просто устроенные сумчатые грибы и инфузории. Мобильные элементы мы находим у всех организмов, а элементы системы защиты от мобильных элементов и вирусов –у всех эукариот (кроме немногих одноклеточных, которые их потеряли) и археобактерий. Мутации по белкам этой защитной системы могут приводит к столь сильной активизации транспозонов, что индуцируемые их перемещениями разрывы ДНК оказываются многочисленными настолько, что клетки могут не пройти один из чекпойнтов клеточного цикла и подвергается апоптозу. Феномены такого рода мы и наблюдаем в случае гибридного дисгенеза: в частности, недоразвитые яичники при дисгенезе в Р-М системе. Однако не все такие мутации и не у всех организмов имеют серьезные последствия, поскольку системы защиты от транспозонов как правило многократно дублированы.
По всей видимости возникновение нового транспозона или скорее проникновение его из другого вида носит характер эпидемии, сопровождающейся вспышкой мутаций. Это довольно очевидно для LTR - ретротранспозонов, которые суть редуцированные ретровирусы, но по всей видимости справедливо для всех классов транспозонов. Однако очень скоро экспрессия их подавляется и они перестают активно размножаться (ретротранспозоны) или транспонироваться (ДНК-транспозоны).
Так, еще недавно серьезно рассматривалась гипотеза о том, что Р-элемент попал в D. melanogaster от другого вида дрозофилы примерно 50 лет назад, первоначально в природные популяции Америки, и стал распространяться, так что к настоящему времени он должен был попасть во все популяции вида. Но эта гипотеза не подтвердилась: Р элемент преимущественно сохранился в тех же популяциях, в которых и был; ситуация в целом не изменилась. А наличие целых двух специфических для Р элемента систем подавления его перемещений – в клетках соматического и зародышевого пути – которые мы рассмотрим ниже, полностью исключает такую возможность. Вторая вообще основана на существовании в геноме «древних», как они называются в публикациях, копий мобильных элементов. Конечно, нельзя исключить исчезновение активных копий и их вторичное проникновение из другого вида.
В 70-80е годы сообщалось о «моде на мутации» в популяциях дрозофилы на Украине (), причем часть этих мутаций была нестабильной. Как выяснилось, эта мода, как минимум частично, связана не с распространением мобильных элементов, а с распространением определенных перестроек, делавших достаточно вероятной рекомбинации между двумя негомологичными копиями транспозонов, уже имеющихся в геноме, причем это были не P-элементы, как ожидалось, а hobo-транспозоны.
20.3.2. Блокировка сплайсинга мРНК транспозазы в соматических клетках дрозофилы
В только что рассмотренном примере гибридного дисгенеза у дрозофилы примечательно то, что он не происходит в соматических клетках. В них присутствует соматический репрессор экпрессии Р-элемента на уровне транскрипции (белок PSI, P-element somatic inhibitor), который препятствует вырезанию из мРНК транспозазы второго интрона, что приводит к появлению обрезанного белка, который ингибирует транспозицию.
20.3.3. РНК-интерференция
В клетках зародышевого пути дрозофилы все по иному. Феномен существования Р-цитотипа, когда функциональный Р-элемент не индуцирует гибридного дисгенеза, основан на сложном и совершенно замечательном явлении, которое является древним и основным механизмом борьбы с мобильными элементами - РНК-интерференцией.
Сразу скажу, что ничего общего с интерференцией кроссоверных событий обсуждаемое явление, как и она свойственное за немногими исключениями всем эукариотам, не имеет; этот латинский термин всего лишь означает, что что-то чему-то мешает (эка невидаль).
РНК-интерференция зиждется на двух семействах белков, называемых Dicer (игрок в кости) и Argonaut и запускается двуцепочечными РНК, которые могут быть как эндогенного, так и экзогенного происхождения. Когда такая РНК появляется в клетке, она режется белками из семейства Dicer на короткие – 20-25 пар – фрагменты с двунуклеотидным висячим 3’-концами. Эти короткие двуцепочечные РНК связываются с одним из белков семейства Argonaut. Этот белок является главным компонентом одного из двух типов белковых комплексов, условно разделяемых по месту и результату своего действия, RISC и RITS. Первый из них локализуется в цитоплазме, второй – в ядре.
Связавший с короткой двуцепочечной РНК, белок семейства Argonaut из RISC-комплекса гидролизуют одну из ее цепей, а второй цепью (leading-chain) сканирует на гомологию с ней клеточный пул мРНК. Когда гомология найдена, RISC-комплекс связывается с такой мРНК и либо просто блокирует трансляцию или чаще осуществляет ее гидролиз за счет РНК-зной активностью все того же Argonaut. Эта активность концентрируется в цитоплазме в районе так называемых P-телец, вокруг которых наблюдается высокая скорость деградации РНК. В данном случае следствием РНК-интерференции является так называемый посттранскрипционный сайленсинг.
В ядре аналогичным образом leading chain короткой РНК в составе RITS-комплекса опознает мРНК в момент их транскрипции и запускает процесс гетерохроматинизации того участка ДНК, с которой эта мРНК считывалась Белок семейства Argonaut, входящий в RITS-complex имеет и такую активность, как модификация гистонов, в частности их метилирование. Гетерохроматиновое состояние исключает транскрипцию и довольно нестрого называется РНК-зависимым транскрипционным сайленсингом. Это явление изучено в основном у пивных дрожжей (у пекарских РНК-интерференция отсутствует) и инфузорий, но имеет весьма имеет широкое распространение у эукариот.
Ученые приходят к согласию во мнении, что первоначальная функция РНК-интерференции уже у общего предка всех эукариот имела своего рода иммунный характер и состояла в сайленсинге сторонних и прочих нежелательных последовательностей, прежде всего вирусов и мобильных элементов. Можно спросить – причем тут двуцепочечная РНК? Интерпретировать это можно так, что встраивание в геном вируса или мобильного элемента более или менее случайным образом случайно же приводит к тому, что в каких-то случаях он попадет в состав транскрипта с какого-нибудь хозяйского промотора, причем с равной вероятностью с матричной и кодирующей цепи. В последнем случае в геноме появится антисенс-РНК, комплементарная РНК собственного законного транскрипта вируса или мобильного элемента, они образуют дуплекс и запустят РНК-интереференцию. Любая двуцепочечная РНК, экзогенная или эндогенная, режется на те самые короткие фрагменты, называемые siRNA (20-25 нуклеотидов), которые запускают сайленсинг гомологичной им мРНК или самого гена, с которого она транскрибировалась.
В настоящее время эта защитная функция особенно характерна для растений, где РНК-интерференция по сути выполняет функцию иммунной системы. В частности, у них имеются множественные варианты белков Dicer, связанные с устойчивостью к разным болезнетворным агентам.
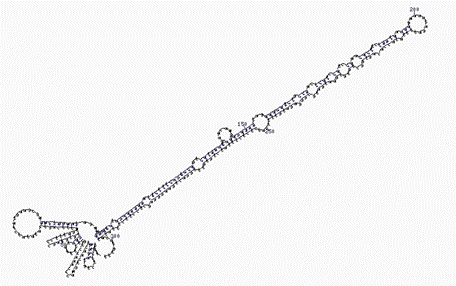
Эволюционно более молодой функцией РНК-интерференции является регуляция трансляции. По геному рассеяны специальные гены малых РНК - miRNA, транскрипция которых дает РНК-предшественники в несколько сот пар оснований, имеющие характерную вторичную структуру с петлями и двуцепочечными участками с несовершенной гомологией. На рисунке показан такой предшественник длиной чуть более 300 нуклеотидных пар из капусты:
Они претерпевают процессинг все теми же белками семейства Dicer и двуцепочечные участки высвобождаются, давая miRNA (21-23 нуклеотида) и запуская посттранскрипционную РНК-интерференцию. Они имеют гомологию с участками мРНК тех генов, трансляцию которых они регулируют. Экспрессия генов малых РНК блокирует трансляцию многих генов сразу, это есть способ быстро и одновременно выключить их трансляцию. В частности, у растений посредством такого посттрансляционного сайленсинга регулируются транскрипционные факторы, у животных – многие факторы регуляции клеточного цикла. Иногда miRNA или siRNA, гомологичные участком промотора, используются не для репрессии, а для активации транскрипции.
Небольшого количества двуцепочечной РНК бывает достаточно, чтобы блокировать экспрессию гомологичного гена на уровне трансляции или трансрипции. Как минимум у Coenorhabdites elegans формируется петля положительной обратной связи, поскольку к первичным присоединяются вторичные, специально строимые РНК-зависимой РНК-полимеразой. Активность этого фермента тесно связана с активностью RISC-комплекса. Однако этот же механизм предполагается и для эпигенетического поддержания гетерохроматинизированного состояния при РНК-зависимом транскрипционном сайленсинге. РНК-зависимые РНК-полимеразы присутствуют у всех эукариот и тех археобактерий, у которых есть и другие элементы РНК-интерференции – семейство белков Dicer и Argonaut (однако они может иметь и вирусное происхождение - от вирусов, носителем наследственности которых является двуцепочечная РНК). Обратим внимание, что мы здесь сталкиваемся с пограничным случаем между эпигенетическим и генетическим наследованием. Наследование молчащего состояния здесь осуществляется при помощи матричного биосинтеза полинуклеиновой кислоты – РНК - и в принципе это мало отличается от наследования РНК-содержащего вируса, у которого носителем наследственности выступает РНК, а в воспроизводство вовлечены ферментные системы хозяина, не кодируемые этой РНК. У растений siRNA могут распространяться от клетки к клетке через плазмодесмы вплоть до того, что ими могут обмениваться привой и подвой. Тем самым, иммунный статус приобретает все растение.
РНК-интерференцию открыли случайно в 1986-начале 1990х гг. у петунии в благородных попытках сделать ее цветки более насыщенно окрашенными путем трансгенеза. Ввели дополнительную трансгенную копию гена халькон синтетазы, вовлеченной в синтез антоциана. При трансгенезе трансгены часто замолкают. Однако тут замолчал и свой законный ген и цветы получились с белыми пятнами.

Выяснилось, что феномен связан с ускоренной деградацией мРНК, его назвали косупрессией генной экспрессии. (По-видимому, трансген встроился вблизи промотора, который обеспечил незаконную транскрипцию с его кодирующей цепи, в дополнение к правильной транскрипции с матричной цепи. Получавшиеся комплементарные РНК образовывали дуплекс.) Затем обнаружили, что небольшие кусочки вирусной ДНК, встроенной в геном растения, вызывает устойчивость к вирусу. Тогда встроили небольшие кусочки растительных генов в вирус и обнаружили, что при инфекции эти гены замолкают. Назвали вирус-индуцированным генным сайленсингом. Роль двуцепочечных РНК выяснили на ценорабдитесе, за что и дали нобелевскую премию 2006 г.
Несложно догадаться, что РНК-интерференция является мощным средством в руках исследователя, врача и биотехнолога, позволяющим выключать гены по своему усмотрению. Это могут быть гены, интересующие генетика, гены болезнетворных агентов, онкогены или гены, экспрессия которых нежелательна для производства. Чтобы это сделать, достаточно каким-либо образом доставить в клетку достаточное количество антисенс-РНК нужного гена, чтобы она образовала дуплекс с сенс-РНК, либо лучше сразу гомологичную ему двуцепочечную РНК. (Антисенс-РНК подавляет транскрипцию не путем прямого образования неспособного к транскрипции РНК дуплекса, как можно было бы наивно подумать). Проще всего организовать доставку у C. elegans – если кормить его кишечной палочкой, продуцирующей двуцепочечную РНК, то она неизвестным образом попадает во все клетки. Доставка у других организмов обычно осуществляется при помощи какого-то переносчика – вектора. Чаще всего используются векторы на основе вируса или транспозона, в ДНК которых методами генной инженерии вставлены соответствующие гены. Вирусный вектор должен инфицировать все или большинство целевых клеток, экспрессировать двуцепочечные или антисенс-РНК и не вредить хозяину. У дрозофилы удобно применять вектор, основанный на Р-элементе. Существуют успешные попытки бороться таким образом со многими вирусными заболеваниями, например СПИДом, перспективна борьба этим методом с раком.
Вернемся к гибридному дисгенезу дрозофилы. Оказалось, что у дрозофилы природа супрессора транспозиций ДНК-транспозонов в зародышевом пути сводится к той самой имунной системе защиты от транспозонов, основанной именно на PiWi-белках (от P-Element Induced Wimpy Testis) из семейства Argonaut и так называемых piRNA, которые длиннее обычных miRNA и siRNA: 26-32 пары нуклеотидов. И те, и другие производятся материнским организмом, накапливаются в заднем полюсе яйца и оказываются в цитоплазме будущих клеток зародышевого пути. При этом высокая экспрессия тех и других поддерживается эпигенетически за счет piRNA. Если их изначально не было в цитоплазме, как у продуктов дисгенного скрещивания, система не запускается. piRNA, связывающаяся с белком PiWi – антисмысловая к мРНК транспозона, она специально кодируется кластером 42AB генома дрозофилы, причем, судя по имеющимся заменам, выглядит считанной в антисмысловой ориентации с фрагментов не современных, а древних транспозонов, являющихся предковыми для современных. Фрагменты древних транспозонов содержатся во многих гетерохроматиновых районах, однако в данном процессе участвуют только последовательности из 42AB-кластера piRNA. Он транскрибируется как единая РНК, которая затем нарезается на piRNA.
В присутствии активного транспозона появляются также piRNA в смысловой ориентации. Они являются фрагментами РНК, считанной с активного транспозона. Предполагается, что эти два класса piRNA – антисмысловой (первичные piRNA) и смысловой (вторичные piRNA) - стимулируют экспрессию друг друга по пинг-понг модели: при этом один класс – первичные - считывается с останков предкового транспозона, а комплементарный ему –– вторичные - с современного активного транспозона. (Здесь есть тонкости, связанные с воспроизводством правильных концов первичных и вторичных РНК, которые не до конца понятны и не очень важны в нашем контексте.) Если же активного транспозона нет, то система с положительной обратной связью не запускается. Можно предположить, что все начинается с некоей слабой базовой экспрессии антисмысловой piRNA; этот транскрипт формируют дуплексы со смысловой мРНК транспозона, которая фрагментировалась бы на отрезки белком из семейства Dicer. Появившись в каком-то количестве, короткие отрезки связываются с белками семейства Argonaut, после чего образуют дуплексы с длинными транскриптами, которые эти белки нарезают на piRNA противоположной орентации. Однако чем именно изначально фрагментируется на первичные piRNA антисмысловой транскрипт кластера piRNA пока неизвестно, этот путь интерференции как минимум не зависит от белков Dicer, задействованных в пути РНК-интерференции через siRNA и miRNA.
Заметим, что аномалии дисгенеза проявляются у взрослых размножающихся мух, а мы говорили о накоплении piRNA в яйце и наследовании их некоторыми клетками бластодермы. Отсюда следует, что «иммунный статус», определяемый наличием piRNA, нарабатываемых за счет «пинг-понг-эффекта», то есть в системе с положительной обратной связью, эпигенетически передается в клетках зародышевого пути, вплоть до образования женских гамет. Казалось бы, здесь не помешала бы РНК-зависимая РНК-полимераза, размножающая как смысловые, так и антисмысловые piRNA, однако она у дрозофилы не найдена. Не исключено, что piRNA сами стимулируют экспрессию кластера 42AB.
У дрозофил существует и еще один известный кластер других piRNA, расположенный в Х-хромосоме и называемый Flamenco. За счет рыхлой структуры своего хроматина он является одной из горячих точек встраивания ретротранспозонов и тем самым превратился в хранилище многих разных ретротранспозонов, то есть по сути антивирусной библиотекой! С него считывается один длинный транскрипт, содержащий фрагменты последовательностей ретротранспозонов преимущественно в антисмысловой ориентации (некоторые - в смысловой) и фрагментируется опять-таки неизвестным белком на короткие piRNA – это называется первичным процессингом, в отличие от пинг-понг механизма, который здесь не работает. В фолликулярных клетках яичников, являющихся частью соматического клеточного пути, они запускают посттранскрипционный сайленсинг, в клетках зародышевого пути – по всей видимости транскрипционный. В соматических клетках дрозофилы интерференция, основанная на piRNA не работает, но работает транскрипционный сайленсинг ретротранспозонов на основе siRNA.
Система защиты от транспозонов, основанная на piRNA – весьма консервативна. Она имеется у всех животных и всегда работает в клетках зародышевого пути, а также в раннем эмбриогенезе (активность этой системы в соматических фолликулярных клетках, которые, однако, тесно взаимодействуют с клетками зародышевого пути, показана только у дрозифолы). Даже косвенные свидетельства существования пинг-понг биогенеза piRNA найдены у губок и кишечнополостных. Считается, что у млекопитающих имеется десятки тысяч разных piRNA – у мыши их найдено болееразных!). У дрозофилы найденоЗаметим, что речь здесь идет именно о готовых коротких РНК, тогда как локусов, с которых транскрибируются их предшественники, не более десятков. Такие локусы традиционно называются кластерами piRNA, их длина может достигать сотен кб и они могут кодировать от нескольких до многих тысяч разных piRNA каждый. Почти все piRNA являются антисмысловыми к мРНК транспозонов, включая ретротранспозоны. (Для одного из их классов был предложен термин rasiRNA - repeat associated small interferring RNA), - который вызвал некоторую путаницу, в данный момент он в основном оставлен.) У млекопитающих piRNA-зависимая интерференция идет по-видимому только в раннем эмбриогенезе и в сперматогенезе, у беспозвоночных - в обоих зародышевых путях. piRNA присутствуют и в цитоплазме, и в ядре и могут участвовать как в посттранскрипционном, так и в транскрипционном сайленсинге, последний вызывает и поддерживает гетерохроматинизацию транспозонов.
Мы видим, как серьезно подошла эволюция к подавлению активности транспозонов в зародышевом пути и раннем эмбриогенезе: piRNA-зависимая интерференция имеет место только там, в соматических тканях с транспозонами борется только только интерференция, зависимая от эндогенных siRNA. Мутации по генам белков piRNA-зависимой системы приводят к стерильности у дрозофилы, но не у мыши, где эта система по всей видимости дублирована альтернативными системами защиты. (Мы с вами помним аналогичные результаты по нокауту самых разных генов у дрозофилы и мыши – у последней молекулярные функции большинства генов оказываются задублированными.)
20.3.4. Индуцированный повторами точковый мутагенез
Во «вводных замечаниях» мы утверждали, что большинство эукариот борятся с нежелательными симтомами присутствия мобильных элементов, но не с самим их присутствием, ввиду огромной технической сложности такой задачи и неоправданно высоких расходов, которые потребовала бы ее реализация. Однако нечто подобное сумели реализовать сумчатые грибы (но не такие их вторично упростившиеся представители, как дрожжи), благодаря чему их геномы содержат очень немного лишней ДНК. Это явление известно для шести их родов и науличшим образом изучено у уже известной нам нейроспоры. Оно называется индуцированный повторами точковый мутагенез . Для него используется англоязычное сокращение RIP, что, например, в англоязычной википедии расшифровывается как repeat induced point-mutation, там же имеется ссылка на статью-первоисточник 1987 года в журнале Cell, откуда мы с изумлением узнаем, что аббревиатура расшифровывается как rearranmegents induced premeiotically (что само по себе несколько misleading, поскольку речь идет о точковых мутациях, а не о том, что мы привыкли называть перестройками). Явление состоит во внесении в множественных транзиций (замен Г / Ц и А / Т) в те участки ДНК, которые присутствуют в гаплоидном геноме более чем в одной копии. Происходит оно на стадии дикариона, то есть после «оплодотворения», но до кариогамии. Гомология повторов, вовлекаемых у такой мутагенез, может быть невысокой – до 80%. Более того, такие мутации вносятся не только в повторы, но и в участки ДНК, вплоть до кодирующих, непосредственно к ним прилегающие, и в тем большей степени, чем ближе они расположены. После внесения таких мутаций, соответствующий участок ДНК часто подвергается метилированию по цитозину, благодаря чему не только перестает быть повтором и не только теряет осмысленную генетическую информацию, если он ее имел, за счет миссенс - и нонсенс-мутаций, но и лишается способности к транскрипции. RIP считается несомненной системой защиты от ретротранспозонов.
Такое явление требует некоего механизма распознания повторов, который пока остается неизвестным. Единственный известный белок, необходимый для RIP, носит название RID и проявляет сходство с ДНК-метилтрансферазой.
Генетики пользуются RIP как источником мутаций. Для этого нужно ввести в геном гриба дополнительную копию какого-то гена, так что после даже одного цикла полового размножения RIP вносит множественные мутации в обе.
20.4. Мобильные генетические элементы в геноме
Ретротранспозоны особенно обильны у растений. У кукурузы до 85% генома, из них 49-78% - ретротранспозоны. У пшеницы – 90% генома – повторы, из них 68% - ретротранспозоны. Поэтому чтение генома пшеницы составляет труднейшую и неблагодарную задачу, поскольку повторенные участки крайне сложно собираются в цельные геномы биоинформатическими средствами и не несут осмысленной информации; дело осложняется тем, что пшеница – амфидиплоид, и в ее гаплоидном геноме присутствуют три очень похожие гомеологоичные хромосомы.
У млекопитающих – 45-48% транспозоны или их остатки.
У человека 44% транспозоны, из них 41% ретротранспозоны, из них 8% - LTR, 20% - LINE (около копий, из которых только 7 000 полноразмерные, и лишь немногие сохраняют активность), 13% - SINE (около 1,5 миллиона копий) и 3% - ДНК-транспозоны.

У человека больше всего Alu-повтора: около 300 пар оснований, около миллиона копий, 10,7% генома. Один из SINE, для транспозиции нужна ревертаза от LINE. Характерен для высших приматов, возник из 7sL РНК рибонуклеопроеидной частицы, распознающий сигнальный пептид белков ЭПР и плазматической мембраны. Содержит остатки двух пептид-кодирующих последовательностей в противоположной ориентации, разделенных АТ-богатой промежуточной последовательностью и полиаденилированный хвост. Источник мутаций, многие случаи рака, один из диабетов, гемофилия. Замены в Alu, приводящие на уровень транскрипции рядом – случаи рака легких, желудка, болезни Альцгеймера.
14 тыс. Mariner-like, склонность к горизонтальному переносу.
У дрозофилы ретротранспозоны занимают около 40% генома, ДНК-транспозоны около 9% генома. SINE отсутствуют.
Гены гетерохроматина обычно имеют огромные интроны, набитые ретротранспозонами.
20.5. Мобильные генетические элементы на службе филогенетики
Ретротранспозоны – прекрасные маркеры общего происхождения, так как их отсутствие всегда есть предковое (плезиоморфное в терминах кладистического анализа) состояние, присутствие – всегда приобретенное свойство (апоморфия), которое не может быть утеряно, а точное попадание туда же (гомоплазия) маловероятно. Китов с бегемотами в парнокопытные, южноамериканская радиация сумчатых с одной ветвью в Австралии и одним представителем вернувшимся.
Один из вариантов RAPD – анализа – электрофоретического анализа длины фрагментов ДНК, апмлифицированных между случайными праймерами. Основан на полиморфизма по местам встроек мобильных элементов – PDR полиморфизм.
20.6. Биологическая роль мобильных элементов
20.6.1. Вводные замечания
Когда мы говорим о биологической роли, мы обычно неявно предполагаем понятие цели, подходя к живым существам как инжинеры и пытаемся ответить на вопрос «бабушка, для чего у тебя такие большие зубы». В принципе, понятие цели применимо к биологическим объектам как к целому если под целью подразумевать увеличение приспособленности, которая едина в трех лицах – жизнеспособность, скорость индивидуального развития и плодовитость. При этом то, что в одних условиях понижает приспособленность, в других может ее повышать. Хуже того: то, что обычно ее понижает – например, мутационная или рекомбинационная изменчивость, иногда может ее повышать – например в условиях быстро меняющейся среды. В частных вопросах устройства биологических систем понятие цели можно условно применять в рамках конкретного технического задания, например – как осуществить перемещение транспозона или как блокировать перемещение транспозона.
Говоря о биологической роли мобильных элементов, нам прежде всего придется рассмотреть их очевидную роль эндогенных мутагенов, которую мы выше признали в целом «отрицательной» - как и мутаций в целом - что, конечно, не исключает положительной роли в качестве случайной или проявляющейся в особых ситуациях. Далее, мы должны рассмотреть участие транспозонов в регуляции генной экспрессии, которая составляет даже и не роль вовсе, а совокупность случайных побочных эффектов. И наконец, мы можем попытаться найти случаи, когда, метафорически выражаясь, эволюция рекрутировала транспозоны для выполнения каких-то конкретных технических заданий.
20.6.2. Роль в спонтанном мутагенезе
Когда открыли, увидели в них основной источник спонтанных мутаций. Считается, что 80% спонтанных мутаций за счет них. Но это если речь идет о мутациях с фенотипом. Рассмотрим подробнее.
Спонтанные точковые замены нуклеотидов возникают минимум на порядок более часто, чем любые перестройки. У человека геномы детей отличаются от геномов родителей (еще вопрос из каких тканей это берется) примерно на сотню замен и примерно десять перестроек, куда относится все, что затрагивает более одного нуклеотида, включая перемещения мобильных элементов, при этом считается, что у млекопитающих каждая особь несет в среднем одну мутацию с фенотипическим эффектом – как правило это слабый количественный эффект. У дрозофилы вне дисгенных ситуаций, просто в изогенных линиях, каждая особь несет в среднем одно новое перемещение Р-элемента, вне какой-либо привязки к функциональным генам и тем более к фенотипу. Если принять кодирующими 1% генома, и учестьгенов у дрозофилы, то вероятность встройки в конкретный ген можно оценить как 1/100 /, то есть 10-6. Это и есть порядок величины вероятности спонтанного мутагенеза по структурным генам, знаменитое 10-6. Мутагенез по количественным признакам больше почти на два порядка, но тут и генов, влияющих на каждый такой признак довольно много, и некодирующая ДНК вовлечена в их регуляцию широко. При этом нужно учесть, что у дрозофилы есть много семейств мобильных элементов. Наиболее активные из них, подобные Р-элементам, как раз и дадут те самые единицы х 10-6.
Перемещение отдельного элемента – редкое событие, и зафиксировать его, следя за мутациями в конкретных генах, очень непросто. Что, однако сделала Мак Клинток.
Иногда спонтанная мутабильность за счет мобильных элементов поднимается до 10-4 под действием разнообразного «стресса» –температура, скрещивание, мутации некоторых генов. Например «гибридизационного стресса». В последнем случае по сути - отсутствуют автономные, а неавтономные могут активироваться автономными при скрещиваниях.
Ретротранспозоны перемещаются очень редко. По сути, речь идет об инфекциях – поначалу множество перемещений, потом подавляется, за счет отбора, после чего сайты на месте стоят десятками миллионов лет, постепенно деградируя вследствие точковых мутаций. Для LTR-ретротранспозонов это почти очевидно, так как они фактически вирусы.
20.6.3. Мусор или оптимально устроенный геном?
Мобильные элементы были классическим примером Junk DNA. Как мы увидим ниже, геном – прежде всего свалка или, если хотите, кладбище ретротранспозонов, на котором ютятся редкие собственные гены организма. (Можно метафорически назвать его кладбищем, если транскрипционный сайленсинг признать за похоронный ритуал.) Эти копии как правило совершенно бесполезны, т. е. представляю собой так называемую мусорную ДНК (junk DNA) и скорее слегка вредны, так как ведут к затратам на вопроизводство ненужной ДНК. Но на шкале приспособленности это оказывается дешевле, чем вырабатывать - ценой селективных смертей – специальный механизм очищения генома от них. Такой механизм, впрочем, существует у сумчатых грибов, геномы которых в основном свободны от мусора.
В то же время мобильные элементы могут поставлять альтернативные промоторы и энхансеры, дулицировать гены в составе сложных SINE и даже осуществлять горизонтальный перенос между неродственными видами (в составе Mariner-подобных транспозонов). Таким образом мобильные элементы могут становиться элементами системы управления генной экспрессии.
В связи с этими соображениями постоянно появляются утверждения о том, что мусорность мусорной ДНК сильно преувеличена и даже что ее не существует вовсе, а вся ДНК так или иначе для чего-то нужна – читай, поддерживается естественным отбором. Надеюсь, вы уже чувствуете, что в прямом смысле это утверждение нереалистично – за любой адаптивный признак нужно платить селективными смертями, так что верным это высказывание могло бы быть лишь в очень переносном или косвенном смысле. С одной стороны, людям по чисто эстетическим соображениям очень хочется, чтобы их геном был сокровищницей информации, а не свалкой, поэтому на такие исследования не жалеют денег, а результаты принимают на ура. Как раз недавно была запущена такая широкая программа, ENCODE – энциклопедия кодирующих элементов человеческого генома. Факты участия мобильных элементов в «осмысленной деятельностьи» - генетической системы управления развитием и жизнедеятельности организмов - имеются и с ними не поспоришь. Однако я бы уподобил наши осмысленные кодирующие гены бомжам, живущим на свалке – они, естественно, приспосабливают разнообразный мусор для своих нужд, и где-то это даже удобно – иной раз чтобы поесть или сделать постель не нужно ничего делать, достаточно протянуть руку. Но свалка остается свалкой, и она очень не похожа на разумно организованное хозяйство, где нет ничего лишнего и все оптимально служит определенной цели, каковым по сути является геном прокариот и даже сумчатых грибов.
В особенности впечатляют размеры геномов у организмов, не испытывающих межвидовой конкуренции – жителей пещер, глубоководных впадин и пр. Отсюда следует, во-первых, что помимо спонтанных делеций, которые, как мы уже знаем, иногда происходят, механизма избавления от дупликаций некодирующей ДНК не существует, а во-вторых, этот неспецифический механизм, поддержанный естественным отбором против больших геномов, все же способен сдерживать распухание генома в условиях межвидовой конкуренции.
20.6.4. Специфические биологические функции, связанные с транспозонами.
Теломераза происходит от ревертазы – и тоже строит ДНК по матрице РНК!
Теломеры дрозофилы – составлены из тандемно расположенных голова к хвосту трех типов non-LTR ретротранспозонов – 6,5, 8 и 10 килобаз, и наращивание теломеры организовано как пристройка ДНК-копий ретротранспозонов.
Оказалось, что у человека неоцентромеры возникают областях, богатых LINE-1, и в неоцентромере они активно транскрибируются, так что их РНК оказывается важным компонентом неоцентромерного хроматина; по-видимому, их транскрипция и устанавливает петлю обратной связи, неоходимую для эпигенетического поддержания центромеры.
Фермент, осуществляющий перестройку генов иммуноглобулинов – родственник транспозазы – вспомним, что он синтезирует теломерный повтор по матрице своей специальной кольцевой РНК.
Хаотичная крапчатая окраска с заданными параметрами. Крапинки на семенах, на яичной скорлупе, на коже, чешуе и шерсти. По закону все или ничего с эпигенетическим поддержанием в клеточных клонах с По большому счету молекулярных механизмов возникновения два – выстригание мобильного элемента и эффект положения.
Последний случай специальной биологической роли мобильных элементов мы рассмотрим отдельно ввиду его фееричности.
20.6.5. Оптимизация соматического генома у инфузорий.
Инфузории живут по заветам Мак-Клинток, осуществляя самый верхний уровень контроля активности генома при помощи мобильных элементов, а также системы борьбы с ними. Наиболее изученными в этом отношении являются три рода – Paramecium, Stylonichia и Oxytricha, принадлежащие к двум разным эволюционным линиям инфузорий, поэтому детали этого механизма у них могут сильно различаться.
Во-первых, при образовании макронуклеуса ДНК рвется на фрагменты от 1-2 Мб у одних до 1-5 кб у других. Считается, что это делается для эффективного деления геномного материала при амитотическом делении макронуклеуса - он просто перешнуровывается как всего навсего мешок с хромосомами, без каких либо механизмов их перераспределения между дочерними ядрами. Во вторых, огромная доля (от 15 до 95%) ДНК микронуклеуса при формировании нового макронуклеуса подвергается элиминации (диминуции). Это в свою очередь делает возможной экономию на умножении лишней ДНК и высокую плоидность именно рабочей ДНК, необходимой для жизнедеятельности, благодаря чему инфузории смогли достичь одних из самых крупных размеров и сложной организации среди одноклеточных.
Длина элиминируемых фрагментов – от 14 пар до десятков кб, общее количество – 5-50 тысяч (у разных инфузорий). В частности, удаляется вся повторенная ДНК из генома микронуклеусов (где она составляет до 20%), в том числе транспозоны, но также много ДНК, не несущей следов происхождения от транспозонов, включая уникальные последовательности. У одних видов, концы такой ДНК – даже той, что представляет собой ретротранспозоны, имеют сходство с концами ДНК транспозонов, таким образом, вырезание элиминируемой ДНК со всей очевидностью произошло от выстригания транспозонов. И действительно, выстригается с висячими концами, закольцовывается, при этом в вырезанной последовательности может сохранять короткие повторы, а может не сохранять. У других видов значение имеет последовательность на некотором отдалении от точек эксцизии. У некоторых видов удаляемая ДНК встречается внутри генов и вырезается очень тщательно – в частности это транспозоны некоторых семейств, у других видов она располагается всегда между генами и вырезается не очень тщательно - в частности, ретротранспозоны, у третьих встречаются оба варианта.
Фермент, который вырезает удаляемые последовательности, есть транспозаза, причем пришедшая от разных транспоонов. У Oxytricha три семейства повторов имеют разные транспозазы, нокаут всех практически блокирует элиминацию, любого из них – несколько снижает, причем показано, что транспозаза одного типа элементов может выстригать другие типы. Таким образом, здесь фрагментация генома – это по сути координированное выстригание множества транспозонов, которое и фрагментирует геном. У других инфузорий ферменты, фрагментирующие геном, уже не является продуктом функциональных транспозонов, но явно происходят от таковых.
Однако то, какие куски ДНК будут элиминированы, а какие останутся, в целом определяет не первичная структура, хотя и ее вклад нельзя отрицать. У Tetrachymena и Paramecium в мейозе чуть не со всего генома микронуклеуса считывается РНК, которые фрагментируются белком из уже знакомого нам семейства Dicer на так называемые сканирующие малые (около 30 н. п.) РНК - scnRNA, весь пул которых по сути составляет копию геном зародышевого пути, которые поступают в цитоплазму и связываются все с теми же белками подсемейства PiWi семейства Argonaut. Они помечают фрагментированные последовательности ДНК будущего макронуклеуса на удаление по гомологии со scnRNA через запуск их гетерохроматинизации. Однако еще до начала формирования будущего макронуклеуса весь пул scnRNA просеивается существовавшим макронуклеусом – если они гомологичны чему-то в его геноме, они с ним связываются и соответствующая ДНК будущего нового макронуклеуса остается непомеченной. Опять мы видим эпигенетически передающуюся информацию, в данном случае о том, какая именно ДНК будет работать. Кстати, легко понять как это исследовалось – в микронуклеус или макронуклеус добавлялись последовательности и следили за тем, войдут ли они в состав макронуклеуса или нет. Причем сравнение с действующим макронуклеусом по всей видимости проходит целиком на уровне РНК – со всего генома макронуклеуса считываются длинные РНК, которые по всей видимости и связывают комплексы scnRNA-PiWi-белки. Фигурально это можно представить как просеивание коротких РНК через сито длинных когда часть из первых прилипает ко вторым.
Можно сказать, что конкретный состав ДНК макронуклеуса поддерживается эпигенетическим путем – хотя опять-таки, наследование черед нуклеиновую кислоту РНК нельзя с полным правом назвать негенетическим. Причем эта по сути соматическая генетическая информация с необходимостью проходит жесткое тестирование естественным отбором.
Поразительно, но у Oxytricha при удалении лишней ДНК та, что остается, часто подвергается закономерным перестройкам, устанавливающим правильный с функциональной точки зрения порядок следования отрезков ДНК. Например, экзоны некоего гена могут быть расположены в микронуклеусе в другом порядке, во встречных ориентациях или на разных хромосомах, но при элиминации эти участки ДНК сшиваются как надо - подчеркнем, что это еще не процессинг РНК, это запрограммированные перестройки ДНК. Оказывается, это происходит по матрице тех самых длинных транскриптов с действующего макронуклеуса, которые имеют правильную ориентацию экзонов!
Весь казус инфузорий показывает нам, что они сумели приручить для целей селекции той ДНК, которая нужна в соматическом пути, как сами транспозоны, используя транспозазы для фрагментации, так и систему борьбы с ними путем сайленсинга соответствующего хроматина – PIWI-белки и scnRNA. Мы в частности убеждаемся здесь, что как сами транспозоны всех основных типов, так и система их сайленсинга сформировались еще у одноклеточных и уже присутствовали полном объеме у общего предка всех ныне живущих эукариот. Очевидно, у инфузорий что все началось именно с сайленсинга определенных участков генома, после чего нужно было лишь «приручить» транспозазу и вырезать эти гетерохроматинизированные участки.
Оборотной стороной этой медали является отсутствие контроля со стороны естественного отбора за геномом микронуклеуса. В атмосфере вседозволенности он работает как насос разнообразной мусорной ДНК. В разных популяций в геном микронуклеуса попадает разный ДНК-мусор, и если представители таких популяций конъюгируют, это не приводит ни к чему хорошему. Специфический для одной популяции элемент генома микронуклеуса может попасть в клетку, где макронуклеус происходит от другой популяции. Поскольку scnRNA производятся во время мейоза, то есть до конъюгации, такая клетка не будет иметь их для элемента генома чуждого микронуклеуса. Поэтому при образовании нового макронуклеуса он не будет помечен на элиминацию и войдет в состав макронуклеуса. Если это большой участок ДНК, не имеющий биологической функции либо даже вредный, клетка потратит большое количество ресурсов на воспроизводство мусорной ДНК или даже погибнет. Такие гибриды будут сильно проигрывать. Поэтому инфузории очень склонны к образованию несовместимых видов-близнецов.
