


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
С. П. РОМАНОВ
Институт физиологии им. РАН, Санкт-Петербург
*****@***edu
ОТ НЕЙРОНА К СТРУКТУРНО-ФУНКЦИОНАЛЬНОМУ АНАЛИЗУ ИНТЕГРАТИВНОЙ АКТИВНОСТИ
ЦЕНТРАЛЬНОЙ (МОТОРНОЙ) НЕРВНОЙ СИСТЕМЫ
Рассматриваются свойства электронного аналога нейрона, являющегося основой модели нейронной сети сегментарного уровня управления мышечным сокращением. Показано, что передать информацию и использовать её для управления можно только в кольцевых нейронных структурах. Основываясь на принципах кольцевой регуляции и иерархической организации системы управления, анализ моторного выхода в исследовании на человеке позволил выявить параметры циклической активности, соотносимые со структурными уровнями регуляции моторной системы.
Ключевые слова: электронный аналог нейрона, структура и функция нейронной сети, анализ временных рядов
Введение
Усложнение объектов управления в направлении одновременного контроля многих переменных и увеличивающиеся объёмы переработки разнообразной информации от многочисленных датчиков “очувствлённых” исполнительных органов робототехнических комплексов заставляют искать приемлемые технические решения, заимствуя их у биологических систем управления. Для создания управляющих нейронных сетей, в отличие от формализации процессов переработки информации в зрительной и слуховой сенсорных системах, возможно напрямую использовать достижения нейронаук в раскрытии функции связей между отдельными нейронами и выявляемыми закономерностями передачи и преобразования импульсных сигналов между структурными образованиями моторной системы для решения задач регуляции и многопараметрического управления.
Обширная нервная система формируется после возникновения особых нервно-мышечных клеток, передающих сенсорный сигнал непосредственно на эффектор, как у сохранившихся кишечнополостных, оснащённых органами зрения и равновесия. Наличие только сенсорных и эффекторных органов без обширной нейронной сети уже обеспечивает многообразие поведения в окружающей нестационарной среде, что эффектно было показано в 60 годах прошлого века на кибернетических моделях черепах и мышей. Но наличие только сенсорных и исполнительных органов не может обеспечить разумное поведение, для осуществления которого необходим интеллект, способный ставить цели перед организмом или роботом. В попытке найти такой интеллект в нейронах коры головного мозга в 2005 году был запущен двухгодичный проект (Blue Brain Project) по компьютерному моделированию структуры корковой колонки крысы, который продолжается до сих пор с наращиванием вычислительной мощности для моделирования процессов в мозге на молекулярном уровне [1].
Цель нашего исследования состоит в том, чтобы понять процессы преобразования сигналов в нервной системе и выявить соотношения между структурой связей и функцией организованной сети нейронов.
Объекты исследования и методы
Объектом исследования была избрана система управления движениями, т. к. при изучении её механизмов в отличие от сенсорных систем всегда можно чётко сопоставить контролируемые вход и выход для выявления принципов преобразования сигналов внутри и между её элементами. Нейрофизиологические исследования не позволяют изучать в достаточно полной мере ни взаимодействие отдельных механизмов управления, ни их роль при совершении произвольных двигательных актов. В то же время исследование поведения целостной структуры при попытке выявить нейрофизиологические механизмы управления остаётся за порогом методических возможностей экспериментатора. Для достижения цели мы применили аналоговое и компьютерное моделирование нейронных структур системы управления движениями и исследовали особенности формирования моторного выхода (движения) на животных и человеке [2-4].
Электронный аналог нейрона
Какой мы видим структурную организацию нервной системы? Прежде всего, мы исследовали преобразования импульсных потоков в простых нейронных сетях на электронных аналогах нейронов, работающих в реальном масштабе времени [5-7]. Функция сети зависит от свойств её элементов. Бинарных свойств генерации импульса нервной клеткой и суммации сигналов было явно недостаточно в электронном аналоге для получения присущих нейрону свойств преобразования импульсных сигналов. Нами впервые было введено в модель нейрона представление о синапсе как ключе. Широко обсуждаемое в нейрофизиологической литературе, оно не нашло отражения в модельных исследованиях и, тем более, в элементах искусственных нейронных сетей. Развиваемый нами постулат о том, что сами входные сигналы управляют замыканием и размыканием "ключа", причем неактивные в данный момент входы не влияют на процессы суммации, происходящие на мембране нейрона, привел к созданию электронного аналога нейрона, в котором учтены процессы выделения и распада медиатора и промоделирована функция синапса (рис. 1).
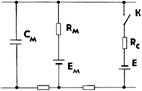
а б
Рис. 1. Упрощенная электрическая схема мембраны нервной клетки (а)
и реализация функции синапса (б) в электронном аналоге нейрона
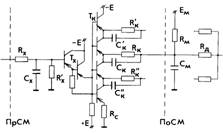
Электронная схема одного входа модели реализует функцию ионного канала в области синапса, которую можно сравнить с эквивалентной электрической схемой мембраны и синаптического входа К. Общим для этих двух схем является наличие сопротивления Rm и ёмкости Cm мембраны нервной клетки с источником Em, создающим внутриклеточный потенциал, и ключа К (Тк) с сопротивлением Rc и источником E, отражающими функцию синапса. На нашей схеме параметры Rк и Cк представляют эквивалентные сопротивления и ёмкость ионного канала, шунтирующие сопротивление Rm мембраны при замыкании ключа. Отношение Rm/Rк соответствует весу входа по данному ионному каналу, а цепочки Rк'Cк' и Rк"Cк" отражают свойства других ионных каналов синапса, которые в общем случае могут подключаться к другим цепочкам RmCm, напряжение с выходов которых, формируя ВПСП или ТПСП, суммируются через сопротивление Rд дендрита или тела клетки в области ее генераторной зоны. Управление электронным ключом Tк осуществляется самим входным сигналом, т. е. напряжением с конденсатора Cх, имитирующим накопление и воздействие химического передатчика на постсинаптическую мембрану (ПоСМ). Постоянные времени RхCх и R'хCх входной цепи модели соответствуют времени выделения и распада медиатора в синаптической щели. В предложенной модели ионного канала количество Kn медиатора, находящееся в синаптической щели после прихода n-го импульса к пресинаптической мембране (ПрСМ), может быть вычислено по формуле
Kn(t)=((K-Kn-1)*(1-exp(-t/RxCx))+Kn-1)*exp(-(t-t')/R'xCx),
где t – текущее время после прихода очередного n импульса длительностью t' по аксону данного входа, K – максимальная концентрация медиатора, Kn-1 – остаточное количество медиатора в синаптической щели. Количество медиатора в синаптической щели пропорционально частоте следования импульсов по данному входу.
Далее идут нелинейные преобразования, связанные с подключением входа: открывание ионного канала, когда резисторы Rк шунтируют сопротивление Rm мембраны, и регулирование текущей концентрацией медиатора тока через ионный канал. Для отождествления свойств ионного канала на модели, измеряли напряжения в следующих точках схемы: напряжение на конденсаторе Cx, соответствующее количеству выделяемого в синаптическую щель медиатора, напряжение на конденсаторе Cm, соответствующее внутриклеточному потенциалу нейрона, и напряжение на входе ионного канала - на резисторе Rк со стороны ключа Tк. Результаты статических измерений представлены графиками на рис. 2.
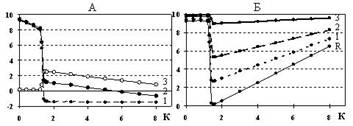
Рис. 2. Характеристики синаптического входа аналога нейрона:
А: 1 – напряжение на “ключе”, 2 – разность напряжений на постсинаптической мембране, 3 – ток через ионный канал;
Б: изменение внутриклеточного потенциала для разных значений сопротивления мембраны, характерных для маленьких (1), средних (2) и больших (3) нейронов, R – сопротивление канала. По оси абсцисс: концентрация медиатора. Все величины в вольтах
Напряжение 1 на “ключе” пропорционально разности напряжений между интегратором Cx и резистором Rк на выходе ионного канала. Если концентрация медиатора мала, то ключ обладает большим сопротивлением (сопротивлением утечки), незначительно снижающим величину мембранного потенциала. При пороговом значении концентрации медиатора напряжение на ключе резко падает и становится отрицательным, поддерживаемым на постоянном уровне несмотря на дальнейшее увеличение количества медиатора. "Ключ" превращается в активный генератор тока, обеспечивающий насосную функцию, т. е. направленное движение ионов через ПоСМ. Напряжение 2 на ПоСМ уменьшается скачком в момент открывания канала и продолжает убывать при дальнейшем увеличении концентрации медиатора, становясь отрицательным (инверсия тока) у достаточно малых нейронов. Ток 3 ионного канала резко возрастает в момент открывания ключа, а затем линейно убывает по мере увеличения концентрации медиатора. Напряжение 2 на ПоСМ определено через разность между внутриклеточным потенциалом и концентрацией медиатора, а изменение тока 3 ионного канала вычислено через разность напряжений на его входе и выходе. Изменение внутриклеточного потенциала также носит ступенчатый характер, но после резкого спада в результате открывания канала внутриклеточный потенциал увеличивается пропорционально росту концентрации медиатора (графики Б). Глубина спада полностью определяется отношением сопротивлений ионного канала и ПоСМ. Чем меньше это отношение, тем больше перепад внутриклеточного потенциала в области данного синаптического входа и, следовательно, тем больше его вес и влияние на генераторный механизм клетки. Количество ионных каналов определяется свойствами мембраны нервной клетки и размером синапса. Параллельное их подключение на модели способствует увеличению веса входа, а различные параметры сопротивления и емкости могут моделировать участие каналов различных ионов в формировании ВПСП или ТПСП. Электронно-дырочная проводимость твёрдого полупроводника позволила промоделировать процесс передачи возбуждения через мембрану нервной клетки, обеспечив пластические свойства преобразования импульсных потоков в “жесткой” структуре связей. В частности воспроизведены "on" и "off" ответы и трансформация ритма только при изменении интенсивности воздействий по параллельным входам (рис. 3).

Рис. 3. Реакции аналога нейрона на возбуждение по двум (1 и 2) входам:
А, Б и В – частота последовательности импульсов по входу 1 соответственно
6, 8 и 50 Гц; 3 – внутриклеточный потенциал, 4 – выход аналога нейрона
Исследование электронного аналога показало адекватность его поведения регистрируемым реакциям биологических нейронов. Модель стала некритичной к числу входов (их может быть столько, сколько необходимо для организации связей в нейронной сети) и осуществляла процессы суммации сигналов со средним только по числу активных в данный момент входов, сохраняя широкий диапазон генерации импульсов независимо от числа возбужденных входов. Важным результатом исследования взаимодействия двух и более нейронов был вывод о невозможности передачи структуры импульсного потока по разомкнутой цепи нейронов вследствие трансформации ритма на выходе каждого нейрона в соответствии с его свойствами. Именно паттерны импульсов в кольцевой структуре являются носителями информации. В качестве примера приведём осциллограммы импульсных потоков “большого” и “маленького” нейронов, соединённых по типу “мотонейрон – клетка Реншоу” (рис. 4).
При малой (А) частоте возбуждения первого нейрона его разряды хаотичны, и малый нейрон на каждый импульс разряжается пачкой импульсов. При увеличении (Б и В) возбуждения второго нейрона происходит трансформация входной последовательности импульсов в собственную ритмику на выходе каждого нейрона. В случае замыкания тормозной связи с выхода нейрона 1 на нейрон 2 (осциллограммы б) генерация импульсов на выходах нейронов становится взаимозависимой и структурируется в характерные паттерны циклической активности.

Рис. 4. Активность последовательно соединённых “большого” (2) и “малого” (1) нейронов в разомкнутой (а) цепи и при замыкании (б) тормозной обратной связи
с выхода нейрона 1 на второй вход нейрона 2 при возбуждении (А, Б, В) первого входа нейрона 2 с частотой соответственно 20, 25 и 40 Гц
При большой частоте возбуждения (В) нейрон 2 может генерировать два разряда с коротким межимпульсным интервалом, которые вызывают более длительные пачки импульсов нейрона 1 и более длительное торможение нейрона 2. Таким образом, существенной в плане считывания состояния активности в нейронной цепи и, следовательно, передающей информацию является циклическая активность в кольцевой структуре в отличие от ритмической активности на выходе каждого нейрона.
Модель регуляции мышечного сокращения
Дальнейшее изучение закономерностей трансформации сигналов и передачи информации в нейронных сетях целесообразно было проводить только в условиях управления каким-либо объектом. Т. к. естественным объектом управления для нервной системы живых организмов служит локомоторный аппарат, необходимо было создать электронные аналоги мышц с обширным рецепторным аппаратом и объединить их вместе с аналогами нейронов в единый работающий в реальном масштабе времени исследовательский комплекс [8-12]. Временному и структурно-функциональному соответствию элементов системы управления движениями в модели и объекте моделирования придавали первостепенное значение (рис. 5).
Промоделировав сегментарные структуры управления мышечным сокращением, мы пришли к заключению, что многочисленные обратные связи от мышечных рецепторов к мотонейронам собственной мышцы представляют гомеостатический механизм (в терминологии [13]) поддержания её длины или усилия. Этот механизм не имеет самостоятельного управляющего значения, но превращает биомеханически нестабильный локомоторный аппарат в управляемый для надсегментарных структур ЦНС.
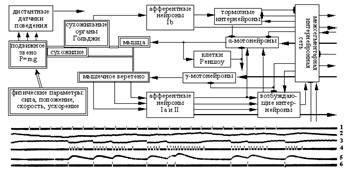
Рис. 5. Структурно-функциональная схема электронного аналога мышцы
и осциллограмма воспроизводимых в модели сигналов:
1 – последовательность импульсов на входе α-мотонейрона;
2 – его внутриклеточный потенциал; 3 – внутриклеточный потенциал клетки Реншоу; 4 – импульсы на её выходе; 5 – сокращение мышцы; 6 – импульсы
на выходе мотонейрона; двойные линии характеризуют механические связи
в системе
Кроме того, показали, что в кольцевых структурах гомеостатированы свойствами элементов кольца паттерны активности, циклически повторяющиеся со свойственной каждому элементу кольца ритмикой. С этих позиций в каждой кольцевой структуре нет неустойчивых состояний. Каждому состоянию распространяющихся в кольце потоков, определяемому внешним (через синапсы) возбуждением или их модуляцией химическим составом окружающей нейроны среды, соответствует свой паттерн активности на каждом из элементов структуры и единая циклическая частота. Поведение модели сегментарного уровня управления мышечным сокращением адаптивно, а нейронная сеть обеспечивает автоматическую регуляцию состояния моторного выхода. Нейронными механизмами проприоцептивных рефлексов минимум три регуляторных кольца осуществляют гомеостатирование двигательной функции. Замыкание всех трёх колец на один α-мотонейрон (или множество нейронов в мотонейронном пуле в случае рассмотрения множества двигательных единиц) повышает надёжность поддержания гомеостаза. На сегментарном уровне гомеостатированы импульсные потоки в структуре в соответствии с влияниями от мышечных рецепторов растяжения и управляющими сигналами от вышележащих структур мозга. Спинальные механизмы регуляции через проприоцептивные пути обеспечивают компенсацию ограниченных по интенсивности возмущений. Внутрисегментарная нейронная сеть осуществляет координацию работы мышц одной конечности, а межсегментарные связи обеспечивают межконечностные взаимодействия. Гомеостатирование в широком диапазоне изменений внешних нагрузок возможно только при участии супраспинальных структур (мозжечок, ствол, подкорковые структуры мозга и кора больших полушарий). Можно наращивать модель всё более высокими контурами регулирования, но нервная система всё равно остаётся автоматом, компенсируя изменение какого-либо параметра по принципу электромеханического гомеостата.
Исследование моторного выхода у человека
Изучая биомеханику движений, установил принципы организации моторной системы для “построения движений”, которые сейчас широко используются в объяснении когнитивных функций. Это, прежде всего, необходимая иерархическая многоуровневая организация и кольцевое управление или регуляция с использованием сенсорных коррекций. Чтобы выявить параметры кольцевой активности в ЦНС, нами предложено анализировать изометрическое усилие, длительное удержание которого сопровождается пропорциональным силе возбуждением всех структур моторной системы (рис. 6).

Рис. 6. Непроизвольные колебания изометрического усилия относительно удерживаемого уровня, возрастающего (от 1 к 4) в диапазоне 0-4 кг
Современные методы математического анализа позволяют разложить временные ряды на главные компоненты (ГК), выделяя в них циклическую активность и другие составляющие изучаемых процессов. Можно было бы представить моторную систему “черным ящиком”, но её организация в настоящее время хорошо известна, хотя функционирование целостной системы остаётся на уровне гипотез. На рис. 7 представлена структурно-функциональная схема основных уровней организации моторной системы в сопоставлении с выделенными ГК циклической активности.
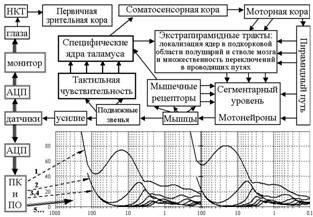
Рис. 7. Соотнесение огибающих главных компонент разложения изометрического усилия как моторного выхода с кольцевыми структурами, сформированными
в моторной системе
У здоровых испытуемых все ГК имеют хорошо выраженные максимумы, расположенные относительно превалирующей частоты, характерной для данного уровня. Первую ГК с наибольшими максимумом и частотой относим к сегментарному уровню. Вторую ГК связываем с участием сенсомоторной коры и пирамидного тракта, ведущим целенаправленные движения. Максимумы первых двух компонент смещаются по амплитуде и частоте однонаправлено в наибольшем диапазоне по сравнению с максимумами следующих ГК. Их мы относим к экстрапирамидной системе, имеющей ограниченное число нисходящих к спинному мозгу трактов. Если принадлежность первых двух ГК не вызывает сомнения, то верификация следующих ГК требует проведения исследования на пациентах с известной этиологией центральных нарушений построения движений.
Заключение
Впервые неинвазивным методом показано вовлечение в гомеостатическую регуляцию всех структур моторной системы. Нейроны коры головного мозга создают в структуре корковых колонок динамический образ конкретного входа с возможностью предсказания его изменений на некоторый интервал времени для компенсации задержек проведения возбуждения в восходящих и нисходящих трактах. Подкорковые структуры на основе активности нейронов в колонке корректируют состояние нейронов моторной коры, где формируется нисходящая команда к исполнительным органам и сохраняется состояние, требуемое для решения задачи. Но где формируются цели нашего поведения, соответствующим образом подготавливающие различные отделы ЦНС к их реализации? Выявленный принцип организации управления мышечным сокращением может быть реализован в сложных робототехнических комплексах.
Список литературы
1. http://bluebrain. epfl. ch/
2. Романов электрической активности различных групп мышц у крыс во время локомоции // Физиол. журн. СССР, 1978. Т.64, № 10. С. .
3. Романов информации в нейронной структуре модели спинального уровня управления мышечным сокращением // Проблемы физиологии движений / Под ред. . Л.: Наука, 1980. С.104-122.
4. Романов элементов системы управления движением // Средства автоматизации физиологических исследований / Под ред. . Л.: Наука, 1988. С. 150-165.
5. Романов механизмов спинального уровня управления мышечным сокращением: Автореферат дис. канд. биол. наук. Ин-т физиологии им. РАН. Л., 1974.
6. Романов нейрона: SU № 1978.
7. Романов свойств ионного канала и исследование его роли в формировании импульсной активности нейрона // Нейрофизиология, 1989. Т. 21 : 3, с. 379-389.
8. Романов организации синаптического аппарата на формирование и передачу импульсной активности в электронных аналогах нейронов. 1994. Депонирована в ВИНИТИ 17 февраля 1994. N 405-В стр., ил.7. Библ.28.
9. Романов мышцы: SU № 1 1985.
10. Романов для моделирования афферентного нейрона: SU № 1 1988.
11. Романов рецептора – мышечного веретена: SU № 1 1989.
12. Романов для моделирования нейронных структур двигательного аппарата: SU № 1 1990.
13. Эшби мозга. Происхождение адаптивного поведения. М.: ИЛ, 1962.
Теория нейронных сетей. II
