
НЕХРОМОСОМНОЕ (ЦИТОПЛАЗМАТИЧЕСКОЕ) НАСЛЕДОВАНИЕ
Для того чтобы та или иная структура клетки могла играть роль материального носителя наследственности и обеспечивать количественные закономерности наследования, она должна обладать тремя свойствами: выполнять жизненно важные функции в метаболизме клетки, обладать способностью к самовоспроизведению, точно распределяться в дочерние клетки при делении. Этим трем условиям полностью удовлетворяют структуры ядра - хромосомы.
Многие органеллы цитоплазмы удовлетворяют первому условию. Например, центриоли участвуют в образовании веретена деления клетки, пластиды обеспечивают синтез органических веществ, митохондрии являются дыхательным центром, в рибосомах синтезируются белки и т. д. Пластиды, митохондрии и центриоли обладают способностью
к саморедукции, то есть удовлетворяют второму условию. Однако, ни одна из органелл цитоплазмы, исключая центриоли, не распределяется при делении клетки так точно, как хромосомы. Именно в этом состоит главное отличие ядерных структур
от цитоплазматических.
Кроме того, есть еще два существенных различия между ядром и цитоплазмой:
1)ядро содержит ограниченное и характерное для каждого вида число хромосом; в цитоплазме много однозначных органоидов, число их, как правило, не постоянно;
2) ядро в большинстве случаев не способно исправить и заменить возникшие дефекты хромосом, они воспроизводятся при делении клетки. Поврежденные органеллы цитоплазмы могут быть замещены путем размножения неповрежденных одноименных структур.
Приведенные различия в свойствах хромосом и органелл цитоплазмы должны обуславливать и различия в закономерностях наследования, определяемых этими элементами клетки.
Наследование, определяемое хромосомами, получило название ядерного
или хромосомного. В тех же случаях, когда материальной основой наследования являются элементы цитоплазмы, оно называется нехромосомным или цитоплазматическим.
Так как яйцеклетка содержит много цитоплазмы, а сперматозоид почти лишен ее, следует ожидать, что цитоплазматическое наследование, в отличие от хромосомного, должно осуществляться по материнской линии.
Так как для органоидов цитоплазмы нет точного механизма распределения при делении клеток, то цитоплазматическое наследование не может характеризоваться такими строгими количественными закономерностями, как ядерное.
Пластидное наследование
Пластиды - своеобразные лаборатории синтеза углеводов. Они содержат ДНК
и размножаются делением. Совокупность пластид клетки как структур, передающих наследственную информацию, называется пластидом, а совокупность генов пластид пластом.
В 1909 г. Баур и Корренс впервые сообщили о фактах пластидного наследования. Корренс изучал наследование пестролистности у ночной красавицы. У этого вида встречаются пестролистные растения, которые имеют в точках роста разные группы клеток:
с нормальными пластидами и с пластидами, неспособными к образованию хлорофилла. Вследствие этого на растении образуются чисто зеленые и абсолютно белые ветви. Семена, полученные с белых ветвей, дают нежизнеспособные всходы, так как у них не идет процесс фотосинтеза.
При опылении цветков с пестролистных ветвей пыльцой от цветков с зеленых ветвей
и при обратном скрещивании результаты получаются разные.
♀ пестролистное х ♂ зеленое: F- зеленые, пестролистные, белые
♀ зеленое х ♂ пестролистное: F- все зеленые
Цветки с зеленых ветвей дают только зеленое потомство, а с белых – только белое (нежизнеспособное), независимо от того пыльцой какого растения они опылялись.
На основании этого был сделан вывод, что наследование пестролистности связано с передачей и распределением при клеточном делении двух типов пластид – зеленых и неокрашенных. Пластиды передаются яйцеклеткой, в результате чего наследование осуществляется по материнской линии. У некоторых видов (герань) пластиды передаются не только яйцеклеткой, но и спермием, поэтому у них пестролистность наследуется по обеим линиям.
Наследование через митохондрии
Митохондрии обеспечивают дыхание клеток эукариотических организмов. Подобно хлоропластам – это самовоспроизводящиеся полуавтономные органеллы клетки, содержащие кольцевые молекулы ДНК.
ДНК митохондрий по нуклеотидному составу и вследствие этого по плотности отличается от ДНК ядра. Митохондрии имеют свой собственный аппарат белкового синтеза, отличающийся от цитоплазматического и близкий к аппарату белкового синтеза прокариот.
Первые сведения о признаках, контролируемых митохондриями, были получены у дрожжей сахаромицетов (Saccharomyces cerevisiae) в конце 40-ых годов ХХ века в лаборатории Б. Эфрусси. Были обнаружены штаммы дрожжей, которые спонтанно образуют карликовые колонии с дыхательной недостаточностью. Поскольку колонии возникают при вегетативном размножении гаплоидных дрожжей, они были названы вегетативными карликовыми. Наряду с вегетативными карликовыми колониями была обнаружена форма, по фенотипу – росту и дыхательной недостаточности – сходная с первой, но она давала расщепление по признаку карликовости, как будто он определяется одним ядерным геном. Эта форма была названа расщепляющимся карликовым штаммом.
Генетический анализ вегетативного и расщепляющегося карликового штаммов показал, что фенотип расщепляющейся карликовости определяется ядерным геном, поскольку при скрещиваниях наблюдается расщепление аскоспор в отношении 1:1. (Сахаромицеты относятся к Аскомицетам, или Сумчатым грибам. Это гаплобионты. Сразу после оплодотворения у них происходит мейоз с образованием гаплоидных спор – аскоспор). При скрещивании вегетативных карликов и нормальных дрожжей диплоидная зигота, в которой есть митохондрии от нормальной формы, не дает расщепления – из аскоспор не появляются мелкие колонии. Следовательно, у этих форм геномы одинаковы, различается только цитоплазма. Расщепление по типу цитоплазмы в мейозе не происходит.
Получено и прямое доказательство роли митохондрий в наследственной
передаче дыхательной недостаточности у дрожжей. Вегетативных карликов, лишенных клеточных стенок, выращивали в присутствии изолированных митохондрий нормальных дрожжей. В результате часть образовавшихся колоний имели нормальные размеры. Этот факт можно объяснить тем, что «нормальные» митохондрии, попав в клетки вегетативных карликов, исправили дефект их дыхательной системы и, передаваясь из клетки в клетку в ходе деления, способствовали образованию нормальных колонии.
Цитоплазматическая мужская стерильность (ЦМС)
Это наиболее хорошо изученный пример цитоплазматического наследования. Обнаружена у кукурузы, лука, свеклы, льна и других растений. ЦМС была открыта у кукурузы в 30-ых годах ХХ Хаджиновым в СССР и Родсом в США.
Кукуруза - однодомное растение. Женские цветки собраны в початок, мужские в метелку. У некоторых сортов обнаружены растения, имеющие в метелках недоразвитые пыльники, часто совсем пустые, иногда с недоразвитой стерильной пыльцой. Оказалось, что этот признак определяется особенностями цитоплазмы. Опыление растений с мужской стерильностью нормальной пыльцой с других растений в большинстве случаев дает в потомстве растения со стерильной пыльцой. При повторных скрещиваниях признак не исчезает и передается по материнской линии. Цитоплазма, обуславливающая стерильность пыльцы, была обозначена символом цитS (или cytS), а цитоплазма с нормальной фертильностью (плодовитостью) – цитN (или cytN).
Установлено, что генотип растения может оказывать влияние на действие стерильной цитоплазмы. Цитоплазма цитS может обуславливать стерильность пыльцы только при наличии в генотипе растения в гомозиготном состоянии рецессивного гена rf. Если же ген представлен доминантным аллелем Rf, то растения цитS RfRf и цитS Rfrf имеют нормальную пыльцу. Таким образом, стерильную пыльцу могут иметь растения цитS rfrf, а фертильную цитN rfrf, цитN Rf - и цитS Rf-. Ген Rf не изменяет структуру стерильной цитоплазмы,
а тормозит проявление ее действия.

Предетерминация цитоплазмы
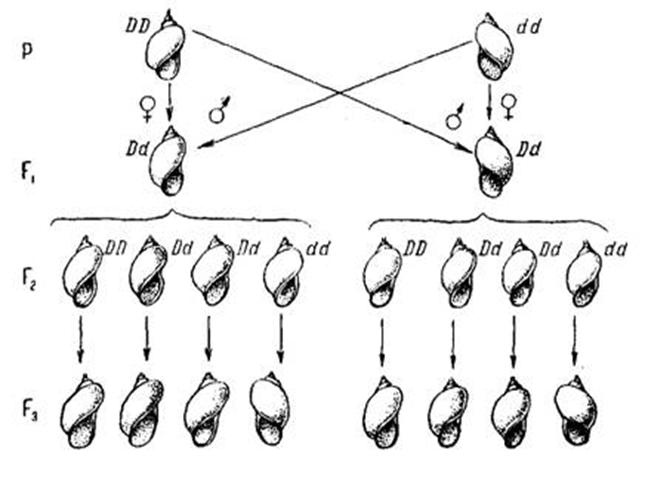
У прудовика Limnaea (лимнеи) существуют особи с правозакрученной и левозакрученной раковинами. Правозакрученность определяется доминантным геном D, левозакрученность - его рецессивным аллелем d.
Реципрокные скрещивания: ♀ DD x ♂ dd и ♀ dd x ♂ DD дают различные результаты. В первом скрещивании все потомки "правые", во втором - все "левые". Так как данный вид гермафродитен, то у него возможно самооплодотворение. В F2 от обоих скрещиваний путем самооплодотворения все потомки имеют правозакрученную раковину. В F3, также полученном путем самооплодотворения, расщепление в соотношении 3 "правых" :
1 "левой". Проявление признака как бы отстает на одно поколение от генетической формулы. Решающую роль в таком отставании играет генотип материнского организма, определяющий свойства цитоплазмы яйцеклетки, от которой и зависит направление закручивания раковины. Этот признак формируется в раннем эмбриогенезе направлением веретена второго дробления. Такое явление получило название предетерминации цитоплазмы генотипом материи, или материнского эффекта.
1) Р: ♀ DD x ♂ dd
F1: Dd
прав.
F2: DD 2 Dd dd
прав. прав. прав.
i i i
F3: DD 2 (DD:2Dd:dd) dd
прав. Прав. Лев. - 3 : 1
2) Р: ♀ dd x ♂ DD
F1: Dd
лев.
F2: DD 2 Dd dd
прав. прав. прав.
i i i
F3 DD 2(DD:2Dd:dd) dd
прав. прав. лев. - 3 : 1
Наследование паразитов и симбионтов
В клетке могут присутствовать некоторые необязательные для нее элементы -
вирусоподобные частицы, плазмиды. Эти частицы придают клеткам определенные свойства и могут передаваться другим клеткам.
У инфузории парамеции аурелии существуют линии-убийцы, выделяющие токсин парамеции, безвредный для его продуцентов, но убивающий других, чувствительных к нему особей данного вида.
В цитоплазме парамеций-убийц есть каппа-частицы, обычно не передающиеся при конъюгации, так как при этом происходит обмен ядрами, а не цитоплазмой. При задержке расхождения конъюгирующих клеток они обмениваются цитоплазмой. Тогда каппа-частицы могут передаваться чувствительным партнерам, и они тоже становятся убийцами. Каппа-частицы это один из видов бактерий-эндосимбионтов парамеции. Их можно культивировать вне клетки и заражать ими парамеций, лишенных бактерий.
Эндосимбионты широко распространены у простейших. Они могут жить не только в цитоплазме, но и в макро - и микронуклеусе. Клетки с эндосимбионтами в ядрах не способны к половому процессу.
Тесная связь между функциями ядра и цитоплазматическими эндосимбионтами была продемонстрирована в конце 60-ых годов ХХ века на амебе протей. Культура амебы была случайно заражена бактериями. Большинство амеб погибло, но часть из них выжила и активно делилась в присутствии бактерий. Число бактерий в амебе достигалоЯдро простейшего стало зависимым от бактерий. Пересаженное в другую амебу ядро уже не могло обеспечивать функционирование и деление цитоплазмы в отсутствии бактерий. Такие факты используют для подтверждения гипотезы о симбиотическом происхождении эукариотических клеток. В частности о бактериальном происхождении митохондрий и хлоропластов.
У дрозофилы известны линии без самцов. Самки этих линий при скрещивании
с любыми самцами дают в потомстве только самок. Выяснилось, что безсамцовые линии заражены спирохетами, которые, проникая в откладываемые яйца, убивают мужские эмбрионы, но не убивают женские. В результате самки становятся носителями инфекции.
К факторам нехромосомного наследования относятся плазмиды. Плазмида - это генетическая структура цитоплазмы бактерий и некоторых простейших, представляющая собой небольшую кольцевую молекулу ДНК, не связанную с хромосомой. Различают три типа плазмид.
1. Факторы генетического переноса.
Обладают генами переноса и репликации. Под контролем генов переноса синтезируются особые белки, осуществляющие транспорт плазмид из клетки в клетку. Бактерии, содержащие плазмиды, служат генетическими донорами.
2. Конъюгативные плазмиды.
Представляют собой фактор переноса, сцепленный с плазмидными генами, контролирующими синтез белков, имеющих значение для бактерий. Например, плазмиды R контролируют синтез ферментов, придающих бактериям устойчивость (резистентность) к антибиотикам, сульфаниламидам и другим лекарственным веществам.
3. Неконъюгативные плазмиды.
Не передаются другим клеткам, так как не содержат фактор переноса. Обеспечивают определенные свойства бактерий. Однако передача их другим бактериям возможна, если они мобилизуются на перенос другой (конъюгативной) плазмидой.
Эписомы - это плазмиды, которые могут существовать автономно, а также встраиваться в геном бактерии.
Факторами неядерного наследования могут служить некоторые вирусные частицы, например, ретровирусы. Переносить информацию могут также “прыгающие гены” (мобильные генетические элементы).
