


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Г. Д. ХОРУНЖИЙ, М. А. ЕГОРОВА
Институт эволюционной физиологии и биохимии им. РАН,
Санкт-Петербург
*****@***com
СВОЙСТВА ЧАСТОТНЫХ РЕЦЕПТИВНЫХ ПОЛЕЙ
ОДИНОЧНЫХ НЕЙРОНОВ ПЕРВИЧНОЙ СЛУХОВОЙ
КОРЫ МЫШИ КАК ОСНОВА КОРТИКАЛЬНОЙ
ОБРАБОТКИ АКУСТИЧЕСКОЙ ИНФОРМАЦИИ[1]
В двух тонотопически организованных полях слуховой коры мыши (AI и AAF) исследованы фундаментальные свойства частотных рецептивных полей одиночных нейронов: форма области возбудительного ответа, острота частотной настройки, характеристические частоты, пороги ответов нейронов. Обсуждаются различия частотно-временных характеристик активности нейронов первичной слуховой коры и стволовых центров слуха и их взаимосвязь с особенностями кодирования звуков на разных уровнях слуховой системы.
Ключевые слова: слуховая кора, одиночные нейроны, частотные рецептивные поля
Введение
Создание целостного представления о кодировании параметров акустических сигналов в слуховой системе подразумевает знание частотного и временного механизмов обработки сигналов центральными слуховыми нейронами. Очевидно, что формирование полной картины реализации этих механизмов невозможно в отсутствие сведений о фундаментальных свойствах частотных рецептивных полей слуховых нейронов. Особый интерес вызывают вопросы, связанные с нейрональными механизмами кодирования акустической информации на уровне слуховой коры как высшего центра слуховой системы млекопитающих. Несмотря на обилие работ, посвященных исследованию слуховой области коры млекопитающих [1-7], сведения об особенностях импульсной активности образующих ее нейронов остаются неполными. Хотя домовая мышь, являясь акустически специализированным объектом, активно используется как модель для изучения механизмов слуха млекопитающих, наиболее достоверная информация о ее слуховой коре связана с особенностями анатомического строения и цитоархитектоники этой структуры. Так, известно, что слуховая кора мыши представляет собой сложное образование, включающее, как минимум, пять полей – три первичных и два вторичных поля. К первичным полям слуховой коры мыши относят первичное слуховое поле (AI), переднее слуховое поле (AAF) и ультразвуковое поле (UF), к вторичным – вторичное слуховое поле (AII), а также дорсопостериальное поле (DP) [2, 4–7]. Данные же об основных характеристиках активности нейронов слуховой коры мыши связаны, главным образом, с особенностями топической организации частот в полях коры [5]. Информация о свойствах частотных рецептивных полей одиночных нейронов слуховой коры мыши и о параметрах их импульсной активности представлена лишь в небольшом числе публикаций [5, 7]. Нами было начато исследование основных характеристик рецептивных полей и импульсной активности одиночных нейронов первичной и передней слуховой коры (поля AI и AAF) [2], продолжением которого является данная работа. Задача настоящего этапа исследования – систематическое описание частотных рецептивных полей одиночных нейронов первичного и переднего слуховых полей, оценка взаимосвязей между их свойствами, а также сравнение рецептивных полей нейронов слуховой коры с нейрональными рецептивными полями стволовых центров слуховой системы, в первую очередь, центрального ядра заднего холма как основного стволового интегративного центра слуха [8, 9].
Материалы и методы
В работе внеклеточно регистрировали импульсную активность одиночных нейронов первичного слухового поля (AI) и переднего слухового поля (AAF) слуховой коры у 24 наркотизированных домовых мышей (Mus musculus), самок в возрасте 8–16 недель, 12 из которых были гибридами линий CBA и C57/BL6, и еще 12 – гибридами линии NMRI с дикими животными. Регистрация осуществлялась путем погружения в мозг животного вольфрамового микроэлектрода сопротивлением 4–7 мОм. Электроды вводились ортогонально поверхности мозга в каудальную часть височной коры левого полушария (поле 41), соответствующую расположению слуховой коры животного [4]. Ввиду выраженности у мышей индивидуальных топографических различий в локализации полей слуховой коры для определения их границ у каждого животного производили структурно-функциональное картирование слуховой коры с шагом 500 мкм в ростро-каудальном направлении и 300 мкм в дорсо-вентральном направлении. Основным критерием при локализации полей АI и AAF служило наличие тонотопической организации с градиентом частот по ростро-каудальной оси, показанное в работе Stiebler с соавт [4]. В первичном слуховом поле характеристические частоты нейронов убывали в ростро-каудальном направлении, в переднем слуховом поле порядок частот инвертировался, т. е. они возрастали в ростро-каудальном направлении. Регистрацию производили на глубине 300-600 мкм, соответствовавшей расположению IV–V слоев слуховой коры животного.
Анестезию поддерживали внутрибрюшинными инъекциями смеси кетамина (кетавет, 35 мг/кг) и ксилазина (ромпун, 0,1 мг/кг), что обеспечивало стабильный неглубокий наркоз животного с минимальной активацией тормозных воздействий на центральные слуховые нейроны [9, 10].
Нейрональные ответы усиливали в 10000 раз, отфильтровывали (0,3-10 кГц, усилитель DAM80) и выводили параллельно на четырехканальный осциллограф (Tektronix 5113), громкоговоритель и оконный дискриминатор (WPI, 120) для дальнейшей регистрации в форме стандартных импульсов и введения в XT-совместимый компьютер для «on-line» и «off-line» анализа. Генерация сигналов, регистрация спайковой активности и обработка полученных результатов обеспечивались возможностями специализированного программного обеспечения (ТМS97), разработанного Dr. Schulze-Krueger (Германия) на базе системной платы TMS 320 C30 AD/DA, встроенной в РС-486 (тактовая частота 200 кГц, разрешение по амплитуде 16 бит). Стимулы подавались посредством электродинамического звукоизлучателя (Fostex FT-17 H, неравномерность частотной характеристики – ± 6 дБ на частотах 5–49 кГц). Частотные рецептивные поля нейронов тестировали одиночными тонами и двухтоновыми комплексами. Тональные сигналы длительностью 100 мс, временем нарастания и спада 5 мс предъявляли с интервалом 900 мс в случайном порядке комбинаций 45 фиксированных частот и 15 интенсивностей (всего 675 различных тонов). В двухтоновой парадигме один тон соответствовал характеристической частоте нейрона уровнем 10 дБ над порогом ответа, другой начинался на 5 мс раньше и изменялся в широком диапазоне частот и интенсивностей, попадающих как в область возбудительной, так и тормозной реакции нейрона. Шаг по интенсивности составлял 5 дБ, по частоте – 1/45 от выбранного частотного диапазона по логарифмической шкале.
Анализ импульсной активности и свойств рецептивных полей выполнен для 80 нейронов поля AI и 80 нейронов поля AAF.
Результаты
Все исследованные нейроны отличались фазным (1 – 5 спайков в ответе на сигнал) или пачечным (до 10 спайков в пачке) типом временного узора разряда.
Анализ формы области возбудительного ответа зарегистрированных нейронов выявил их широкую частотную настройку как в первичном, так и в переднем слуховом поле (рис. 1).
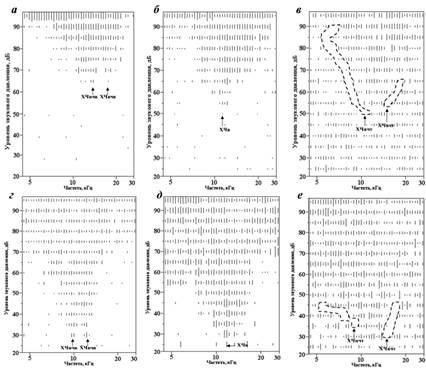
Рис. 1. Диаграммы частотной области ответа типичных нейронов первичного
слухового поля (а – в) и переднего слухового поля (г – е), демонстрирующие
особенности мультипиковых (а, г) и V-образных (б, д) частотных возбудительных рецептивных полей
На рис. 1 для каждого V-образного нейрона представлена диаграмма его тормозных зон, полученная методом двухтонового торможения (в, е, тормозные зоны выделены пунктирными линиями). Указаны стрелками: ХЧнчв – низкочастотная характеристическая частота возбуждения, ХЧвчв – высокочастотная характеристическая частота возбуждения (у мультипиковых нейронов), ХЧв – характеристическая частота возбуждения, ХЧнчт – низкочастотная характеристическая частота торможения, ХЧвчт – высокочастотная характеристическая частота торможения (у V-образных нейронов). Минимальная высота черты соответствует одному спайку.
Значения коэффициента Q10, характеризующего остроту (добротность) частотной настройки нейрона, составляли 0,5 – 3,7 для нейронов поля AI и 0,4 – 4,5 для единиц поля AAF (рис. 2, а, б). Коэффициент Q10 вычислялся как отношение характеристической частоты возбуждения нейрона к ширине частотной области ответа нейрона, вызванного действием стимула уровнем 10 дБ над порогом ответа. Почти 64 % нейронов имели значения добротности, лежащие в диапазоне 0,9–1,9. Значения Q10 нейронов полей AI и AAF достоверно не различались (p > 0,05, тест Манна-Уитни). Ни для нейронов поля AI, ни для единиц поля AAF регрессионный анализ не показал зависимости добротности от характеристических частот зарегистрированных нейронов (p > 0,05). Около трети исследованных нейронов поля AI (31 %) и 21 % единиц поля AAF имели частотные рецептивные поля сложной мультипиковой формы и не менее двух характеристических частот возбуждения (низко - и высокочастотной) (рис. 1, а, г). Остальные нейроны имели V-образные частотные рецептивные поля, характеризующиеся симметричными, широкими низкочастотными и высокочастотными зонами возбудительного ответа (рис. 1, б, д). Частоты встречаемости нейронов с разными типами рецептивных полей в слуховой коре и в центральном ядре заднего холма [9] достоверно различались (χ2-тест, р < 0,001).
Характеристические частоты исследованных нейронов, определенные аудиовизуально, занимали диапазон от 8 до 34 кГц в первичном слуховом поле и 7–30 кГц в поле AAF. Как для поля AI, так и для переднего слухового поля было отмечено преобладание единиц, с характеристическими частотами, лежащими в диапазоне 10–25 кГц (70 % нейронов в поле AI и 78 % единиц в поле AAF).
Минимальный пороговый уровень ответа нейрона на сигнал характеристической частоты составлял -10 дБ УЗД в поле AI и 0 дБ УЗД в поле AAF, максимальный – 60 дБ УЗД как в первичном, так и в переднем слуховом поле. Регрессионный анализ зависимости пороговых уровней ответов нейронов от их характеристических частот выявил взаимосвязь этих параметров с высокой степенью достоверности (рис. 2, в, г) (полиномиальная регрессия, p < 0,01). Наименьшие пороги ответов нейронов и в поле AI, и в поле AAF наблюдались в диапазоне частот 15 – 25 кГц.
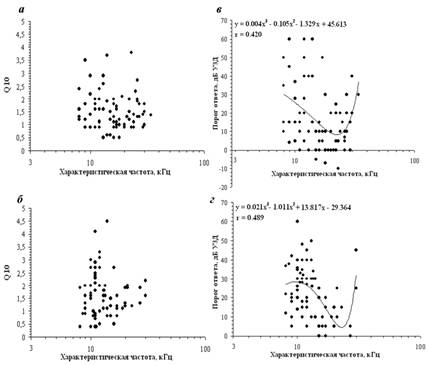
Рис. 2. В левой части рисунка – распределение значений Q10 в зависимости
от ХЧв нейронов полей AI (а) и AAF(б); в правой части рисунка – распределение
значений порогов ответов нейронов полей AI (в) и AAF (г) в зависимости от их ХЧв. Жирные линии на графиках – линии полиномиальной регрессии (р < 0,01), соответствующие приведенным на графиках уравнениям регрессии
Следует отметить, что точное определение характеристических частот и порогов ответов нейронов было затруднено ввиду выраженной спонтанной активности и широкой частотной настройки нейронов слуховой коры. Выраженность спонтанной активности была отмечена в ранее выполненных исследованиях как первичных, так и вторичных слуховых полей [2, 4] и подтверждена в данной работе – спонтанная активность была обнаружена у подавляющего большинства нейронов поля AI (98 %) и у всех нейронов поля AAF.
Исследованные нейроны, как правило, характеризовались наличием боковых тормозных зон, симметрично ограничивающих центральную область возбудительного ответа с низко - и высокочастотной сторон (см. рис. 1, в, е). Их наличие также подтверждалось подавлением спонтанной активности по периферии возбудительного рецептивного поля в однотоновой парадигме стимуляции. Однако в силу нерегулярного ответа нейронов слуховой коры в двухтоновой парадигме и их высокой спонтанной активности, точное определение тормозных характеристических частот и, тем более, границ областей торможения было сильно затруднено. Как правило, тормозные зоны исследованных нейронов имели вид узких областей, которые могли быть обнаружены только по сниженной нейрональной активности вдоль границы возбудительного рецептивного поля нейрона (см. рис. 1, в, е), точно определить их протяженность не представлялось возможным.
Обсуждение
Отсутствие каких-либо различий по свойствам частотных рецептивных полей и характеристикам импульсной активности между нейронами полей AI и AAF очевидно указывает на общие источники восходящих проекций к первичному и переднему слуховым полям от стволовых центров слуховой системы и, возможно, на схожий характер реализации стратегий кодирования акустической информации нейронами этих полей.
Показано, что на уровне центрального ядра заднего холма значительная часть нейрональной популяции в условиях поверхностной анестезии, не оказывающей серьезного влияния на тормозные процессы, демонстрирует узкую частотную настройку (вплоть до формирования зон возбудительного ответа закрытого типа в рецептивных полях нейронов) и тонические характеристики активности [8, 9]. Следовательно, актуальным становится вопрос о механизмах, лежащих в основе формирования принципиально иных свойств нейронов слуховой коры. Поскольку результаты настоящей работы согласуются с ранее полученными данными о низкой частотной избирательности нейронов слуховой коры [2, 5], то обоснованным выглядит предположение, что в слуховой коре происходит изменение принципов кодирования акустической информации, поступающей от нижележащих центров слуховой системы; возможно, спектральный механизм обработки стимула становится менее значимым на уровне слуховой коры, чем на уровне стволовых центров слуховой системы.
Таким образом, полученные данные о форме области возбудительного ответа, характеристических частотах, порогах ответов на сигнал характеристической частоты, типе временного узора разряда, а также о соотношении тормозных и возбуждающих характеристических частот в частотных рецептивных полях исследованных нейронов позволили установить, во-первых, что нейроны первичного и переднего слуховых полей не различаются ни в одном из этих аспектов, а также подтвердить принципиальные отличия свойств нейронов первичной слуховой коры от единиц нижележащих центров слуховой системы, например, центрального ядра заднего холма [3, 8, 9]. В то же время, вопрос о функциональной специализации первичного и переднего слуховых полей, во многих отношениях дублирующих друг друга, требует дальнейшего изучения.
Список литературы
1. Вартанян анализ сложных звуков. Л.: Наука, 1978.
2. Егорова настройка нейронов первичного слухового поля (АI) и переднего слухового поля (ААF) слуховой коры мыши (Mus Musculus) // Журн. Эвол. Биохим. и физиол. 2005. Т.41. № 4. С. 379-382.
3. Aitkin L. M., Irvine D. R.F., Nelson J. E., Merzenich M. M., Clarey J. C. Frequency representation in the auditory midbrain and forebrain of a marsupial, the northern native cat // Brain Behav. Evol. 1986. V. 29. P. 17-28.
4. Stiebler I., Neulist R., Fichtel I., Ehret G. The auditory cortex of the house mouse: left-right differences, tonotopic organization and quantitative analysis of frequency representation // p. Physiol. A. 1997. V. 181. P. 559-571.
5. Linden J. F., Liu R. C., Sahani M., Schreiner C. E. Spectrotemporal Structure of Receptive Fields in Areas AI and AAF of Mouse Auditory Cortex // J Neurophysiol. 2003. V. 90. P. 2660–2675.
6. Imaizumi K., Lee C. C., Linden J. F., Winer J. A., Schreiner C. E. The anterior field of auditory cortex: neurophysiological and neuroanatomical organization // The Auditory Cortex. Eds. König R., Heil P., Budinger E., Scheich H., Mahwah, London: Lawrence Erlbaum Associates Inc. 2005. P. 95–110.
7. Shen J., Xu Z., Yao Y. Topography of acoustic response characteristics in the auditory cortex of the Kunming mouse // Chinese Science Bulletin. 2000. V. 45. P. 443-448.
8. Егорова свойства импульсной активности одиночных нейронов задних холмов мыши // Сенсорные системы. 2008. Т. 22. №3. С. 203-213.
9. Egorova M., Ehret G., Vartanian I., Esser K.-H. Frequency response areas of neurons in the mouse inferior colliculus. I. Threshold and tuning characteristics // Exp. Brain Research. 2001. V.140. P. 145-161.
10. Zhou R., Assouline J. G., Abbas P. J., Messing A., Gantz B. J. Anatomical and physiological measures of auditory system in mice with peripheral myelin deficiency // Hear. Res. 1995. V. 88. P. 87–97.
1Работа выполнена при финансовой поддержке РФФИ (грант № ).
