

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ЛЕКЦИЯ 17. РОЛЬ ХРОМОСОМНЫХ ПЕРЕСТРОЕК В ЭВОЛЮЦИИ. ЭФФЕКТ ПОЛОЖЕНИЯ (незавершенный черновик!)
, ИЦиГ СО РАН и ФЕН НГУ, Новосибирск, 2012 г.
17.1. Роль инверсий и транслокаций в эволюции.
17.1.1. Общие замечания
Совсем недавно, каких-нибудь полвека назад, в научном мире царила атмосфера опьянения от победы дарвинизма. Я убежденный дарвинист и считаю, что это основа всей биологии и содержание его негативно: состоит оно в том, что специальных механизмов эволюции не существует и вся биологическая эволюция сводится к случайным процессам, с неизбежностью происходящим в мире не вполне точно самовоспроизводящихся открытых систем, помещенных в конкретную среду. Недавно же царил так называемый панселекционизм, когда было принято считать, что все существующее существует исключительно для повышения Дарвиновой приспособленности. При этом любой особенности любого организма придумывалось объяснение с точки зрения его полезности, и никого не смущало, что столь же легко можно было бы придумать объяснение и для чего-то противоположного. Вот и цитологи старой школы обычно убеждены, что раз хромосомные перестройки существуют, значит они для чего-то очень нужны. Определенная тенденция переоценивать их значение сохранилась и до сих пор.
Неудивительно, что имеет место тенденция переоценки значения и такого результата хромосомных перестроек, как изменение гаплоидного числа хромосом в эволюции, в частности в видообразовании. Хочу заметить, что речь идет не более чем о переупаковке генома в то или иное количество чемоданов, общее содержание которых никак не меняется, и даже как правила не меняются условия его хранения. Различия в числе хросомом могут опять таки сказываться на протекании мейоза у гибридов (о чем пойдет речь ниже), что может действительно способствовать репродуктивной изоляции. Но и этот эффект склонны переоценивать, так как проблемы в мейозе гибридов происходят не всегда.
Среди хромосомных перестроек наиболее часто встречаются инверсии и транслокации. Именно встречаются, в качестве внутривидового полиморфизма, а не возникают – рассмотренные выше варианты возможныХ-хромосомных перестроек скорее всего имеют другие вероятности – не исключено, что вероятнее всего происходит обмен с сестринской хроматидой или гомологом. Однако данные на этот вопрос было бы сложно получить в связи с летальностью многих типов образующихся перестроек. Инверсии и транслокации – это именно те два класса перестроек, которые не изменяют состав и качество генетической информации, то есть это те перестройки, которые виды могут позволить себе иметь. Но аномалии в мейозе – вещь серьезная, и против них должен действовать естественный отбор. Тем не менее, многие виды не спешат избавляться от полиморфизма по таким перестройкам, и он едва ли не поддерживается естественным отбором. В особенности часто встречается полиморфизм по инверсиям. В чем же может состоять возможная польза инверсий и транслокаций?
Как мы убедились, инверсии резко сокращают или совсем запирают кроссинговер в гетерозиготе, так что находящиеся в пределах инверсии аллели начинают наследоваться как одно целое. Это открывает хорошую возможность к созданию супергенов. Удачные сочетания аллелей сцепленных локусов будут наследоваться как одно целое, без рекомбинации. У гомозигот по инверсии кроссинговер будет идти без нарушений, но предполагается, что и аллели этих локусов будут находиться в гомозиготе, то есть удачное сочетание не будет нарушено. При протяженных инверсиях, способных создавать петли при спаривании в профазе мейоза, ситуация ухудшается тем, что любоее нечетное количество кроссоверных событий, затрагивающих одну хроматиду, превращает ее в нежизнеспособную перестройку, так что фертильность таких гетерозигот падает. Однако кроссинговер можно вообще исключить если, к примеру, добавить внутрь инверсии еще одну инверсию. Это резко ограничит способность к образованию петель и зону гомологичного спаривания внутри охватывающей инверсии, то есть сделает сам кроссинговер редким событием, что восстановит фертильность гетерозигот. Как мы увидим, по тому же пути пошел и экспериментатор при создании балансерных линий. Более того, само по себе гетерозиготное состояние значительного участка генома может быть выгодно за счет эффекта гетерозиса, что может создать предпосылку для сбалансированного полиморфизма. Однако такое возможно только в случае сверхдоминирования по какому-то локусу; тогда как более вероятная интерпретацию гетерозиса как следствие высокой средней гетерозиготности, что снижает вероятность выхода в гомозиготу гипо - или аморфных аллелей важных генов, не поддерживает нашу модель, так как инверсии и блоки находящихся в них сцепленных аллелей будут оказываться также и в гомозиготе.
17.1.2. Хромосомные комплексы Реннера
Казалось бы, транслокации не дают даже и такого выигрыша – кроссинговер подавляется лишь в непосдерственной близости от точки разрыва, а фертильность гетерозигот падает вдвое. Тем не менее известен почти невероятный случай облигатного полиморфизма по множественным транслокациям, который обеспечивает постоянную гетерозиготность. При всей экзотичности этого случая он попал в поле зрения генетики в самый момент ее переоткрытия и даже привел к появлению термина «мутация», хотя само явление, как выяснилось, не имело ничего общего с тем понятием изменения генов, которое имел в виду предложивший термин Де Фриз и которое вкладывается в него и по сию пору.
Есть такое растение из семейства кипрейных – ослинник. У нас в Сибири оно едва появляетсякак заносный сорняк только в последнее время, причем это тот же самый ослинник двулетний (Oenothera biennis),



который с XVII века разводится в Европе как овощ, известный под названием «рапонтика» или «рапунцель»; этот же и другие виды разводятся в качестве декоративных цветов. Род этот по происхождению чисто американский. Русское название он получил в качестве перевода устаревшего латинского названия Onagra, что означает «пища кулана», в генетике же он известен по своему гораздо более благозвучному валидному латинскому названию энотера (Oenothera), данному позднее Линнеем и означающему, по-видимому, «ловец ослов» по-гречески. В орбиту генетики попали удивительные виды из подрода Euoenothera. Считается, что плейстоценовые похолодания и потепления в Северной Америке, сопровождавшиеся миграционными волнами, привели к встрече не до конца дивергировавших популяций, в результате чего сформировалась следующая поразительная картина. Все энотеры имеют 2n=14 метацентрическиХ-хромосом, однако у многих видов (деление на которые в данном случае условно) большинство или все хромосомы вовлечены в реципрокные обмены в прицентромерном гетерохроматине, причем каждая особь всегда гетерозиготна по всем этим транслокациям. Если обозначить плечи хромосом цифрами от 1 до 14, то, в случае, когда вовлечены все – например у Oenothera muricata и O. laevigata, один родительский кариотип будет содержать хросомомы 1-2, 3-4, 5-6, 7-8, 9-10, 11-12, 13-14, а второй – 2-3, 4-5, 6-7, 8-9, 10-11, 12-13, 14-1. Получается, что в кариотипе таких видов вообще нет ни одной пары гомологичныХ-хромосом. Нетрудно понять, что получается в профазе первого деления мейоза – это будет гептавалент, в котором все эти хромосомы будут спарены, чередуясь друг с другом. В пахитене он очень похож на звезду. У O. lamarckiana в круговой обмен плечами вовлечены 12 из 14 хромосомных плеч, а два плеча образуют пару свободных гомологов
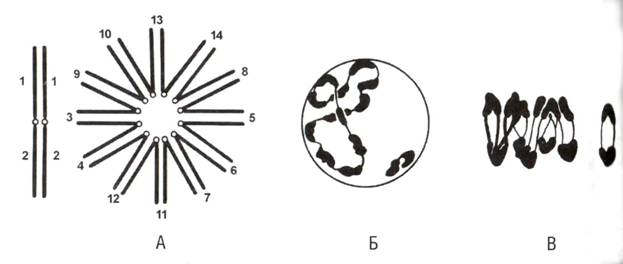
У нашей знакомой O. biennis в транслокации вовлечены все хромосомы, но образуется два кольца, а не одно. У других видов в круговой обмен вовлечено еще меньше хромосом.
Сложнее понять, как такая структура будет расходиться в анафазе. Ведь даже гетерозигота по одной транслокации делает половину гаметофитов нежизнеспособными, а тут их целых семь, что, казалось бы, уменьшает вероятность жизнеспособных продуктов мейоза до катастрофически низкого уровня. (По какой-то причине этот вопрос замалчивают все наши учебники по генетике.) Однако тут на помощь приходит механическая функция кроссинговера, причем путем отказа от его рекомбинационной функции. Все хиазмы у ослинника локализованы на концаХ-хромосом. Это делает центромеры механически свободными друг от друга, и гептавалент спонтанно находит состояние, обеспечивающее максимальное натяжение микротрубочек, а следовательно, максимальную стабильность микротрубочек, каковое и сохраняется в метафазе. Это состояние, при котором центромеры притягиваются к разным полюсам деления через одну. В результате, начиная с диакинеза и в метафазе мы вместо звезды наблюдаем мы уже кольцо из 14 хромосом, соединенных концами плеч. Как следствие, в первом делении мейоза к одному полюсу пойдет в полном составе один родительский геном, а к другому – другой. В этой ситуации как одно целое наследуется весь гаплоидный геном, превращающийся в один суперген. Эти совместно наследуемые как одно целое гаплоидные наборы хромосом были названы комплексами Реннера. Казалось бы, для оправдания такой системы каждый геном должен содержать очень удачное сочетание аллелей. Но все оказывается еще хуже – удачным оказывается лишь гетерозиготное состояние. Чтобы обеспечить его воспроизводство, каждый их геномов содержит летали. Причем у разных видов в них фиксируются разные летали – у O. biennis это специфичные для пола гаметофитные летали – один хромосомный набор оказывается летальным для семяпочек, другой – для пыльцы, в результате все семяпочки несут только один родительский набор, а вся пыльца – только другой, и все спорофиты образуются только гомозиготными. У O. lamarckiana родительские хромосомные наборы несут разные рецессивные спорофитные летали, так что леталь в одном наборе перекрывается нормальным аллелем в другом. Гомозиготы по каждому из наборов не выживают. Данные по гомозиготным видам энотеры дают основания полагать, что прителомерная локализация хиазм и, как следствие, потеря мейотической рекомбинации эволюционно предшествовала транслокациям и способствовала их возникновению.
Да, а как же мутации, открытые Де Фризом у O. lamarckiana? Под ними он понимал одноактные спонтанные и в дальнейшем наследуемые изменения наследственных факторов, в результате которых один вид превращается в другой. В течение семи лет на материале около 53 тысяч растений он нашел таких мутаций около 800 – т. е. 1,5 %! Причем многие из них были идентичными. На самом деле он столкнулся не с мутациями в нашем современном понимании, а со спонтанными сбоями правильного наследования комплексов Реннера, в частности, с нерасхождениями, когда одно из колец «прихватывает» хромосому из второго кольца между двумя своими с образованием первичных трисомиков, что встречается не так уж редко,
а также с изменениями плоидности. Де Фриз опубликовал свою двухтомную мутационную теорию в, а умер в 1935 г. и в учебниках почему-то замалчивается тот факт, что он сам участвовал в выяснении природы своих мутаций. В журнале Genetics за 1922 г. статья его и Boedijn, где он классифицирует свои мутации на истинные рецессивные мутации и мутации со сверхчисленными хромосомами, в основном трисомики, но также множественные трисомики и полиплоиды.
Пример с энотерой демонстрирует не только мощь адаптивной эволюции, но одновременно и ее бессмысленность и как минимум ненаправленность. Его принято рассматривать как апофеоз преимуществ высокой гетерозиготности и полного закрепления гетерозиса. Но ведь сам по себе гетерозис объясняется низкой вероятностью выхода в гомозиготу аморфных мутаций. В данном же случае растение пошло на то, чтобы для обеспечения постоянной гетерозиготности специально фиксировать летали и выводить их в гомозиготу, в каждом поколении «преднамеренно» теряя половину своих гамет. Кроме того, вся система комплексов Реннера смогла возникнуть лишь у растений, где кроссинговер исключен везде, кроме концов хромосом, а сама она исключает независимую сегрегацию хромосом в мейозе. То есть виды, наслаждающиеся постоянной гетерозиготностью, пошло на то, чтобы вообще расстаться с рекомбинацией, как кроссоверной, так и сегрегационной, полезность которой обычно считается настолько фундаментальной, что она явилась причиной возникновения самого полового размножения.
Комплексы Реннера существуют и в некоторых других родах того же семейства Onagraceae, а также описаны у некоторых скорпионов и кузнечиков. С таким же явлением мы сталкивались и в случае половыХ-хромосом ехидны, у самцов которой в мейозе образуется гептавалент (совпадение с числом хромосом у энотеры - случайно).
17.3. Последствия хромосомных перестроек для структуры генома
17.3.1. Синтения
В наш XXI век геномных проектов в огромных количествах начала поступать информация о том, что собой представляет первичная структура ДНК каждой из хромосом анализируемого объекта (количество которых растет экспоненциально), включая набор, порядок и взаимную ориентацию генов, блоков повторов и уникальных некодирующих отрезков. Эти проекты сразу же столкнулись с проблемой индивидуальной изменчивости (и начались с вопроса «чей персонально геном секвенировать»). Эта изменчивость распадается на два класса: нуклеотидные замены в гомологичных последовательностях и изменения состава и/или порядка следования гомологичных последовательностей, то есть как раз хромосомных перестроек. При секвенировании геномов выявляются в том числе очень небольшие перестройки, недетектируемые на цитологическом уровне. Однако особи одного вида с необходимостью достаточно родственны друг другу, имея последнего общего предка не столь давно. Например все люди имели его примерно 80 тысяч лет назад; вернее, можно говорить об облаке общих предков в пространстве и времени, так как для каждого фрагмента генома он будет свой. Последний общий предок разных видов и прочих таксонов как правило отстоял во времени от современного момента гораздо дальше, тем дальше, чем выше ранг таксона, хотя вот здесь возможны самые удивительные несоответствия, так как все таксономические ранги - условны. За это время в ведущих к ним разных эволюционных ниточках могли зафиксироваться разные совместимые с жизнью хромосомные перестройки. Мы можем их отследить при сравнении геномов, однако лишь до таких эволюционных дистанций, когда мы еще сможем распознать неперестроенные фрагменты геномов, сохранившие схожую структуру, имевшуюся у их общего предка – прежде всего состав, порядок и ориентацию ортологичных локусов первоначальную структуру. Такие фрагменты называют синтенными , а само явления сходства состава, порядка и ориентации локусов какого-либо участка генома организмов вследствие общности их происхождения называется синтенией. Синтению можно считать косвенно связанной с «законом гомологических рядов» Вавилова – родственные организмы имеют не только сходные гены, но и сходный их порядок, или иными словами сходные генетические карты, с точностью до того или иного количества хромосомных перестроек, произошедших после их дивергенции.
Синтения имеет теоретическое и практическое значение. С теоретической точки зрения она дает возможность отслеживать хромосомные перестройки и скорость, с которой они фиксируются в ходе эволюции. Кроме того, сохранение синтении каких-то участков генома в течение длительного времени и у многих организмов может указывать на нечастые случаи, когда близкое расположение и даже порядок каких-то генов может иметь значение для их функционирования – допустим, этот район имеет особую структуру хроматина или в нем присутствуют некие цис-взаимодействия генов.
Мы могли бы наблюдать синтению и оценивать ее степень, даже сравнивая рекомбинационные генетические карты родственных организмов. Однако подробные генетические карты существуют пока только для очень немногих модельных организмов. То же верно и для физических карт, основанных на чтении полных геномов. При этом физические карты появляются с нарастающей скоростью для все большего числа видов, в связи с прогрессом и удешевлением высокопроизводительного секвенирования, а рекомбинационные карты требуют исследований по частной генетике объекта, включая создание генетических коллекций, в течение многих десятилетий, и такая работа сейчас непопулярна. Поэтому практическое значение имеет обратная задача – экстраполяция известных генетических карт на родственные виды, у которых генетические карты отсутствуют. В качестве нулевой гипотезы при этом принимается синтения, и исходя из нее мы можем предсказывать местонахождение тех или иных генов в геноме такого объекта и действительно находить их. Вот довольно яркий пример: у гороха есть подробные рекомбинационные генетические карты, однако геном гороха не прочитан. В то же время у бобовых растений полностью прочитан геном лишь модельного вида люцерна обрезанная Medicago truncatula, у которой он очень невелик, а частично - у японского лядвенца Lotus japonicus и у сои Glycine max. Все они находятся в других трибах семейства бобовых, чем горох. В то же время значительные участки генома у всех бобовых сохраняют синтению. Многие из генов, стоящих у гороха на генетической карте, все же идентифицированы на молекулярном уровне. Это позволяет искать их ортологи у видов с просеквенированным геномом и, убедившись в сохранении синтении, использовать информацию о сцеплении для идентификации генов гороха, известных пока только по своему фенотипическому проявлению. Например, если у гороха изучаемый ген стоит между маркерами с известной первичной структурой, можно найти их ортологи у люцерны и перебрать все гены – по сути открытые рамки считывания – угадывая по первичной структуре и предполагаемой функции тот, который мог бы обеспечить подобное фенотипическое проявление. Генов на таком участке прочитанного генома (у нас – генома люцерны) обычно бывает очень много, а молекулярная функция многих неясна. Однако их можно использовать как молекулярные маркеры для сужения интервала уже у непросеквенированного вида (у нас – гороха) – разрабатывая праймеры к люцерне и применяя их на горохе. В конце концов можно сузить интервал до небольшого и указать искомый ген на молекулярном уровне, причем у обоих объектов сразу. Проверкой идентификации будет полное отсутствие рекомбинации предполагаемого гена с изучаемым фенотипическим проявлением и прочтение этого гена у мутантных линий. Такой подход называется метод генов-кандидатов. Применение этого метода ограничено районами, сохраняющими синтению, его применение исключает перестройка с точкой разрыва в интересующем районе, зафиксированная в одной из эволюционных линий, ведущих к сравниваниям видов. Однако одна точка разрыва разделяет синтенную область на две, и можно попытаться определить их границы у каждого из сравниваемых видов и продолжать пользоваться синтенией.
Синтению можно косвенным образом наблюдать и цитологически, используя сравнительный хромосомный пэйнтинг (comparative chromosome painting) или «Zoo Fish». Для этого флюоресцентно меченый ДНК-зонд, специфичный для из индивидуальной хромосомы одного из видов (который еще нужно подобрать), гибридизуют на кариотипе другого вида и смотрят, как распределены соответствующие последовательности по его хромосомам.
Чтобы примерно почувствовать с какой скоростью накапливаются перестройки, приведу картинку из книги Корякова и Жимулева, показывающую гомологию хромосом человека, шимпанзе и свиньи.
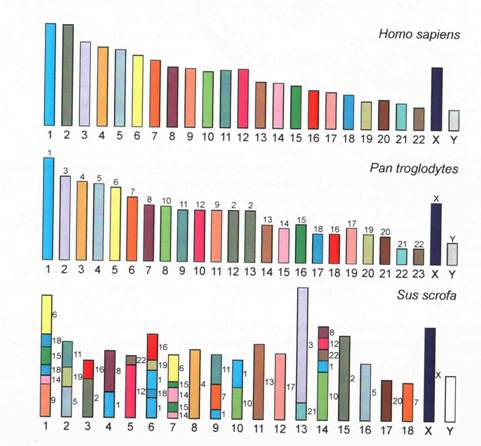
Человек и свинья имели последнего общего предка около 100 млн назад (заметтье, это было еще в меловом периоде), и в большинстве хромосом мы видим следы примерно 2-5 точек разрыва транслокаций (но совершенно не видим небольших инверсий). Но есть и хромосомы, полностью свободные от транслокаций! Всего мы насчитываем на этой картинке у свиньи 46 хромосомных фрагментов, синтенных человеческим (на уровне присутствия хромосомоспецифичной ДНК, но не порядка генов, который нам не известен). Получается в среднем 2 фрагмента, то есть одна транслокация, на однУ-хромосому гаплоидного набора человека (круглость цифры – чистая случайность). Если учесть, что эти транслокации могли зафиксироваться как в линии, ведущей к человеку, так и к линии, ведущей к свинье, у нас получается, что в среднем одна транслокация фиксируется за лет. И это притом, что носители транслокаций – по сравнению с нормальным кариотипом - попадаются даже среди нас, однако эти транслокации не зафиксировались у всех людей или свиней, как было с теми, которые сейчас отличают нас от свиней), а являются всего лишь морфами внутривидового полиморфизма, которые постоянно появляются в популяциях и исчезают из них.
При таких медленных темпах фиксации транслокаций выглядит даже странно, что за 5 млн лет со времени последнего общего предка человека и шимпанзе нам «удалось» зафиксировать одну Робертсоновскую транслокацию – центрическое слияние в линии человека. И в то же время недавно была опубликована потрясающая цифра, полученная в результате сравнения полностью прочитанных геномов человека и шимпанзе – в ходе дивергенции этих двух видов зафиксировалось 1,5 тысячи инверсий! (Впрочем, эта цифра сильно зависит от минимального размера, начиная с которого инверсии начинают учитываться. Снизить его до физического минимума в два нуклеотида невозможно, поскольку при очень малых размеров перестройки нельзя будет отличить от вариантов гомологического выравнивания участков генома с несколькими точковыми заменами.) Инверсии, особенно небольшие, не выявляются цитологическими методами, поэтому их количество долгое время недооценивалось. В то же время бОльшая частота инверсий, чем транслокаций не должна нас удивлять – ведь инверсия – это продукт обмена хромосомы сама с собой, когда партнер обмена находится рядом по определению, а для того, чтобы произошла транслокация, в момент разрыва должны встретиться в ядре две негомологичные хромосомы. Впечатляет здесь то насколько инверсии оказались чаще транслокаций. Популяционные исследования, проведенные на дрозофиле, у которой на цитологическом уровне на политенныХ-хромосомах можно наблюдать и небольшие инверсии, также показывают гораздо большую частоту инверсий, чем транслокаций.
После этого неудивительно, что принято различать синтению в узком смысле как нахождение генов в одной и той же хромосоме, синтению по сегменту – нахождение генов в одном и том же участке хромосом, и синтению по порядку – когда сохранен порядок между генами. Ведь инверсии эффективно перетасовывают гены в пределах одной хромосомы, тогда как транслокации переносят их в другую хромосому достаточно редко.
Впрочем, в разных группах различаются как минимум скорости транслокаций: например, Робертсоновские транслокации идут очень активно у грызунов и почти отсутствуют у хищных. Возможно, это связано с тем, что по неизвестной пока причине Робертсоновские триваленты расходятся у грузынов всегда правильно.
17.3.2. Дупликации ретротранспозонов.
Огромные последствия для структуры генома проистекают из такого специфического класса перестроек, как перемещение транспозонов. В особенности это касается дупликаций ретротранспозонов, поскольку при этом происходит увеличение количества их копий в геноме благодаря особенностей механизма их размножения, тогда как механизма уменьшения этих копий не существует. Надо сказать, что результаты весьма впечатляющи – геномы эукариот не менее чем наполовину (цифра сильно различается у разных организмов), а зачастую и на 90% состоят из копий ретротранспозонов, в подавшяющем большинстве дефектных, неспособных к перемещениям и находящихся на разных стадиях мутационной деградации, а осмысленная часть генома – то есть кодирующие последовательности и несомненные регуляторные некодирующие области - составляет всего лишь первые проценты (обычно не более 5%).
17.4. Эффект положения.
17.4.1. Общие замечания
17.4.2. Эффект положения конститутивного типа
Стёртевант. 1925. Bar. +/BB эффект сильнее, чем B / B. И там, и там – четыре копии, но когда 1 и 3 сильнее, чем когда 2 и 2.
Эффект Дубинина (Дубинин и Сидоров, 1934). Cubitus interruptus. Из прицентромерного гетерохроматина в эухроматин нормальный аллель из доминантного становится полудоминантным. То есть ослабление эксрессии в непривычном месте.
17.4.3. Эффект положения мозаичного типа
Position effect variegation
Мёллером в 1930 г. Inv(1)wm4
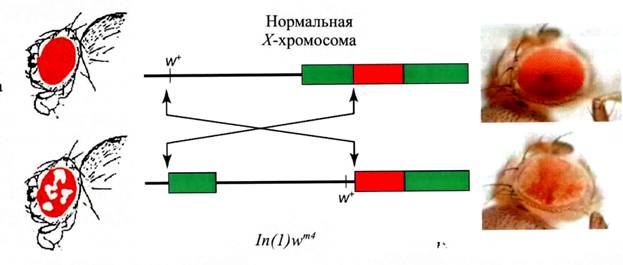
Перенос из эухроматина в гетерохроматин, но не в любой
Ген не страдает, можно вернуть и восстановится
Стохастика, все или ничего, клеточные клоны. На уровне транскрипции.
Множество перестроек. При перенесении в окрестности гетерохроматина много разных генов.
Эффект распространения (spreading effect) – вероятность падает, но инактивация все равно полная. Гетерохроматин докуда-то. На десятки сантиморганов, до 170 дисков, 5 мегабаз. Прерывистая инактивация (1991 г., ).
Им может стать любая ДНК, это состояние. Плотная упаковка, белковый состав (белок HP-1) поздняя репликация, эктопические контакты.
17.4.4. Модификация эффекта положения
Три группы модификаций – температура, количество гетерохроматина в клетке, мутантные аллели. Все действуют одновременно и скореллировано на процент инактивированных клеток и на дистанцию эффекта распространения.
Температура. 16оС белые, распространяется дальше, 25оС мозаичные, 29оС красные.
в чувствительный период. В раннем эмбриогенезе, как раз при формировании гетерохроматина. 16
Изменение количества гетерохроматина. Числа У-хромосомы. Гетерохроматиновая
Количество инактивирующего белка. На определенное количество мест инициации.
Варьировать дозу Н-хромосомы – схемы скрещиваний при помощи компаундов. (ХХ) без игрека слева берется справа.
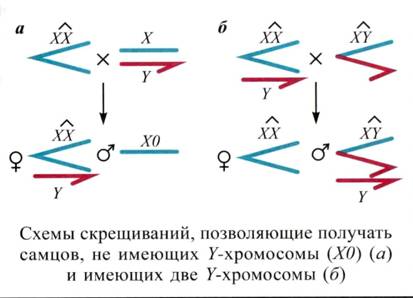

Супрессоры Su(var) и энхансеры En(var). Доминантные, скорее полудоминантные, зависящие от дозы. В гомозиготе обычно летальны. Супрессоры – инактивирующие, энхансеры – активирующие. Гапло-супрессор-трипло-энхансер и наоборот. Известно около десятка.
Самок с перестройкой, например той же самой. С мутагенизированными самцами, от которых аутосомы и У-хромосома. Смотрят у самцов
Среди них оказались гены многих структурных белков гетерохроматина. Гены белков гетерохроматина – супрессоры, гены белков эухроматина – энхансеры. Генетический анализ компонентов хроматина. Например Su(var)2-5 - тот самый белок НР-1 - и белков, которые могут влиять на принятие хроматином того или иного состояния. А Su(var)3-9 – метилирует гистон Н3, создавая сайт посадки НР-1. Сопряженным деметилированием Н3 по другому остатку занимается продукт Su(var)3-3.
Центры инициации, распространение за счет кооперативного эффекта, подобно росту кристалла, до терминатора. Нужны терминаторы, а тут его нет. В пользу – не всякая перестройка вызывает эффект положения. Формируется изначально в эмбриогенезе по закону все или ничего, потом лишь поддерживается эпигенетически, также за счет кооперативного эффекта. Почему по закону все или ничего – нельзя быть немного гетерохроматином, как немного беременным. Состояние – тонким балансом белковых факторов. Мозаичность – в зоне неопределенности, флуктуации локальной концентрации ключевых белков.
17.4.5. Использование транспозонов.
Изучение: выше - влияние разных белков на один случай эффекта. Сохраняется положение, меняется генетический фон.
Транспозоны. З-элемент с геном w+ на фоне ww. Как конститутивный, так и мозаичный. Чаще конститутивный, так как менее нарушена нормальная организация гетерохроматина в хромосоме и расположение терминаторов. Влияние разного гетерохроматина на одну инсерцию. Меняется положение гена, сохраняется генетический фон и ключевые точки в хромосомах.
Дрожжи. S. cerevisiae Ade2 синтез аденина. Встройка транспозона в теломеру – мозаичный эффект.
У Shizosacharomyces pombe Ade6 в прицентромерный или в инактивированный половой локус mating type
