
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
 БИЛЕТ 3
БИЛЕТ 3
•Химические свойства пептидной связи. Первичная структура белков. Зависимость биологических свойств белков от первичной структуры. Химические взаимодействия, участвующие в поддержании пространственной структуры белков и пептидов (водородные связи, электростатические и гидрофобные взаимодействия)
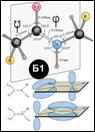
Пептид. связь: част. двойной характер (своб. вращение вокруг С-N-связи): [Б1]. 2 “крайние” формы связи: ·чист. дв. связь (но возможно вращение по C-N-связи) или ·С=N+ (но слишком больш. отриц. заряд на О и N). Истина где-то рядом. !Сα-СО-N-атомы лежат в одной плоскости! На N-атоме преобл.+заряд, а на С-атоме —заряд ®«диполь».
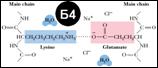
Хим. вз-ия, поддерживающие структуру белка: 1.Водородные связи; 2.Гидрофобные вз-ия (entropically driven – уменьшает вз-ие неполярн. остатков с водой); 3.Электростатические вз-ия [Б4], минимизир-ся выс. конц-ями ионов (мобильнее самих аминок-т); 4.Ван-дер-Ваальсовы вз-ия (дипольные);
 На всякий:
На всякий:
Определение первичн. структуры белка: I.Если белок гетеромультимер, то разделяем полипепт. цепи (мочевина, оч. выс. рН или конц. солей, гуанидин-гидрохлорид). II.Разрыв дисульфид. мостиков (либо надмуравьиной к-той ®обр-ся SO3- ®мостик не восст-ся; либо 2-меркаптоэтанол ®обр-ся SH-группы Cys ®надо остан. обр-ие мостиков ®+йодоацетат). III.АК-анализ с N-конца (Эдмановская деградация) [Б2]. С С-конца пептид фикс-ся на нер-римой основе ®после отщепления отмываем произв-ое ®след. цикл ®опред-ем до 50 послед. аминок-т. Неэффект. для длинных белков. IV.АК-анализ с С-конца – при помощи карбоксипептидаз (разн-ти: А – отщепл. все, кроме Pro, Arg, Lys; В – только для Arg и Lys; C и Y – универс.). V.Фрагментация полипепт. цепи: Трипсин (гидролиз по связям с Arg и Lys ®остаются пепт. с этими АК на С-концах и пептид С-конца всей цепи); Химотрипсин (по аромат. АК – Phe, Tyr & Trp и – слабо – др. АК, т. е.низко специфичен); Протеазы стафиллококка (по Asp & Glu); Цианобромид (CNBr – по остаткам Met); Неспецифич. эндопептидазы (пепсин, папаин и др.); VI.Восст-ие всей АК-послед-ти; VII.Определение места дисульфид. мостиков (не разруш-ся при опр-ии АК-послед-ти ®1 цикл электрофореза ®+надмуравьин. к-та ®еще 1 цикл электрофореза, [диагональный электрофорез]).
 2.Макроэргические соединения. Роль АТФ, УТФ, ГТФ и ЦТФ в различных процессах, протекающих в клетке. Взаимопревращения трифосфатов.
2.Макроэргические соединения. Роль АТФ, УТФ, ГТФ и ЦТФ в различных процессах, протекающих в клетке. Взаимопревращения трифосфатов.
Исследовали мышечные сокращения Þраспад АТР и креатинфосфата. Ресинтез – за счет окислительных процессов в мышце.
«Макроэргические» соединения – не только АТР и др. фосфаты, но и тиоловые эфиры СоА (i. e. ацетил-СоА), ацилпереносящий белок, эфиры аминокислот, S-аденозилметионин (при переносе метильной группы), UDP-глюкоза (при синтезе гликогена).
См.таблицу (энергия гидролиза): в-во может быть донором высокоэнерг. фосфата для соед-ий, располож.ниже в таблице.
Источники макроэрг. связи в фосфатах: 1.Окислит. фосфорилирование; 2.Гликолиз (2 связи; р-ции, катализируемые фосфоглицераткиназой и пируваткиназой); 3.Цикл Кребса (1 связь; сукцинилтиокиназа).
Фосфагены – «резервуары» высокоэнергет. фосфатов. Поддерж. конц-ию АТР при его быстром расходовании, а при избытке – обр-ся креатинфосфат.
!Сам «высокоэнергет.» фосфат в своб. состоянии не присутствует – всегда в составе какого-нибудь соединения.
Взаимопревращение макроэргических фосфатов – за счет фермента аденилаткиназы (миокиназы): АТР +АМР Û2АDР. Функции: 1.Синтез АТР из АDР; 2.Утилизация АМР; 3.Повышает конц-ию АМР при снижении уровня АТР (повышение уровня АМР – сигнал к усилению катаболических процессов Þусилению обр-ия АТР).
!Говорим о роли промежуточного соединения АТР с Mg2+!
Пирофосфат – обр-ся при гидролизе АТР до АМР. PPi расщепл-ся неорганической пирофосфатазой (например при активации длинноцепочечных жирн. к-т под действием ацил-СоА-синтетазы: АТР +СоАSH +RCOOH ÞАМР +РРi +RCO-SH; энергия рассеивается в виде тепла).
Нуклеозиддифосфаткиназа Þсинтез нуклеозиддифосфатов с неадениновыми основаниями:
ADP +UDP ÛADP +UTP (уридинтрифосфат)
ATP +GDP ÛADP +GTP (гуанозинтрифосфат)
ATP +CDP ÛADP +CTP (цитидинтрифосфат)
3.Сверхспирализация ДНК. ДНК-топоизомеразы.
Суперспиральность-когда замкнут кольцо сворачив вокруг собствен оси или когда скручив участок линей ДНК, концы кот зафиксированы. Е перехода мол ДНК к др типу надмолекулярн стр-ры мод пониж за счет образов участков отрицат скручивания. Один из таких переходов- разделение цепей при подготовке к репликации и транскрипции. Вот почему суперспирализац ДНК выгод в биолог сист. Фермен, катализир-ие топологич измен мол ДНК – топоизомеразы.(наибол изучен-бактериальная гираза, инициирующая образ отриц супервитков.)
