


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
, ,
Возможная общность макроскопических флуктуации скоростей биохимических и химических реакций, электрофоретической подвижности клеток и флуктуации при измерениях радиоактивности, оптической активности и фликкерных шумов
Oб авторе -
Oб авторе -
Oб авторе -
Oб авторе -
Oб авторе -
Oб авторе -
На протяжении 25-ти лет изучения феномена макроскопических флуктуации [1, 2] был совершен переход от представлений о том, что эти явления свойственны лишь белкам актомиозинового комплекса, к доказательству существования макроскопических флуктуации в самых различных биохимических и химических реакциях [3]. В последнее время было показано, то макроскопические флуктуации скоростей различных биохимических и химических реакций, по-видимому, обусловлены космофизическими причинами [4]. К аналогичному выводу пришел , на протяжении ряда лет исследующий дрейф (флуктуации) нулевых точек визуальных полутеневых поляриметров [5, 6].
В настоящем кратком сообщении показано, что явления, аналогичные макроскопическим флуктуациям скоростей биохимических и химических реакций, наблюдаются также при измерениях электрофоретической подвижности клеток, радиоактивности и фликкерных шумов. Показано также, что флуктуации, наблюдаемые в опытах и флуктуации в указанных выше процессах существенно сходны.
Для макроскопических флуктуации характерные дискретные распределения («статистические спектры») измеряемых величин [7]. Статистические спектры состояний, реализуемых в ходе макроскопических флуктуации в различных объектах претерпевают день за днем закономерную эволюцию форм. Статистические спектры макроскопических флуктуации различных объектов в синхронных опытах оказываются сходными по форме. Сходство формы статистических спектров является основным критерием общности изучаемых явлений.
На рис. 1 приведено сопоставление статистических спектров, полученных при измерениях макроскопических флуктуации в креатинкиназной реакции [3, 7] и при измерениях нулевой точки визуального поляриметра [5, 6]. Опыты с креатинкиназой проводили в нашей лаборатории и в Пущине, опыты с поляриметром в Москве. Видно, что в синхронных опытах наблюдается сходство макроскопических флуктуации разных по физической природе объектов, разделенных расстоянием порядка 100 км.
На рис. 2 приведено сопоставление статистических спектров флуктуации скоростей реакции аскорбиновой кислоты с дихлорфенолиндофенолом [3, 7] и флуктуации при измерениях радиоактивности препаратов 3Н, выполненных по нашей просьбе и в Институте биологической физики, посредством автоматического регистрирующего сцинтилляционного счетчика LS-30 фирмы Intertechnique в синхронных опытах. В каждом опыте проводилось по 100 измерений с интервалом в 1 мин скорости химической реакции (, ) или радиоактивности препарата 3Н (В. И.. Брусков, ). Как видно из рис. 2, форма статистических спектров - гистограмм распределения результатов сходна для двух совершенно разных по природе процессов в синхронных опытах. Результаты не зависят от средней активности измеряемого образца 3Н:имп/мин или имп/мин. Статистические спектры различны в разные дни.
|
Увеличить >>> |
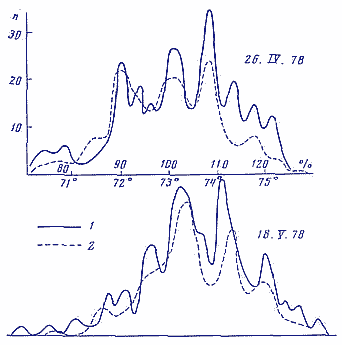
Рис. 1. Статистические спектры (гистограммы) флуктуации углов поворота лимба при измерении нулевой точки визуального поляриметра (1) и флуктуации скоростей креатинкиназной реакции (2). Синхронные опыты. Оси абсцисс - углы поворота лимба в градусах и ферментативная активность в % от средней арифметической величины. Оси ординат - число измерений, соответствующее данной величине абсциссы после двукратного сглаживания гистограмм.
|
Увеличить >>> |
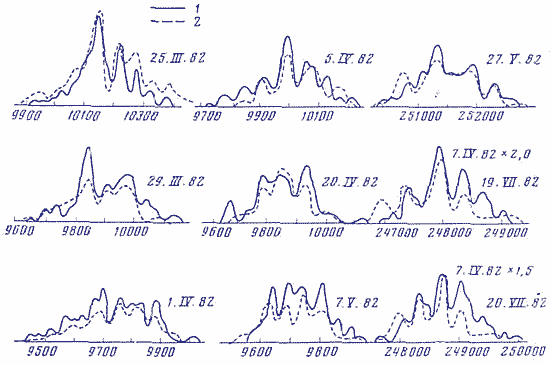
Рис. 2. Статспектры флуктуации скоростей реакции аскорбиновой кислоты (АК) с дихлорфенолиндофенолом (ДХФИФ) и флуктуации при измерениях радиоактивности препарата 3Н посредством жидкого сцинтиллятора. Синхронные опыты. Оси абсцисс - скорость реакции в % от средней арифметической величины и активность препарата в имп/мин. Оси ординат как на рис. 1.
|
Увеличить >>> |
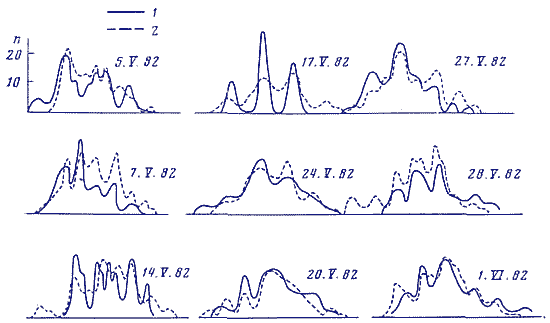
Рис. 3. Статспектры флуктуации времен ожидания разряда неоновой лампы в RC-генераторе, флуктуации скоростей реакции аскорбиновой кислоты с дихлорфенолиндофенолом, радиоактивности препаратов 3Н. Синхронные опыты. Оси абсцисс - длительность интервалов времени между вспышками неоновой лампы в % от средней арифметической величины и как на рис. 2. Оси ординат как на рис. 1.
|
Увеличить >>> |
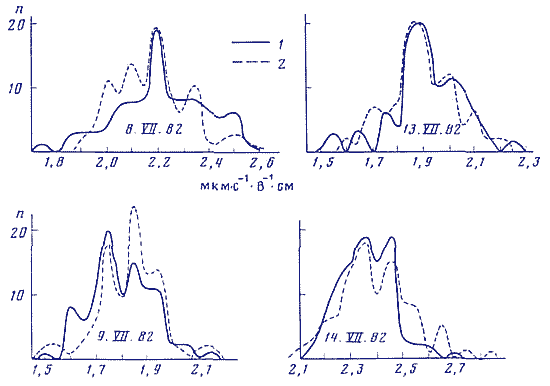
Рис. 4. Статспектры флуктуации электрофоретической подвижности (ЭФП) эритроцитов барана, фиксированных формальдегидом, и флуктуации скорости реакции аскорбиновой кислоты с дихлорфенолиндофенолом. Синхронные опыты. Оси абсцисс - ЭФП в мкм·с-1 ·В-1 ·см и как на рис. 2. Оси ординат как на рис. 1.
На рис. 3 сопоставлены статистические спектры макроскопических флуктуации в трех различных процессах в синхронных опытах при измерениях: скорости реакции аскорбиновой кислоты с дихлорфенолиндофенолом (, ), радиоактивности препарата 3Н (, ) и времени релаксации неонового генератора (, , ). Флуктуации времени релаксации, т. е. времени ожидания разряда неоновой лампы в RC-цепи - являются типичным примером фликкерных шумов (1/f процессов) [8]. Видное на рис. 3 сходство статистических спектров флуктуации всех трех процессов в синхронных опытах позволяет предположить, что фликкерные шумы и макроскопические флуктуации в изучаемых нами процессах могут быть следствием общих причин. В пользу такого заключения свидетельствует принадлежность макроскопических флуктуации к классу 1/f процессов [9].
На рис. 4 приведено сопоставление статистических спектров флуктуации электрофоретической подвижности эритроцитов барана, фиксированных формальдегидом (опыты ) и флуктуации скорости реакции аскорбиновой кислоты с дихлорфенолин-дофенолом - синхронные опыты и . Измерения электрофоретической подвижности клеток проводились на автоматическом регистрирующем приборе Пармоквант фирмы Цейсс, Йена, ГДР. Как и в других опытах проводили по 100 последовательных измерений с интервалом в 1 мин. При каждом измерении получали величину, среднюю для 7-10 клеток. Видно, что флуктуации электрических свойств поверхности фиксированных клеток характеризуются такими же статистическими спектрами, как и в синхронных опытах с химической модельной реакцией.
Таким образом, флуктуации при измерениях: радиоактивности, оптических, электрических свойств различных объектов., скоростей биохимических и химических реакций и фликкерных шумов сходны в синхронных опытах и различаются день за днем в каждом из процессов. Дискретные стат. спектры - дискретные величины скоростей химических и биохимических реакций, дискретные значения результатов измерений радиоактивного распада, дискретные флуктуации оптических и электрических свойств различных объектов, дискретные значения времен релаксации в генераторе фликкерных шумов - являются, по-видимому, проявлением весьма общих физических свойств.
Более подробное изложение имеющихся данных и обсуждение возможной физической природы описываемого явления - макроскопических флуктуации - предполагается осуществить в более пространной публикации.
Институт биологической физики
Биофизика, 1982, т. 28, вып.1, стр.153
ЛИТЕРАТУРА
1. Вопр. мед. химии, 1958, т. 4, с. 443.
2. В кн.: Колебательные процессы в биологических и химических системах. М, Наука, 1967, с. 22.
3. , , Б рицина Т. Я. Биофизика, 1980, т. 25, с. 409.
4. , , 1-й Всесоюзный биофизический съезд, Тез. докл., плен. лекц. и симпоз. засед. Пущине: Изд. ЦБИ АН СССР, 1982, с. 119.
5. Изв. АН СССР, сер. - биол., 1982, № 3, с. 467.
6. В кн. Проблемы космической биологии. М.: Наука, 1982, т. 43, с. 197-211.
7. , , Биофизика,, 1980, т. 25, с. 213.
8. Тр. ин-та инжен. электротехн. и радиоэлектрон. М: Мир, 1982, т. 70, №2, с. 212.
9. Биофизика, 1982, т. 27, с. 529.
Институт биологической физики
Биофизика, 1982, т. 28, вып.1, стр.153
, , Морозова общность макроскопических флуктуации скоростей биохимических и химических реакций, электрофоретической подвижности клеток и флуктуации при измерениях радиоактивности, оптической активности и фликкерных шумов // «Академия Тринитаризма», М., Эл , публ.11466, 01.09.2004
