
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Л. Н. ВАСИЛЬЕВА
Институт высшей нервной деятельности и нейрофизиологии РАН, Москва
Московский государственный университет им.
*****@***ru
СТАБИЛЬНОСТЬ РЕГИСТРАЦИИ АКТИВНОСТИ
ОДИНОЧНЫХ НЕЙРОНОВ И РАЗРАБОТКА КРИТЕРИЕВ ЕЕ ОЦЕНКИ
Настоящее исследование направлено на решение проблемы стабильности регистрации нейронной активности; оценивали качество регистрации одиночных нервных клеток с помощью пучка микропроволочек и микроэлектродов, а также работали над созданием критериев оценки стабильности регистрации по форме спайков. Несмотря на сниженное качество регистрации по сравнению с микроэлектродами, благодаря своим биомеханическим свойствам микропроволочки могли бы стать решением проблемы стабильности регистрации при создании нейропротезов.
Ключевые слова: регистрация нейронной активности, инвазивные нейропротезы, интерфейс мозг-компьютер, сортировка спайков, критерии стабильности регистрации
Введение
Под нейропротезами понимают устройства, устанавливающие связи между мозгом и внешним миром путем регистрации его активности или стимуляции определенных структур [Hatsopoulos, Donoghue, 2009]. Ожидается, что нейропротезы расширят возможности человеческого тела: они бы позволили людям управлять объектами внешнего мира, сбылась бы мечта человечества об управлении предметами на расстоянии. По этой же причине создание нейропротезов актуально для клинической практики, причем были бы востребованы и сенсорные, и моторные нейропротезы.
Назначение моторных нейропротезов состоит в восстановлении утраченных или ослабленных двигательных функций. Их применение позволило бы увеличить качество жизни людей с различными двигательными расстройствами: параличами, травмами спинного мозга, нейродегенеративными заболеваниями, а также людей с отнятыми конечностями и т. д.
Моторные нейропротезы состоят из нескольких функциональных блоков. Регистрирующий блок получает сигнал от головного мозга с поверхности скальпа или непосредственно от нервной ткани. Он соединен с блоком, осуществляющим математическую обработку полученного от мозга сигнала и вычисляющим двигательную программу. Третий – исполнительный блок, который управляет искусственной конечностью или электрически стимулирует мышцы пациента.
Моторные нейропротезы можно разделить на две группы: неинвазивные и инвазивные. Первые управляются сигналом электрo-энцефалограммы, полученным со скальпа испытуемого. Инвазивные нейропротезы управляются сигналом, отводимым непосредственно от нервной ткани, то есть требуют операционного вмешательства для имплантации регистрирующих электродов.
За последние годы нейропротезирование достигло больших успехов. В опытах на животных было показано, что с помощью сигнала корковых нейронов можно управлять искусственными эффекторами [Chapin et al., 1999; Velliste et al., 2004]. В настоящее время управление нейропротезами производится с участием различных корковых зон, вовлеченных в программирование и реализацию движения, среди которых первичная моторная кора, заднетеменная кора, дополнительная моторная область, дорзальная премоторная кора, первичная сенсомоторная кора [Hatsopoulos, Donoghue, 2009].
Тем не менее, перед нейропротезированием стоит несколько вопросов, которые необходимо решить, прежде чем мы сможем использовать нейропротезы в клинической практике [Lebedev, Nicolelis, 2006]. Одним из таких вопросов является решение проблемы биосовместимости регистрирующих электродов. При погружении электродов в нервную ткань довольно быстро развивается глиальная реакция в месте нахождения электрода. Астроциты и микроглиоциты скапливаются вокруг электрода и изолируют нервную ткань от чужеродного объекта [Marin, Fernandez, 2010]. Это ведет к снижению качества регистрируемого сигнала на данном электроде со временем, что ограничивает срок службы нейропротезов до нескольких недель или месяцев. Увеличение срока службы нейропротезов путем решения проблемы биосовместимости регистрирующих устройств позволило бы продвинуться на пути к внедрению нейропротезов в клиническую практику.
Тонкие нихромовые микропроволочки можно рассматривать как возможное решение проблемы долговечности нейропротезов. Известно, что с помощью таких микропроволочек регистрация может проводиться в течение 7-ми лет [Krüger et al., 2010]. Из-за своих биомеханических свойств микропроволочки меньше повреждают ткань при сдвигах, так как смещаются вместе с ней, в отличие от микроэлектродов. По этой же причине микропроволочки могут стать решением проблемы стабильности регистрации в нейропротезировании.
Известно, что в силу различных причин электрод может сдвигаться в мозговой ткани, а его кончик может смещаться из области, в которой проводится регистрация. То есть в разные периоды регистрации может быть записан сигнал разных нейронов. Для создания нейропротезов, а также в ряде нейрофизиологических исследований необходима регистрация одних и тех же нейронов на протяжении долгого времени.
Для решения проблемы стабильности регистрации используют методы сортировки спайков, так как требуется ответ на вопрос, проводится ли регистрация активности одного и того же нейрона или различных нейронов в разные периоды времени. Методы сортировки спайков достаточно разнообразны, причем каждый метод обладает своими преимуществами и ограничениями [Lewicki, 1998]. Самый простой способ разделения спайков – классификация на основании их параметров. Форму спайка описывают с помощью нескольких параметров: длительности, амплитуды и т. п. В таком случае, чем больше параметров мы будем рассматривать, тем эффективнее классифицируем спайки. Достаточно распространено разделение ПД нейронов с помощью метода главных компонент. С помощью данного метода форма нейрона описывается как линейная комбинация набора кривых – главных компонент. Главные компоненты назначаются по убыванию того, какую часть изменчивости данных они описывают, причем каждая следующая перпендикулярна предыдущей в многомерном пространстве. Добавление новых главных компонент позволяет увеличить качество описания данных, однако вклад каждой новой компоненты ниже, чем у предыдущей. Таким образом, можно задействовать новые и новые главные компоненты, пока спайк не будет описан полностью. Существуют и другие методы сортировки спайков, например, с использованием кластерного анализа или вейвлет-преобразования.
Ранее был предложен метод оценки стабильности регистрации нейронной активности, основанный на анализе частоты разряда нейрона и ее модуляций в постстимульный период [Бондарь, 2011]. Кроме того, для повышения стабильности регистрации нейронной активности используют автоматизированные методы. В случаях, когда регистрация проводится с помощью множества электродов, у экспериментатора нет возможности подстраивать положение каждого электрода для получения оптимального сигнала на нем. В этих случаях используют алгоритмы, которые изменяют положение отдельных электродов в матрице для получения оптимального качества сигнала [Wolf et al., 2009, Chakrabarti et al., 2012].
Классификация спайков, а также присвоение их разным нейронам позволило бы выяснить, проводится ли регистрация в разные дни от одних и тех же нейронов или различных.
Цель исследования
Настоящее исследование направлено на оптимизацию регистрации активности одиночных нервных клеток с помощью пучка множественных микроэлектродов и выработку критериев оценки стабильности регистрации по формам потенциалов действия.
Задачи
· В экспериментах на анестезированных животных осуществить регистрацию нейронной активности с помощью заостренного электрода и пучка множественных микропроволочек.
· Оптимизировать процесс начального выделения ПД нейронов из нативной записи и провести дополнительную кластеризацию данных с помощью метода главных компонент.
· Сравнить качество регистрации нейронной активности с помощью заостренного микроэлектрода и пучка.
· Разработать критерий оценки стабильности регистрации с использованием корреляционного анализа, примененного к усредненным формам ПД одного и того же, а также разных нейронов.
Материалы и методы
Регистрацию нейронной активности проводили на анестезированных крысах и кошках, регистрацию нейрональной активности обезьян проводили на бодрствующих животных. Нейронную активность регистрировали с помощью игольчатых электродов (импеданс 2–4 МОм) и самостоятельно изготовленных пучков микропроволочек (диаметр 12 и 18 мкм). Кусочки проволоки длиной 10 см припаивали к стандартным матрицам для крепления микросхем. Пучок продевали в полиимидную трубочку (внешний диаметр 200 мкм). Помещали в металлическую направляющую канюлю (внешний диаметр 600 мкм). Готовый пучок прикрепляли к съемному микроманипулятору, который позволял двигать пучок относительно направляющей канюли.
На анестезированных крысах и кошках проводили операцию по трепанации черепа, во время которой освобождали доступ к коре. После окончания операции начинали погружение электрода. Поверхность мозга преодолевали с помощью канюли, заливали поверхность мозга раствором агарозы, а дальнейшее погружение микропроволочек в мозг осуществляли с помощью съемного микроманипулятора, которым был снабжен пучок. У обезьян вживление пучка микропроволочек осуществляли через трепанационное отверстие диаметром 5 мм.
Усиление составляло 25000 раз в диапазоне частот 300 Гц – 10 кГц. Частота опроса составляла 16,5–25 кГц. Запись электрической активности нейронов, а также начальная обработка данных осуществлялись с помощью программы Spike2. Начальная обработка данных включала в себя детектирование ПД по порогу амплитуды. Участки записи, содержащие ПД, были выделены и импортированы в формат программы Matlab. Дальнейшая обработка данных осуществлялась с помощью программ, созданных в среде Matlab. Спайки классифицировали с помощью метода главных компонент, а также рассчитывали коэффициенты корреляции внутри классов ПД (усредняя по 20 спайков) и между классами (усредняя все спайки внутри класса).
Результаты
Оценка физических свойств микропроволочек и микроэлектродов показала, что импеданс микропроволочек ниже импеданса микроэлектродов. Среднее значение импеданса микропроволочек толщиной 18 мкм составило 1,07 ± 0,17 МОм (параметры тестирования: частота – 1 кГц, сила тока 100 нА; 86 микропроволочек). На микропроволочках толщиной 12 мкм было получено среднее значение импеданса 1,41 ± 0,19 МОм (параметры тестирования те же, 27 микропроволочек). Импеданс игольчатого микроэлектрода – 3,4 МОм.
В ходе трех экспериментов на крысах было зарегистрировано 37 нейронов с помощью микропроволочек толщиной 18 мкм и 17 – с помощью микропроволочек толщиной 12 мкм. Средняя амплитуда ПД, полученных с помощью проволоки 18 мкм, составила 89,03 ± 39,46 мкВ, а величина фонового шума 26,08 ± 4,66 мкВ. Таким образом, в среднем отношение сигнала к шуму для микропроволочек толщиной 18 мкм составило 3,63 ± 2,2. Для микропроволочек диаметром 12 мкм средняя амплитуда ПД составляет 108,78 ± 58,73 мкВ, а величина фонового шума – 30,42 ± 16,08 мкВ.
Физические свойства электродов влияют на качество записи. На рис. 1 отображены фрагменты нативной записи, полученной с помощью различных электродов. Запись наглядно иллюстрирует, что при регистрации с помощью игольчатого микроэлектрода отношение сигнал–шум заметно выше, чем при регистрации с помощью нихромовых микропроволочек. Надо отметить, что иногда при регистрации с помощью микропроволочек толщиной 12 мкм достигается отношение сигнал–шум, сравнимое с таковым для игольчатого электрода, хотя в большинстве случаев оно все-таки ниже.
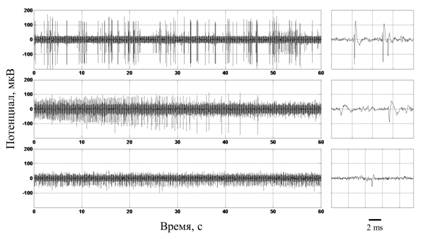
Рис. 1. Фрагменты нативной записи, полученной с помощью игольчатого
микроэлектрода (верхний ряд) и пучка микропроволочек (два нижних ряда).
В левом столбце представлены фрагменты записи длиной 1 мин, в правом – 10 мс
Кроме того, при регистрации с помощью игольчатого электрода отчетливо выделяются потенциалы действия нейронов разной амплитуды. При регистрации с помощью проволочек также можно выделить два класса ПД, различающихся по амплитуде, но ПД меньшей амплитуды оказывается сильно зашумленным.
К классам ПД, выделенным с помощью метода главных компонент, был применен корреляционный анализ. Преимущество корреляционного анализа состоит в том, что с его помощью можно оценить степень схожести ПД нейронов, не прибегая к нормализации данных по амплитуде. Был произведен расчет коэффициентов корреляции между усредненными формами ПД каждого класса. Внутри класса коэффициент корреляции был рассчитан между ПД, усредненными по 20.
На основании полученных значений коэффициентов корреляции были построены гистограммы распределения коэффициентов корреляции отдельно для корреляционного анализа между классами ПД и внутри одного класса.
Для разработки критерия стабильности регистрации были проанализированы коэффициенты корреляции, полученные для ПД крыс и кошек в остром опыте (рис. 2). По мнению автора, назначение некоторого критического значения коэффициента корреляции является целью разработки критерия стабильности регистрации ПД нейронов. Найденное критическое значение коэффициента корреляции должно быть таким, чтобы вероятности ошибочного отнесения двух ПД одному или двум классам были оптимальны.
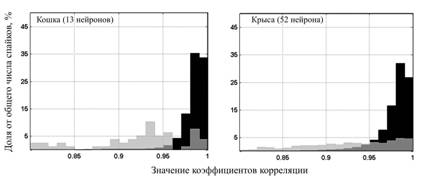
Рис. 2. Корреляционный анализ ПД нейронов внутри одного класса (черный
график) и между классами (серый график). Верхний график: гистограммы
распределения коэффициентов корреляции ПД кошек (13 нейронов, игольчатый электрод). Нижний график: гистограммы распределения коэффициентов
корреляции ПД крыс (52 нейронов, пучок микропроволочек)
При значении коэффициента корреляции 0.96 оказывается, что и в случае кошек (игольчатый микроэлектрод) и в случае крыс (микропроволочки) не более 84 % заведомо схожих ПД имеют коэффициент корреляции выше 0,96. Кроме того, до 16 % заведомо различных ПД имеют коэффициент корреляции выше критического значения. То есть при назначении 0,96 критическим значением коэффициента корреляции будет довольно высоко количество ошибок 1-го рода (ПД, являющиеся на самом деле различными, присвоены одному классу) и ошибок 2-го рода (ПД, принадлежащие одному классу, назначены различными).
По результатам наших опытов вне зависимости от типа использованных электродов при классификации ПД по значению коэффициента корреляции ошибки 1-го и 2-го рода происходят с вероятностью 12–16 % и 83–84 % соответственно.
Для дальнейшей разработки нашего критерия были использованы записи экстраклеточной нейронной активности обезьян, предоставленные . Данные были получены путем регистрации активности нейронов нижневисочной коры обезьян с помощью пучка микропроволочек.
К данным, зарегистрированным на обезьянах, также был применен корреляционный анализ, результаты которого отображены на диаграмме (рис. 3). Приведенные на рисунке гистограммы были использованы для проверки критерия оценки стабильности регистрации. Оказалось, что 86-88 % ПД, заведомо принадлежащих одному классу, имеют коэффициент корреляции больше 0,96, что сравнимо с аналогичной долей ПД, полученной у кошек и крыс. Тем не менее, доля ПД, заведомо принадлежащих разным классам и имеющих коэффициент корреляции более 0,96, составляет 24 %, что существенно выше аналогичной доли у крыс и кошек.
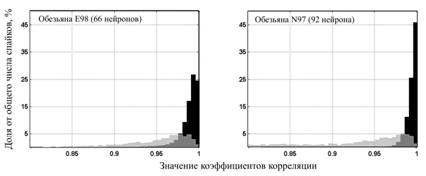
Рис. 3. Корреляционный анализ ПД нейронов обезьян внутри одного класса
(черный график) и между классами (серый график). А. Гистограммы распределения коэффициентов корреляции ПД обезьяны E98 (66 нейронов). Б. Гистограммы распределения коэффициентов корреляции ПД обезьяны N97 (92 нейронов)
Высокая доля ПД, принадлежащих различным классам и дающим высокий коэффициент корреляции, может быть объяснена наличием однообразных артефактов среди выделенных классов ПД, которые не были отвергнуты. При усреднении такие артефакты могут влиять на форму ПД и, следовательно, влиять на результат корреляционного анализа. При расчете коэффициентов корреляции ПД, заведомо принадлежащих одному классу, наличие внутри класса артефактов может приводить к занижению коэффициентов корреляции.
Необходимо помнить, что количество нейронов, зарегистрированное у крыс и кошек (52 и 13 нейронов, соответственно) было ниже, чем у обезьян E98 и N97 (66 и 92 нейрона, соответственно). По этой причине наша первичная оценка критического значения коэффициента корреляции могла оказаться не совсем точной.
Таким образом, дальнейшее совершенствование данного критерия может быть направлено на более эффективное выделение спайков и исключение из рассмотрения артефактов записи, что позволит увеличить стабильность регистрации и качество ее оценки.
Заключение
1. Микропроволочки позволяют получать записи приемлемого качества с хорошо заметными спайками на них и поэтому могут быть использованы в конструировании инвазивных нейропротезов, хотя и обеспечивают более низкое качество регистрации, чем микроэлектроды.
2. В данной работе была предпринята попытка разработки критерия стабильности регистрации нейронной активности. Анализ значений коэффициентов корреляции позволил нам выдвинуть предположение о критическом значении коэффициента корреляции, равном 0,96.
3. Анализ распределений коэффициентов корреляции между формами спайков показал, что выдвинутый критерий не позволяет судить о принадлежности спайков одному классу или различным с достаточной точностью и поэтому требует дальнейшей доработки.
Список литературы
1. Hatsopoulos N. G., Donoghue J. P. The science of neural interface systems // Annual Reviews Neuroscience. 2009. № 32. P. 249-266.
2. Chapin J. K., Moxon K. A., Markowitz R. S., Nicolelis M. A. Real-time control of a robot arm using simultaneously recorded neurons in the motor cortex // Nature Neuroscience. 1999. V. 2. № 7. P. 664-670.
3. Velliste M., Perel S., Spalding M. C., Whitford A. S., Schwartz A. B. Cortical control of a prosthetic arm for self-feeding // Nature. 2008. V. 453. № 000. P. .
4. Lebedev M. A., Nicolelis M. A. Brain-machine interfaces: past, present and future // Trends in Neurosciences. 2006. V. 29. № 9. P. 536-546.
5. Marin C., Fernández E. Biocompatibility of intracortical microelectrodes: current status and future prospects // Frontiers in Neuroengineering. 2010. V. 28. № 3. P. 8.
6. Krüger J., Caruana F., Volta R. D., Rizzolatti G. Seven years of recording from monkey cortex with a chronically implanted multiple microelectrode // Frontiers in Neuroengineering. 2010. V. 28. № 3. P. 6.
7. Lewicki M. S. A review of methods for spike sorting: the detection and classification of neural action potentials // Network. 1998. V. 9. № 4. P. 53-78.
8. Бондарь признаков изображения и сложных зрительных образов нейронами коры головного мозга млекопитающих. Дис. д-ра биол. наук. М., 2011.
9. Wolf M. T., Cham J. G., Branchaud E. A., Mulliken G. H., Burdick J. W., Andersen R. A. A robotic neural interface for autonomous positioning of extracellular recording electrodes // The International Journal of Robotics Research. 2009. V. 28. № 9. P. .
10.Chakrabarti S., Hebert P., Wolf M. T., Campos M., Burdick J. W., Gail A. Expert-like performance of an autonomous spike tracking algorithm in isolating and maintaining single units in the macaque cortex // Journal of Neuroscience Methods. 2012. V.205. № 1. P. 72-85.
М. В. СКОРНЯКОВА1,², А. Г. СУХОВ², В. Н. КИРОЙ²
¹Научно-исследовательский институт системных исследований РАН, Москва
²НИИ нейрокибернетики им. Южного федерального университета, Ростов-на-Дону
*****@***ru
ИССЛЕДОВАНИЕ И МОДЕЛИРОВАНИЕ
АЛЬФА-ЧАСТОТНОЙ ВЕРЕТЕНООБРАЗНОЙ
АКТИВНОСТИ В АНСАМБЛЕ НЕЙРОНОВ
Возможный механизм генерации альфа-частотной веретенообразной активности, основанный на пейсмекерных свойствах нейронов, исследовался в физиологическом и численном экспериментах. Проведена классификация веретенообразной активности, наблюдаемой в электрофизиологических экспериментах. Предложен механизм генерации альфа-частотной веретенообразной активности в ансамбле нейронов, объясняющий все формы веретена, а не только классическую.
Ключевые слова: веретенообразная активность, модель пейсмекерного нейрона, бифуркационный анализ
Введение
Ритмогенез является одним из фундаментальных свойств всех живых организмов, обеспечивающее поддержание их внутренней среды и адаптивное поведение. Эндогенные пейсмекерные осцилляции были зарегистрированы у одноклеточных эукариот [1], в онтогенезе млекопитающих ритмогенез в сердечной мышце проявляется еще до нервной стадии развития организма. Выраженный ритмический характер имеет электрическая активность мозга. Каковы механизмы этой активности? В частности, каковы механизмы спонтанной веретенообразной альфа-частотной активности головного мозга? Согласно наиболее распространенной гипотезе Андерсена и Экклса [2], кора является вторично вовлекаемой в ритмогенез структурой мозга. Согласно данной гипотезе, длительная гиперполяризация при формировании веретена обусловлена частичным суммированием ТПСП [3].
Однако, несмотря на то, что данные представления получили достаточно широкое распространение, существуют серьезные основания, позволяющие сформулировать предположения о том, что в основе возникновения сонных веретен лежит совершенно иной механизм, обусловленный не цикличным чередованием ВПСП и ТПСП, а эндогенной пейсмекерной активностью, формируемой потенциал-зависимыми K+ -, Ca2+-и Na+-каналами, встроенными в мембрану нейронов. Предположительно, и в таламусе и в коре могут существовать латентные пейсмекерные клетки, которые в эволюционном плане являются «наследниками» пейсмекерных свойств одноклеточных организмов. Латентные пейсмекерные клетки коры при определенных условиях способны генерировать ритмические осцилляции. Одним из таких условий является функциональное состояние коры головного мозга, которое определяется множеством факторов, в том числе, уровнем ее постоянного потенциала. Последний обусловлен локальным метаботропным влиянием подкорковых структур на кору. Проведенные нами исследования зависимости ритмогенеза от постоянного потенциала коры показали, что смещение последнего в сторону позитивности (что соответствует гиперполяризации отдельных клеток) сопровождается появлением спонтанных сонных веретен в соматосенсорной коре мозга крысы, которые могут возникать независимо от характера активности таламических ядер, имеющих проекции к этим областям [4].
Постановка задачи
Целью данной работы являлось исследование ритмогенеза альфа-частотной веретенообразной активности в физиологическом и численном экспериментах. На основе анализа экспериментальных данных по регистрации фокальной активности коры мозга крысы необходимо выдвинуть гипотезу, объясняющую все формы альфа-частотной веретенообразной активности. А также, обосновать данную гипотезу математическими методами.
Методы
Электрофизиологические микроэлектродные исследования биоэлектрической активности нейронных колонок соматической коры проводились на лабораторных белых крысах из вивария НИИ Нейрокибернетики ЮФУ по методике, одобренной комитетом по биоэтике РАН (протокол от 01.01.2001).
На основе разработанной ранее модели нейрона [5] была построена нейронная сеть, нейроны которой связаны попарно электрическими синапсами [6].
Результаты
Классической веретенообразной активностью называется осцилляторный процесс, состоящий из трех фаз, последовательно следующих друг за другом:
• фаза нарастания веретена, характеризуется постепенным увеличением амплитуды осцилляций;
• срединная фаза веретена, осцилляции с постоянной амплитудой;
• фаза завершения веретена, характеризуется постепенным угасанием амплитуды осцилляций вплоть до полного прекращения осцилляторного процесса.
Проведенный анализ электрофизиологических данных выявил наличие спонтанных осцилляций фокального потенциала коры мозга крысы на частоте альфа-ритма. Форма данных осцилляций часто отличалась от формы классической веретенообразной активности. Отличия эти заключались либо в отсутствии одной из трех выше перечисленных фаз, либо в отсутствии двух фаз, либо представляли собой циклические чередования всех трех фаз. Это позволяет обобщить понятие веретенообразной активности следующим образом. Альфа-частотной веретенообразной активностью называется осцилляторный процесс на частоте альфа ритма содержащий, по крайней мере, одну ярко выраженную фазу. Таким образом, под понятие веретенообразной активности попадает широкий класс явлений:
классическое веретено;
веретено с отсутствием ярко выраженной фазы нарастания веретена (правое полуверетено);
веретено с отсутствием ярко выраженной фазы угасания веретена (левое полуверетено);
веретено с длинной средней частью, с постоянной амплитудой и частотой;
атипичное веретено, несколько веретен перечисленной ранее формы, следуют друг за другом с модулированными по амплитуде спайками.
Неклассические формы веретенообразной активности (2,3,4,5) наблюдаются в тех же областях коры мозга крысы и при тех же функциональных состояниях коры, что и классические веретена, а также часто сменяют друг друга в течение одного эксперимента.
Выдвижение гипотезы единого механизма генерации различных форм веретенообразной активности
Желательно, в рамках одной математической модели охватить все формы веретен, а не объяснять каждую форму отдельным механизмом или моделью. Поэтому правдоподобная модель веретенообразной активности должна объяснять все формы, а не только классическую форму веретена. Предположительно, в основе механизма генерации веретенообразной активности может лежать реакция мембранного потенциала нейрона на некоторое несинаптическое воздействие. Фаза нарастания веретена происходить в результате монотонного воздействия одной направленности, фаза завершения веретена так же происходит в результате монотонного воздействия, но противоположной направленности, а срединной фазе веретена соответствует постоянное воздействие.
Данное воздействие является несинаптическим и представляет собой ток, который может иметь эндогенную природу в живой клетке. Моделируется данный ток дополнительным слагаемым в уравнении мембранного потенциала нейрона, также как и эндогенные ионные токи мембраны. Далее будет показано, что форма веретена модельного нейрона коррелирует с формой тока.
Ранее, численным интегрированием системы дифференциальных уравнений, описывающей модельный нейрон, было показано, что модель нейрона обладает пейсекерными свойствами при некоторых условиях. Данным условием является гиперполяризирующий ток из некоторого диапазона, данный диапазон назван диапазоном осцилляций [5]. Методом бифуркационного анализа системы уравнений показано, что наличие диапазона осцилляций обуславливается собственными значениями матриц линеаризации системы дифференциальных уравнений, описывающей нейрон при различных значениях тока гиперполяризации. Существование устойчивого мембранного потенциала модельного нейрона означает, что система дифференциальных уравнений, описывающая динамику модельного нейрона, имеет устойчивое равновесие. Это равновесие является следствием равенства нулю всех производных системы. Приравняв к нулю правые части системы дифференциальных уравнений, мы получим систему алгебраических уравнений, переменные которой являются функциями времени t. Данную систему можно рассматривать как однопараметрическое семейство систем. В качестве параметра используется гиперполяризирующий ток Iinjection(t). Этот ток является внешним для системы и влияет на её динамику. Изучение влияния этого параметра на состояние системы показало, что если внешний ток Iinjection(t), подаваемый на нейрон, является постоянной функцией времени Iinjection(t)= Iinjection, система дифференциальных уравнений, описывающая нейрон, представляет собой однопараметрическое семейство автономных систем дифференциальных уравнений с пятью неизвестными функциями времени, одной из которых является мембранный потенциал. Параметром семейства является гиперполяризирующий внешний ток Iinjection. При значении параметра Iinjection=0 получаем нелинейную систему алгебраических уравнений, решение которой дает точку равновесия. Точкой равновесия является вектор: (E=-62., mCa=0., hCa=0., mH=0.). В частности, при отсутствии внешних воздействий мембранный потенциал модельной клетки равен -62. mV. Для определения устойчивости данной точки равновесия был использован первый метод Ляпунова. Построена матрица линеаризации системы в данной точке равновесия при рассматриваемом значении параметра Iinjection и вычислены ее собственные значения. Метод Ляпунова исследования систем по первому приближению позволяет утверждать, что если все вещественные части собственных значений линеаризованной системы в точке равновесия строго меньше нуля, то равновесие нелинейной системы асимптотически устойчиво. Проведенные вычисления показывают, что все вещественные части собственных значений строго меньше нуля. Следовательно, можно утверждать, что равновесие асимптотически устойчиво при данном значении параметра.
Проведение серии аналогичных вычислений при разных значениях параметра Iinjection показало, что при любых значениях последнего на интервале [0:-30] система имеет положение равновесия, на интервале [0: -14] система дифференциальных уравнений имеет решение, асимптотически устойчивое по Ляпунову, на интервале [-14: -15] вещественная часть одной пары комплексно сопряженных собственных значений проходит через нуль и становится положительной. Следовательно, при значении параметра -15 система имеет неустойчивое положение равновесия. Аналогично, на всем интервале [-15:-19] система уравнений имеет неустойчивое положение равновесия. Таким образом, на интервале [-14:-15] имеется точка, в которой вещественная часть обращается в нуль. Эта точка была определена методом обратной интерполяции. Значение параметра (гиперполяризирующего тока), при котором происходит потеря устойчивости решения системы дифференциальных уравнений, оказалось равным -14.2823. Это значение гиперполяризирующего тока было обозначено как порог возникновения осцилляций, Thr, поскольку при прохождении значения гиперполяризирующего тока через это значение происходит потеря устойчивости равновесия и образование периодического решения. Образование периодического решения следует из того, что в этой точке выполняются условия теоремы Хопфа. Непосредственная проверка показала, что матрица линеаризации системы при значениях Iinjection из некоторой окрестности точки Thr имеет пару комплексно-сопряженных собственных значений. Вещественные части этих собственных значений обращаются в нуль в точке Thr, а производные по параметру вещественных частей собственных значений отличны от нуля. Поскольку остальные собственные значения имеют ненулевые действительные части, согласно теоремы Хопфа при параметре Thr от положения равновесия ответвляется однопараметрическое семейство замкнутых траекторий, отвечающих периодическим решениям системы уравнений. С физиологической точки зрения, при прохождении параметра через Thr возникают осцилляции мембранного потенциала.
Единичный модельный нейрон способен генерировать веретенообразную активность под воздействием тока гиперполяризации определенной формы рис. 1.
Форма данного тока отвечает требования выдвинутой гипотезы. График рисунка 1 также демонстрирует нарастание амплитуды внутриверетенных колебаний, что согласуется с экспериментальными данными [7]. Осцилляторная веретенообразная активность, полученная в вычислительном эксперименте, в основных чертах сходна с классическим веретеном, наблюдаемом в коре мозга крыс рис. 2.
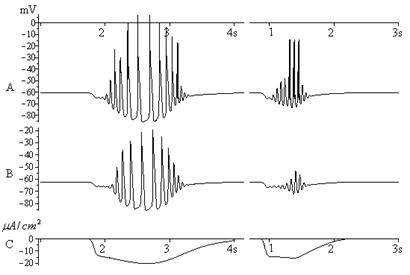
Рис. 1. Веретенообразная активность в двух не связанных нейронах. A: модель
без натриевых токов. B: модель с натриевыми токами. С: ток гиперполяризации.
Данный ток подавался на оба нейрона
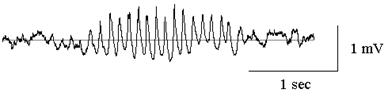
Рис. 2. Фокальная ритмическая активность сенсорной коры крысы. Спонтанное
веретено возникает при небольшом позитивном отклонении постоянного
потенциала коры
Пример веретенообразной активности в единичном нейроне при натриевых токах, отличных от нуля, приведен на рис. 1, A. В момент возникновения веретена натриевые потенциалы действия отсутствуют и появляются на кальциевых скайпах лишь в центральной части веретена, что соответствует физиологическим данным [7,8]. Таким образом, наличие натриевых каналов не является обязательным условием развития веретенообразного ритмогенеза в клетке, в основе которого может лежать сугубо эндогенный механизм.
В разработанной нами модели форма веретена коррелирует с формой тока. Механизм веретенообразной активности в единичном нейроне заключается в "реакции" нейрона на ток гиперполяризации: фаза нарастания веретена обусловлена появлением гиперполяризирующего толчка тока, амплитуда и период внутриверетенных осцилляций зависят от "глубины" (максимального абсолютного значения) тока гиперполяризации. Завершение («спад») веретена происходит при уменьшении абсолютной величины тока гиперполяризации. В зависимости от длительности, глубины и формы гиперполяризирующего тока в отдельных модельных нейронах можно наблюдать веретена различной формы и длительности. Варьируя параметры тока можно промоделировать все, перечисленные выше, формы альфа-частотной веретенообразной активности. Например, существует ток, в результате воздействия которого фаза нарастания веретена практически отсутствует (рис. 3, A, B), что нередко наблюдается в электрофизиологических экспериментах (рис. 3, C).

Рис. 3. Веретенообразная активность неклассической формы.
Резкое возникновение веретена и плавное завершение. А: мембранный потенциал модельного нейрона. В: ток, инжектируемый на нейрон. С: фокальная активность моторной коры мозга крысы
Согласно гипотезе Андерсена и Экклса [2], фаза нарастания веретена происходит за счет вовлечения новых нейронов в осцилляторный процесс и предполагает активное участие химических синапсов и натриевых каналов. Завершение веретена связано с десинхронизацией осцилляторной активности нейронов, участвующих в ритмогенезе. Однако, точный механизм, обуславливающий завершение веретена, остается до сих пор не выясненным в рамках данной гипотезы. В рамках данной гипотезы трудно объяснить неклассические формы веретенообразной активности.
Ранее нами в вычислительном эксперименте была исследована возможность синхронизации пейсмекерной активности в ансамбле нейронов [6]. Механизм ритмогенеза веретенообразной активности в ансамбле нейронов оказался связан с синхронизацией веретенообразной активности отдельных нейронов ансамбля посредством электрических синапсов (рис. 5).
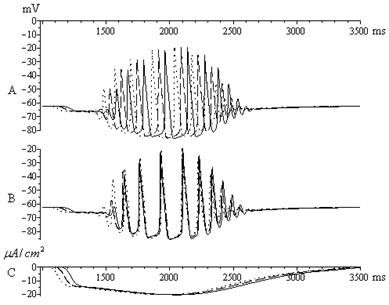
Рис. 5. Синхронизированная веретенообразная активность в ансамбле нейронов. A: колебания мембранных потенциалов трех не связанных нейронов.
B: веретенообразная активность в ансамбле из трех нейронов,
взаимодействующий посредством электрических синапсов.
На графике С изображены соответствующие формальным нейронам
токи гиперполяризации
Заключение
Настоящее исследование показало, что натриевые каналы играют лишь второстепенную роль в развитии веретенообразной активности единичной клетки и что модельный нейрон проявляет свойства генератора, управляемого током. Период активации низкопороговых кальциевых спайков, их амплитуда, частота и амплитуда быстрых натриевых спайков, генерируемых на кальциевых спайках, являются функциями входного тока. Причем, при определенных значениях гиперполяризирующего тока: 1) пейсмекерные колебания мембранного потенциала модельного нейрона являются подпороговыми, т. е., не достигают порога генерации натриевых спайков, 2) колебания мембранного потенциала модельного нейрона имеют частоту альфа-ритма, например, при значениях тока ![]() частота осцилляций равна 11. Гц, а при токе
частота осцилляций равна 11. Гц, а при токе 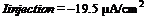 , равна 6.92983 Гц. Методами математического моделирования доказано существование незатухающих колебаний мембранного потенциала модельного нейрона при определенных условиях. Параметры модельного нейрона, а именно, плотности распределения ионных каналов на локусе мембраны, сопротивление мембраны, концентрации ионов кальция, были подобраны с учетом физиологических данных, полученных экспериментально. Следовательно, есть основания утверждать, что в коре мозга существуют локусы пейсмекерной активности, способные при определенных условиях генерировать пейсмекерные колебания с частотой альфа-ритма. Такие локусы могут располагаться в пределах одной корковой колонки, что может определять идентичность метаботропных влияний на них со стороны подкорковых. Кроме того, между ними могут существовать электротонические связи. При достаточном количестве локусов пейсмекерной активности и достаточной плотности электрических синапсов между ними ритмические колебания с частотой альфа-ритма могут носить локальный характер и наблюдаться лишь в пределах одной корковой колонки.
, равна 6.92983 Гц. Методами математического моделирования доказано существование незатухающих колебаний мембранного потенциала модельного нейрона при определенных условиях. Параметры модельного нейрона, а именно, плотности распределения ионных каналов на локусе мембраны, сопротивление мембраны, концентрации ионов кальция, были подобраны с учетом физиологических данных, полученных экспериментально. Следовательно, есть основания утверждать, что в коре мозга существуют локусы пейсмекерной активности, способные при определенных условиях генерировать пейсмекерные колебания с частотой альфа-ритма. Такие локусы могут располагаться в пределах одной корковой колонки, что может определять идентичность метаботропных влияний на них со стороны подкорковых. Кроме того, между ними могут существовать электротонические связи. При достаточном количестве локусов пейсмекерной активности и достаточной плотности электрических синапсов между ними ритмические колебания с частотой альфа-ритма могут носить локальный характер и наблюдаться лишь в пределах одной корковой колонки.
Список литературы
1. Греченко эндогенных осцилляторов в организации памяти// Материалы XV международной конференции по нейрокибернетике. 2009. Т.1. С. 91.
2. Andersen P., Eccles J. Ingibitory phasing of neuronal discharge // Nature. 1962. V. 196. P. 645-647.
3. Гусельников головного мозга, М.: Высшая школа. 1976.
4. Дис. ... канд. биол. наук // ЮФУ. Ростов-на-Дону, 2011.
5. , , Кирой ритмогенеза веретенообразной активности в единичном нейроне // XII Всероссийская научно-техническуя Конференция «Нейроинформатика-2010»: Сб. научн. трудов. 2010. Ч.1. С.105-112.
6. , , Кирой ритмогенеза в ансамбле кортикальных нейронов, связанных электрическими синапсами // Нейрофизиология. 2011. № 5. C. 399-407.
7. , , Коняхина механизм генерации веретенообразной активности в колонках соматической коры крысы // ВЕСТНИК ЮЖНОГО НАУЧНОГО ЦЕНТРА РАН. 2007. Т. 3..№ 2. C. 86-94.
8. Timofeev J., Bazhenov M., Seinowski T. J., Steriade M. Contribution of intrinsic and synaptic factory in the desynchronization of thalamic oscillatory activity // Thalamus & Related Systems. 2001. V. 1. P. 53 – 69.
