
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Перевод статьи
Karlic H., PhD, and Lohninger A., PhD. Supplementation of L-сarnitine in athletes: does it make sense? // Nutrition 2004; 20: 709–715.
From the Ludwig Boltzmann Institute for Leukemia Research and Hematology, Vienna, Austria; and the Department of Medical Chemistry, University of Vienna, Vienna, Austria
Применение L-карнитина спортсменами:
имеет ли это смысл?
Х. Карлиц, А. Лонингер
Резюме
Исследования с участием спортсменов выявили, что применение карнитина может способствовать лучшему выполнению физических упражнений. Как было показано в большинстве исследований этого рода, увеличение потребления кислорода и снижение дыхательного коэффициента свидетельствуют о том, что прием карнитина стимулирует катаболизм липидов. Применение L-карнитина вызывает значительное снижение уровня плазматического лактата после нагрузки, образующегося и утилизируемого в полностью аэробных условиях. Получены предварительные данные о том, что применение карнитина приводит к ослаблению вредных последствий гипоксических тренировок и увеличению скорости восстановления после физических нагрузок. Результаты последних исследований позволяют говорить о способности карнитина предотвращать повреждения клеток и увеличивать их возможность восстанавливаться после стресса. Поглощение L-карнитина клетками крови может иметь несколько различных последствий:
1) стимуляция гематопоэза,
2) подавление индуцируемой коллагеном агрегации тромбоцитов,
3) предотвращение программируемой смерти клеток иммунной системы.
Как было показано недавно, карнитин способен напрямую регулировать экспрессию генов (карнитин-ацилтрансфераз) и модулировать концентрацию внеклеточных жирных кислот. Таким образом, получены свидетельства положительного влияния карнитина на тренировочный процесс, соревнования, восстановительный период после напряженных тренировок и регенерацию.
Ключевые слова: прием L-карнитина, метаболизм, физические упражнения.
Введение
Диетические добавки для увеличения результативности знакомы многим спортсменам. Производители с большим или меньшим упорством утверждают, что предлагаемые ими препараты увеличивают результативность спортсменов и/или скорость восстановления после нагрузок. Большинство таких утверждений носит спекулятивный характер и основано на представлениях о том, как тот или иной препарат воздействует на метаболизм. L-карнитин приобрел особую популярность как увеличивающий работоспособность препарат из-за участия в катаболизме жиров [1, 2].
L-карнитин был открыт двумя русскими исследователями [3], придумавшими название от латинского слова carnis (плоть или мясо). Его химическая структура была установлена в 1927 г., а в 1935 г. вышла первая научная статья [4], открывшая дорогу многочисленным исследованиям физиологической роли этого соединения. В 1959 г. Fritz показал, что карнитин интенсифицирует окисление жирных кислот в печени и сердце (5). Другое название карнитина – витамин Bт (T = tenebrio) – было дано этому соединению, когда было обнаружено, что он необходим для развития нимф черного таракана Tenebrio molitor (Tenebrionidae, Coleoptera) наряду с фолиевой кислотой и другими витаминами ряда B. По своей химической структуре L-карнитин (3-гидрокси-4-N, N,N-триметиламинобутират, L-3-гидрокси-4-N-триметиламинобутановая кислота, или γ-триметиламино-β-гидроксибутановая кислота) представляет собой холиноподобный четвертичный амин. В препаратах диафрагмального нерва карнитин вызывает тетанические сокращения диафрагмы, и этот эффект может быть частично подавлен добавлением холина [6].
Наиболее достоверно с научной точки зрения было показано участие карнитина в транспорте жиров через внутреннюю мембрану митохондрий. Среди других установленных функций карнитина – сохранение целостности мембран, стабилизация физиологического отношения коэнзима A (CoA) к ацетилкоэнзиму A (ацетил-СoA) в митохондриях, подавление образования лактата [7, 8]. In vitro было показано, что L-карнитин способен подавлять апоптоз (программируемую клеточную смерть) [9–11].
Внутриклеточное содержание карнитина контролируется различными мембранными транспортерами. Более важными являются транспортеры органических катионов (OCTNs), в частности OCNT2, осуществляющие кишечное всасывание и почечную реабсорбцию карнитина.
Они во многом определяют перераспределение карнитина в тканях и отвечают за интенсивность тканевого транспорта. Врожденные или приобретенные дефекты в системе карнитинового транспорта приводят к первичному или вторичному дефициту карнитина. Содержание мРНК OCTN2 уменьшается с возрастом [12] и под действием активных форм кислорода [13]. Некоторые соединения, вызывающие системный дефицит карнитина, ингибируют OCTN2 напрямую.
Вторичный карнитиновый дефицит часто наблюдается у пациентов с метаболическими нарушениями, регулярно проходящих гемодиализ [14], во время беременности [15].
L-карнитин широко доступен в продаже и часто используется спортсменами. Слухи о том, что применение L-карнитина помогло национальной сборной Италии выиграть чемпионство в 1982 г., безмерно увеличили его популярность. Самые серьезные ожидания возлагаются на участие карнитина в катаболизме жиров. Карнитин представляется как средство, усиливающее метаболизм жиров, снижающее массу жира и увеличивающее мышечную массу.
Карнитин также рекомендуется применять в условиях снижения веса. В тех видах спорта, где выносливость выходит на первый план, спортсмены используют карнитин, чтобы, усилив окисление жиров, сохранить мышечный гликоген. Данный обзор посвящен критическому анализу данных о роли карнитина в этих процессах.
Синтез и регуляция пула эндогенного карнитина
В организме млекопитающих карнитин образуется из незаменимых аминокислот лизина и метионина [16, 17]. Доступность интермедиата этого синтетического процесса – триметиллизина – ограничивает уровень образования карнитина.
Большая часть триметиллизина образуется в белках скелетных мышц. Таким образом, считается установленным, что скорость преобразования мышечных белков является лимитирующим фактором биосинтеза карнитина [18]. Последний этап синтеза, гидроксилирование бутиробетаина, происходит в печени, почках и мозге. Другие ткани получают карнитин из кровотока. Участники этого метаболического пути подробно описаны в работе Vaz [19].
Карнитин в человеческом организме
Скелетные мышцы – главный резервуар карнитина – содержат его в концентрациях, в 20–500 раз превышающих концентрацию плазматического карнитина, которая колеблется от 41 (у женщин) до 50 (у мужчин) мкМ/л [20]. Соответствующие диеты, мясо и пищевые добавки представляют собой дополнительный источник карнитина. Но даже на диете, в значительной степени бедной карнитином, здоровый человек способен синтезировать его в достаточных количествах. Таким образом, карнитин не может считаться витамином, несмотря на свою витаминоподобную структуру.
Карнитин выводится из организма только с мочой и желчью, где, как было показано, аккумулируются ацильные производные. Ежедневные потери карнитина минимальны (< 60 мг/сут.), и на безкарнитиновой вегетарианской диете снижаются до 20 мг/сут.
Роль карнитина в метаболизме жиров
L-карнитин играет важную роль в метаболизме жиров. Во время сна, отдыха или средней физической нагрузки жирные кислоты представляют собой до 80 % от общих источников энергии. Лучше всего описана роль карнитина – как кофактора соответствующих ацилтрансфераз – в транспорте жирных кислот через внутреннюю мембрану митохондрий. В отсутствии L-карнитина внутренняя мембрана митохондрий непроницаема для жирных кислот и их производных, таких как ацил-CoA. Проникнув через мембрану, в митохондриях эти соединения могут быть превращены в ацетил-CoA в ходе β-окисления. Карнитин играет решающую роль в поддержании отношения ацетил-CoA/CoA в клетке. Во время интенсивной физической нагрузки увеличивается скорость образования ацетил-CoA. Это, в свою очередь, подавляет активность пируватдегидрогеназного (PDH) комплекса [22], а образующийся ацетил-CoA превращается в молочную кислоту. Ацетил-CoA способен реагировать с карнитином, передавая последнему ацетильную группу:
ацетил-CoA + карнитин → ацетил-карнитин + CoA
Таким образом, карнитин подавляет накопление молочной кислоты, увеличивая работоспособность в условиях интенсивных физических нагрузок. Это было показано в нескольких независимых исследованиях, результаты которых приведены в таблице 1.
Предварительные данные, полученные на ВИЧ-инфицированных пациентах, получавших препараты нуклеотидных аналогов (приводящие к увеличению содержания молочной кислоты в плазме крови), показали, что L-карнитин может оказаться полезным для пациентов с молочным ацидозом, когда уровень лактата в крови поднимается выше 10 мМ/л [23]. Sweeney et al. показали, что применение карнитина может приводить к увеличению концентрации тромбоцитов благодаря нормализации pH, снижению потребления глюкозы и подавлению образования молочной кислоты [24].
Скелетные мышцы раньше рассматривались как основной источник образования лактата – главной причины утомления – в условиях недостатка кислорода, расходуемого в ходе сокращения. Сейчас ясно, что скелетные мышцы играют важную роль не только в процессе образования лактата, но и участвуют в его утилизации. Это привлекло дополнительный интерес к судьбе молочной кислоты в мышцах и других тканях. Использование радиоактивно меченного лактата позволило установить, что мышцы активно извлекают лактат из кровяного русла и обладают значительными способностями к его последующему окислению [25, 26].
Сохранение пула митохондриального коэнзима A
В некоторых физиологических условиях, таких как физическая нагрузка, ишемия, острый стресс, увеличение активности пируватдегидрогеназного комплекса и интенсивности расщепления жиров, может повыситься уровень окисления ацетил-CoA, что, в свою очередь, приведет к накоплению ацетил-CoA и коротких ацильных производных CoA, представляющих собой главным образом продукты распада аминокислот с разветвленной боковой цепью. Отношение ацетил-CoA/CoA – важный фактор, регулирующий окисление пирувата, α-кетоглутарата и жирных кислот.
Окисление всех этих соединений происходит в присутствии свободного митохондриального CoA, выступающего в качестве акцептора ацетильных групп [27]. На этой стадии может происходить перенос избытка ацильных групп на карнитин с образованием ацилкарнитина под действием карнитинацетилтрансферазы. Кроме того, карнитин способен образовывать эфиры с разнообразными ацильными производными [20]. Статус ацитилирования подвижного карнитинового пула связан с доступностью и локализацией CoA. Карнитинтранслоказы могут осуществлять перенос избытка ацильных групп из митохондрий в цитоплазму в виде карнитиновых эфиров. Таким образом, освобождается CoA в матриксе митохондрий. Цитоплазматическая концентрация карнитина приблизительно в 100 раз выше концентрации свободного CoA (2,5 и 0,014 мМ соответственно). Важно, что равновесие между содержанием карнитиновых эфиров в тканях и свободным карнитином в плазме устанавливается быстро [28–30]. В сравнении со свободным карнитином карнитиновые эфиры быстрее выходят в плазму (вероятно, через OCTN1) и эффективнее экскрегируются почками.
Необходимо отметить, что равновесие между тканевым и плазматическим ацилкарнитином устанавливается в мышцах медленнее, чем в печени и почках.
Вопрос, могут ли клетки осуществлять перенос ацетильных групп между CoA и карнитином в цитоплазме, представляет особый интерес, потому что имеет отношение к судьбе ацетилкарнитина, освобождаемого из митохондрий и пероксисом, и к возможности взаимодействия митохондриального и цитоплазматического CoA через ацетилкарнитин.
Система приобретает дополнительную буферную емкость – когда скорость образования ацетил-CoA высока, циркулирующий ацетилкарнитин может осуществлять пополнение пула ацетил-CoA где-либо [27, 31].
Карнитин как фактор снижения веса
Причина использования карнитина в качестве агента снижения веса состоит в допущении, что регулярное употребление этого соединения приведет к увеличению его внутриклеточной концентрации. А это, в свою очередь, усилит окисление жиров и постепенно приведет к сокращению запасов жиров в организме.
Некоторое количество исследований показало, что употребление карнитина (до 6 г/сут. в течение 14 дней) не привело к изменению концентрации карнитина в мышцах здоровых людей без отклонений веса от нормы и не привело к снижению веса [32, 33]. Это не согласуется с данными, полученными в ходе клинических исследований, в которых карнитин употребляли тучные пациенты [34]. Недавно было показано, что подавление функции карнитинпальмитоилтрансферазы-1 в гипоталамусе снижает потребление пищи [35]. Регулярный прием карнитина стимулирует транскрипцию и активацию пальмитоилтрансферазы, вероятно, этим объясняется усиление аппетита, сопутствующее приему препарата [36]. Таким образом, утверждение, что прием карнитина приводит к снижению веса у здоровых людей, является недостаточно обоснованным и требует дальнейших исследований.
Недавние публикации демонстрируют способность L-карнитина (в высоких дозах) модулировать функцию глюкокортикоидного рецептора и, таким образом, замещать некоторые биологические активности глюкокортикоидов, которые, в частности, индуцируют липолиз в жировой ткани [37]. В случае гиперлипидемии регулярный прием карнитина вызвал усиление функции карнитинацилтрансферазы в клетках крови [38]. Для полноты изложения следует отметить, что этомоксир (etomoxir), ингибитор карнитинпальмитоилтрансферазы, подавляет системное окисление жиров, а антиконвульсанты, вызывающие карнитиновый дефицит, приводят к увеличению веса [39, 40].
Нагрузки
Карнитин необходим для нормального функционирования мышц по трем причинам. Во-первых, L-карнитин необходим для окисления жирных кислот. Во-вторых, он участвует в удалении избытка ацильных групп из митохондрий. В-третьих, он играет важную роль в процессах детоксикации. Мышцы требуют выполнения всех трех функций во время максимальной нагрузки. Теоретически, доступность карнитина может служить лимитирующим фактором для окисления жирных кислот и удаления ацетил-CoA во время нагрузки. Если это так, дополнительные порции карнитина могут увеличить производительность здоровых людей.
Результаты, представленные в таблице 1, показывают, что усиление мышечной функции и/или уменьшение времени восстановления наблюдалось в большинстве случаев регулярного употребления карнитина (305 случаев в 14 исследованиях), в то время как положительный эффект не был обнаружен в 70 случаях (в 7 исследованиях). Результаты исследований, в которых не было обнаружено воздействия регулярного приема карнитина на потребление кислорода и дыхательный коэффициент у здоровых спортсменов, вероятно, можно объяснить индивидуальными отличиями.
Различные методологические подходы также могут приводить к различным результатам. Так, наблюдение, что применение L-карнитина не оказывает влияния на содержание карнитина в мышцах [32, 41] противоречит обнаруженной стимуляции энзиматической активности в мышечных клетках при регулярном приеме карнитина [42, 43]. Эти данные демонстрируют усиление липидного метаболизма, ассоциированное со снижением дыхательного коэффициента [44–46], что, вероятно, объясняется способностью карнитина стимулировать мышечный метаболизм без изменения стационарной концентрации карнитина. Уменьшение содержания лактата в плазме крови, вызываемое приемом L-карнитина, может объясняться модуляцией активности ферментов в клетках крови, обладающих активным карнитиновым метаболизмом.
Защитные эффекты L-карнитина в клетках крови, особенно в тромбоцитах, известных своим участием в заживлении ран, могут объяснить участие L-карнитина в предотвращении мышечных повреждений в ходе тренировок [47, 48]. Кроме того, клинические исследования продемонстрировали эффективность применения карнитина при заболеваниях сердца.
Препараты, нарушающие функцию OCTN2, высокоафинного транспортера L-карнитина и диализ, могут снизить содержание карнитина в мышечной ткани у пациентов с почечными расстройствами, так как низкоафинный транспортер OCTN1 для адекватной работы требует более высоких концентраций карнитина. Предварительные данные выявили благотворные эффекты приема карнитина на мышечную функцию и работоспособность пациентов с почечными расстройствами.
Приблизительно 80 % мышечного карнитинового пула представлено в виде L-карнитина, 20 % – ацильным производным с коротким радикалом, 5 % – ацильным производным с длинным радикалом. Во время умеренной физической нагрузки (ниже лактатного порога) значительных изменений карнитинового пула в мышцах не наблюдается.
После 10 минут интенсивной нагрузки мышечный карнитиновый пул претерпевает значительные изменения в сторону увеличения доли ацильных производных с коротким радикалом (представленных преимущественно ацетилкарнитином) до 75 %.
Такое перераспределение карнитинового пула постепенно нормализуется после нагрузки, но это требует значительного времени – более 1 часа после 30 минут упражнений. Несмотря на такие драматические флуктуации карнитинового пула в мышцах, физические нагрузки оказывают незначительное влияние на состояние карнитина в плазме [52]. Прежние наблюдения о том, что степень и глубина изменений метаболизма карнитина зависят от объема и интенсивности работы, подтверждают тот факт, что эффект карнитина усиливается с увеличением нагрузки.
Упражнения на выносливость
Мнение о способности карнитина увеличивать выносливость основывается на трех гипотезах.
Первая заключается в том, что концентрация карнитина в мышцах может быть слишком низкой, чтобы позволить карнитинацилтрансферазам функционировать на высоком уровне и поддерживать, таким образом, высокий уровень окисления жиров во время выполнения упражнений. Согласно второй гипотезе, прием карнитина способен увеличить его концентрацию в мышцах. Третья гипотеза состоит в том, что увеличение концентрации карнитина в мышцах приведет к усилению окисления жиров и позволит сохранить клеточный гликоген. Имеются экспериментальные данные, подтверждающие эти гипотезы.
Интенсивные упражнения
В ходе интенсивных физических упражнений содержание свободного карнитина в мышцах падает в результате взаимодействия с ацетил-CoA. Падение концентрации карнитина коррелирует с интенсивностью нагрузки и может быть весьма значительным (до 0,5 мМ/кг влажного веса мышцы). Это падение концентрации рассматривается как один из механизмов снижения окисления жирных кислот в плазме и триацилглицеролов в мышцах во время интенсивной нагрузки [53].
Большинство исследований показало большее или меньшее увеличение производительности мышц и потребления кислорода у профессиональных и непрофессиональных спортсменов, особенно при длительном применении карнитина в высоких дозах [7, 42–46, 54–61]. В то же время некоторые исследователи не обнаружили влияния L-карнитина на потребление кислорода, производительность мышц и аккумуляцию лактата у здоровых спортсменов [32, 41, 62–67].
В процессе интенсивной тренировки развивается гипоксия, увеличивающая концентрацию аммония в крови [68]. Будет ли наблюдаться корреляция между концентрациями лактата и аммония в плазме, зависит от интенсивности нагрузки [69]. Упражнения в условиях гипоксии стимулируют транспорт глюкозы в мышцы и снижение концентрации свободного карнитина [70]. У крыс, использованных в качестве модельного объекта, введение карнитина предотвращало токсические эффекты накопления аммония в ходе физических нагрузок на трех уровнях: 1) активация ферментов цикла мочевины, 2) взаимодействие с рецепторами глутамата, 3) снижение уровня свободных радикалов. Мышечные боли и накопление креатинкиназы в течение восстановительного периода были ослаблены в результате приема карнитина (3 г/сут. в течение трех недель). Эти данные показывают, что карнитин способен оказывать благотворное влияние на процесс восстановления после нагрузки [54]. В некоторых работах изучалась роль карнитина в условиях острого мышечного стресса и его влияние на процессы восстановления. Ряд биохимических маркеров указывает, что карнитин снижает катаболизм пуринов, образование свободных радикалов и ощущение болезненности [48].
Иммунная система
Продолжительные и интенсивные тренировки зачастую ассоциированы с супрессией иммунной системы и увеличением риска инфекционных заболеваний. Хотя высококвалифицированные спортсмены не могут быть описаны в терминах клинического иммунодефицита, существуют свидетельства того, что некоторые параметры иммунной активности подавляются длительными и интенсивными тренировками. Среди них – функция нейтрофилов, концентрация иммуноглобулинов в плазме и слюне, активность нормальных киллеров, уровень секреции иммуноглобулина А, активность макрофагов.
Кроме того, значительно возрастает риск респираторных заболеваний [72–6]. Это позволяет говорить о клинически релевантном подавлении иммунной системы. Проблема осложняется эффектом диеты, так как режим питания способен влиять на иммунный ответ. Увеличение содержания жиров в рационе приводит к увеличению выносливости и не оказывает неблагоприятного воздействия на уровень воспалительных цитокинов (интерлейкин-1β, интерлейкин-6, TNFα) у бегунов [77, 78].
Поглощение L-карнитина клетками крови
Увеличение концентрации карнитина в лимфоцитах и гранулоцитах может отражать усиление их метаболического состояния в процессе синтеза иммуноглобулина или фагоцитоза. Поглощение L-карнитина гранулоцитами было описано при воспалительных процессах, вызванных множественными травмами и бактериальными инфекциями. У пациентов с синдромом Крона (хронический воспалительный процесс) увеличено содержание карнитина в T-лимфоцитах [79, 80]. Было показано, что периферические одноядерные клетки крови пациентов с приобретенным иммунодефицитом и нормальным плазматическим уровнем карнитина демонстрируют карнитиновое истощение. Плазматическая концентрация карнитина не всегда прямо отражает внутриклеточную.
У пациентов с приобретенным иммунодефицитом, принимавших высокие дозы карнитина (6 г/сут.) в течение 2 недель, была выявлена тенденция к восстановлению внутриклеточной концентрации карнитина [82].
Принимая во внимание сходные механизмы реактивации вируса Эпштейн-Барр у пациентов с приобретенным иммунодефицитом и профессиональных спортсменов [72], такие фармакологические дозы карнитина могут быть рекомендованы спортсменам. Предварительные данные свидетельствуют в пользу того, что L-карнитин способен защищать периферические лимфоциты от оксидативного стресса [83]. Учитывая возрастное снижение активности карнитин-
ацилтрансфераз, прием карнитина может оказаться полезным не только для спортсменов.
Стимуляция гематопоэза
Работа Matsumura et al. была призвана установить взаимосвязь между L-карнитином, стволовыми клетками и предшественниками клеток крови [84]. Препарат оказался способен стимулировать эритропоэз. Исследования, проведенные с участием пациентов с почечной анемией, подтвердили положительное влияние карнитина на эритропоэз [85]. Таким образом, прием L-карнитина может оказаться полезным при тренировках в высокогорных условиях.
Подавление апоптоза в клетках иммунной системы
Передача сигнала через поверхностный гликопротеин Fas, также известный как CD95, представляет собой один из важнейших путей регуляции программируемой клеточной смерти (апоптоза). L-карнитин подавляет апоптоз, взаимодействия с лигандом и рецептором [86]. Передача сигнала через Fas-рецептор активирует кислую сфингомиелиназу, приводящую к освобождению керамида. Добавление L-карнитина ингибирует сфингомиелиназу, функции каспаз 3, 7, 8 и предотвращает изменение проводимости митохондриальной мембраны [10, 87]. Другой антиапоптотический механизм с участием карнитина был обнаружен в Т-лимфоцитах, где добавление карнитина стимулировало увеличение уровня IGF1. IGF1 подавляет димеризацию проапоптотических генов BCL2-BAX в митохондриальной мембране [89]. Кроме того, это приводит к подавлению транскрипции BCL2-респонсивных генов.
Вопросы безопасности применения L-карнитина
Не всегда просто провести классификацию соединения, которое, с одной стороны, синтезируется в организме, а с другой – усваивается из пищи. L-карнитин принадлежит к группе веществ, которые продаются в качестве пищевой добавки, но могут быть описаны как лекарственные препараты.
За исключением пациентов на диализе, клинические дозы карнитина колеблются в пределах от 50 до 350 мг на кг веса тела. Экстраполяция этих доз позволяет говорить о 3 г/сут. для пациента весом 70 кг как о минимальной дозе, позволяющей ожидать фармакологический эффект. Прием карнитина в количестве, превышающем 4 г/сут., может вызвать легкий стресс пищеварительного тракта [33]. Доза в 2 г/сут. представляется достаточной для восстановления [47, 48].
Заключение
Польза от применения карнитина во время интенсивной физической нагрузки была показана более чем в 300 исследованиях. Однако индивидуальные отличия в отклике на прием препарата должны быть приняты в расчет. L-карнитин способен ослаблять побочные эффекты интенсивных тренировок, снижая уровень гипоксии. L-карнитин способствует восстановлению мышц и активизирует иммунную систему.
Новые данные о влиянии карнитина на тиреоидную и стероидную сигнализацию представляют основу для дальнейших исследований системных эффектов применения L-карнитина спортсменами.
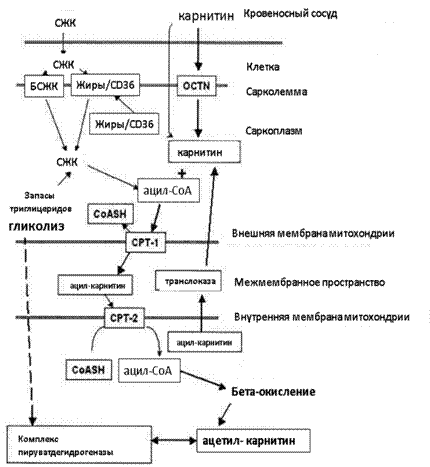
Рис. 1. Роль L-карнитина в окислительном метаболизме
Первичная функция L-карнитина состоит в переносе жирных кислот (СЖК) в митохондрии с помощью CPT-1 (карнитинпальмитоилтрансферазы). Вторичная функция карнитина состоит в поддержании отношения ацетил-CoA/CoA. Карнитин способен реагировать с избытком образующихся в течение физической нагрузки ацетильных групп, освобождая CoA. В результате происходит снижение отношения ацетил-CoA/CoA, что, в свою очередь, приводит к активации PDH. PDH катализирует превращение пирувата в ацетил-CoA, предотвращая образование вызывающей симптомы утомления молочной кислоты.
Карнитин попадает в клетку вместе с переносчиком органических катионов (OCTN).
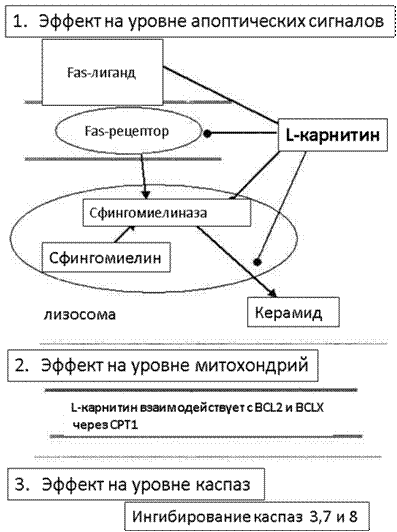
Рис. 2. Биохимические и физиологические изменения в ходе апоптоза
На уровне передачи сигнала карнитин предотвращает образование керамида путем прямого ингибирования сфингомиелиназы. Взаимодействие карнитинпальмитоилтрансферазы-1 c проапоптотическими белками семейства BCL оказывает антиапоптотический эффект на уровне митохондрий, предотвращая изменение проницаемости внутренней мембраны и выход цитохрома С. Кроме того, карнитин способен ингибировать каспазы 3, 7, 8.
Таблица 1
Доза | Результаты | Число пациентов (n) | Источник |
1 г эндогенно перед тренировкой | Увеличение работоспособности | 17 | Dragan et al.56 |
2 г перед интенсивной тренировкой | Стимуляция активности PDH, снижение уровня плазматического лактата plasma lactate and pyruvate | 10 | Siliprandi et al.7 |
2 г перед интенсивной тренировкой | Увеличение Vo2max | 10 | Vecchiet et al.58 |
2 г/сут. в течение 4 недель | Усиление активности дыхательной цепи в мышцах | 14 | Huertas et al.43 |
2 г/сут. в течение 4 недель | Стимуляция активности PDH, увеличение Vo2max у бегунов на длинные дистанции | 16 | Arenas et al.42 |
4 г/сут. в течение 2 недель | Увеличение Vo2max | 6 | Marconi et al.60 |
6 г + введение глюкозы | Снижение дыхательного коэффициента | 47 | Angelini et al.61 |
2 г/сут. в течение 28 дней | Снижение дыхательного коэффициента | 10 | Gorostiaga et al.45 |
3 г/сут. в течение 7 дней | Снижение дыхательного коэффициента | 7 | Wyss et al.44 |
3 г/сут. в течение 10 дней | Усиление активности дыхательной цепи | 10 | Muller et al.46 |
1 г/сут. в течение 3 недель | Увеличение работоспособности | 110 | Dragan et al.55 |
1 г/сут. в течение 6 недель + 2 г/сут. в течение 10 дней перед соревнованиями | Увеличение работоспособности | 7 | Dragan et al.57 |
1 г/сут. в течение 6 недель | Положительный эффект на восстановление | 24 | Arenas et al.59 |
3 г/сут. в течение 3 недель | Увеличение работоспособности | 6 | Giamberardino et al.54 |
2 г/сут. в течение 3 недель | Увеличение работоспособности | 10 | Kraemer et al.47 |
Всего человек | 305 | ||
2 г перед стартом | Отсутствует увеличение работоспособности | 7 | Colombani et al.62 |
1 г перед стартом | Отсутствует увеличение работоспособности | 9 | Nuesch et al.63 |
4 г/сут. в течение 14 дней | Нет эффекта на образование лактата | 8 | Barnett et al.33 |
3 г/сут. в течение 7 дней | Нет изменения дыхательного коэффициента | 9 | Decombaz et al.65 |
5 г/сут. в течение 5 дней | Отсутствует увеличение работоспособности | 7 | Soop et al.66 |
2 г/сут. в течение 4 недель | Нет эффекта на Vo2max | 10 | Oyono-Enguelle et al.67 |
2 г/сут. в течение 7 дней | Нет эффекта на Vo2max | 20 | Trappe et al.64 |
Всего человек | 70 |
Литература
1. Cerretelli P., Marconi C. L-carnitine supplementation in humans. The effects on physical performance. Int J Sports Med 1990; 11: 1.
2. Rebouche C. J., Chenard C. A. Metabolic fate of dietary carnitine in human adults: identification and quantification of urinary and fecal metabolites. J Nutr 1991; 121: 539.
3. Gulewitsch W. K.R. Zur Kenntnis der Extraktionsstoffe der Muskeln. Mitteilungen über das Carnitin (extracted substances in muscle, report on carnitine). Hoppe-Seyler Z Physiol Chem 1905;45:326.
4. Strack E., Neubauer E., Geissendörfer H. Über den Gehalt von Cholin, Acetylcholinund Carnitin im Muskel (content of choline acetylcholine and carnitine in muscle). Hoppe-Seyler Z Physiol Chem 1935; 233: 189.
5. Fritz I. B. Action of carnitine on long chain fatty acid oxidation by liver. Am J Physiol 1959; 197: 297.
6. Lopes G., Bazotte R. B., Curi R., Alves-Do-Prado WL - and DL-carnitine induce tetanic fade in rat neuromuscular preparation. Braz J Med Biol Res 2003; 36: 1255.
7. Siliprandi N., Di Lisa F., Menabo R. Clinical use of carnitine. Past, present and future. Adv Exp Med Biol 1990; 272: 175.
8. Brevetti G., Chiariello M., Ferulano G., et al. Increases in walking distance in patients with peripheral vascular disease treated with L-carnitine: a double-blind, cross-over study. Circulation 1988; 77: 767.
9. Vescovo G., Ravara B., Gobbo V. et al. L-carnitine: a potential treatment for blocking apoptosis and preventing skeletal muscle myopathy in heart failure. Am J Physiol Cell Physiol 2002; 283: C802.
10. Mutomba MC, Yuan H, Konyavko M. et al. Regulation of the activity of caspases by L-carnitine and palmitoylcarnitine. FEBS Lett 2000; 478: 19.
11. Di Marzio L., Alesse E., Roncaioli P. et al. Influence of L-carnitine on CD95 cross-lining-induced apoptosis and ceramide generation in human cell lines: correlation with its effects on purified acidic and neutral sphingomyelinases in vitro. Proc Assoc Am Phys 1997; 109: 154.
12. Karlic H., Lohninger A., Laschan C. et al. Downregulation of carnitine acyltransferases and organic cation transporter OCTN2 in mononuclear cells in healthy elderly and patients with myelodysplastic syndromes. J Mol Med 2003; 81: 435.
13. Brown H. R., Ni H., Benavides G. et al. Correlation of simultaneous differential gene expression in the blood and heart with known mechanisms of adriamycininduced cardiomyopathy in the rat. Toxicol Pathol 2002; 30: 452.
14. Matera M., Bellinghieri G., Costantino G., Santoro D., Calvani M., Savica V. History of L-carnitine: implications for renal disease. J Ren Nutr 2003; 13: 2.
15. Schoderbeck M., Auer B., Legenstein E. et al. Pregnancy-related changes of carnitine and acylcarnitine concentrations of plasma and erythrocytes. J Perinat Med 1995; 23: 477.
16. Bremer J. Carnitine-metabolism and functions. Physiol Rev 1983; 63: 1420.
17. Bieber L. L. Carnitine. Annu Rev Biochem 1988; 57: 261.
18. Berardi S., Stieger B., Wachter S., O’Neill B., Krahenbuhl S. Characterization of a sodium-dependent transport system for butyrobetaine into rat liver plasma membrane vesicles. Hepatology 1998; 28: 521.
19. Vaz F. M., Wanders R. J. Carnitine biosynthesis in mammals. Biochem J 2002; 361: 417.
20. Ramsay R. R., Gandour R. D., van der Leij F. R. Molecular enzymology of carnitine transfer and transport. Biochim Biophys Acta 2001; 1546: 21.
21. McGarry J. D., Brown N. F. The mitochondrial carnitine palmitoyltransferase system. From concept to molecular analysis. Eur J Biochem 1997; 244: 1.
22. Jeukendrup A. E. Regulation of fat metabolism in skeletal muscle. Ann NY Acad Sci 2002; 967: 217.
23. Claessens Y. E., Cariou A., Monchi M. et al. Detecting life-threatening lactic acidosis related to nucleoside-analog treatment of human immunodeficiency virus-infected patients, and treatment with L-carnitine. Crit Care Med 2003; 31: 1042.
24. Sweeney J. D., Blair A. J., Cheves T. A., Dottori S., Arduini A. L-carnitine decreases glycolysis in liquid-stored platelets. Transfusion 2000; 40: 1313.
25. Van Hall G. Lactate as a fuel for mitochondrial respiration. Acta Physiol Scand 2000; 168: 643.
26. Van Hall G., Gonzalez-Alonso J., Sacchetti M., Saltin B. Skeletal muscle substrate metabolism during exercise: methodological considerations. Proc Nutr Soc 1999; 58: 899.
27. Bakker A., Biermans W., Van Belle H., De Bie M., Bernaert I., Jacob W. Ultrastructural localisation of carnitine acetyltransferase activity in mitochondria of rat myocardium. Biochim Biophys Acta 1994; 1185: 97.
28. Faergeman N. J., Knudsen J. Role of long-chain fatty acyl-CoA esters in the regulation of metabolism and in cell signalling. Biochem J 1997; 323(pt 1): 1.
29. Idell-Wenger J. A., Grotyohann L. W., Neely J. R. Regulation of fatty acid utilization in heart. Role of the carnitine-acetyl-CoA transferase and carnitine-acetyl carnitine translocase system. J Mol Cell Cardiol 1982; 14: 413.
30. Lysiak W., Lilly K., DiLisa F., Toth P. P., Bieber L. L. Quantitation of the effect of L-carnitine on the levels of acid-soluble short-chain acyl-CoA and CoASH in rat heart and liver mitochondria. J Biol Chem 1988; 263: 1151.
31. Abbas A. S., Wu G., Schulz H. Carnitine acetyltransferase is not a cytosolic enzyme in rat heart and therefore cannot function in the energy-linked regulation of cardiac fatty acid oxidation. J Mol Cell Cardiol 1998; 30: 1305.
32. Barnett C., Costill D. L., Vukovich M. D. et al. Effect of L-carnitine supplementation on muscle and blood carnitine content and lactate accumulation during highintensity sprint cycling. Int J Sport Nutr 1994; 4: 280.
33. Villani R. G., Gannon J., Self M., Rich P. A. L-carnitine supplementation combined with aerobic training does not promote weight loss in moderately obese women. Int J Sport Nutr Exerc Metab 2000; 10: 199.
34. Walter P., Schaffhauser A. O. L-carnitine, a vitamin-like substance for functional food. Proceedings of the symposium on L-carnitine, April 28 to May 1, 2000, Zermatt, Switzerland. Ann Nutr Metab 2000; 44: 75.
35. Obici S., Feng Z., Arduini A., Conti R., Rossetti L. Inhibition of hypothalamic carnitine palmitoyltransferase-1 decreases food intake and glucose production. Nat Med 2003; 9: 756.
36. Karlic H., Lohninger S., Koeck T., Lohninger A. Dietary L-carnitine stimulates carnitine acyltransferases in the liver of aged rats. J Histochem Cytochem 2002; 50: 205.
37. Alesci S., De Martino M. U., Mirani M. et al. L-carnitine: a nutritional modulator of glucocorticoid receptor functions. FASEB J 2003; 17: 1553.
38. Lohninger A., Agu C. A., Hofbauer R., Nissel H., Karlic H. Carnitine and transcription of carnitine palmitoyltransferases – in vitro and in vivo studies. In: Richter V, Reuter W., Rassoul F., Thiery J., eds. Lipoproteinmetabolismus und Atherosklerosepraevention. Leipzig: Wissenschaftliche Skripten, Zwickau, 2002: 113.
39. Hinderling V. B., Schrauwen P., Langhans W., Westerterp-Plantenga M. S. The effect of etomoxir on 24-h substrate oxidation and satiety in humans. Am J Clin Nutr 2002; 76: 141.
40. Jallon P., Picard F. Bodyweight gain and anticonvulsants: a comparative review. Drug Saf 2001; 24: 969.
41. Vukovich M. D., Costill D. L., Fink W. J. Carnitine supplementation: effect on muscle carnitine and glycogen content during exercise. Med Sci Sports Exerc 1994; 26: 1122.
42. Arenas J., Huertas R., Campos Y., Diaz A. E., Villalon J. M., Vilas E. Effects of L-carnitine on the pyruvate dehydrogenase complex and carnitine palmitoyl transferase activities in muscle of endurance athletes. FEBS Lett 1994; 341: 91.
43. Huertas R., Campos Y., Diaz E. et al. Respiratory chain enzymes in muscle of endurance athletes: effect of L-carnitine. Biochem Biophys Res Commun 1992; 188: 102.
44. Wyss V., Ganzit G. P., Rienzi A. Effects of L-carnitine administration on VO2max and the aerobic-anaerobic threshold in normoxia and acute hypoxia. Eur J Appl Physiol Occup Physiol 1990; 60: 1.
45. Gorostiaga E. M., Maurer C. A., Eclache J. P. Decrease in respiratory quotient during exercise following L-carnitine supplementation. Int J Sports Med 1989; 10: 169.
46. Muller D. M., Seim H., Kiess W., Loster H., Richter T. Effects of oral L-carnitine supplementation on in vivo long-chain fatty acid oxidation in healthy adults. Metabolism 2002; 51: 1389.
47. Kraemer W. J., Volek J. S, French D. N. et al. The effects of L-carnitine L-tartrate supplementation on hormonal responses to resistance exercise and recovery. J Strength Cond Res 2003; 17: 455.
48. Volek J. S., Kraemer W. J., Rubin M. R., Gomez A. L., Ratamess N. A., Gaynor P. L-carnitine L-tartrate supplementation favorably affects markers of recovery from exercise stress. Am J Physiol Endocrinol Metab 2002; 282: E474.
49. Brass E. P., Hiatt W. R. Carnitine metabolism during exercise. Life Sci 1994; 54: 1383.
50. Brass E. P. Supplemental carnitine and exercise. Am J Clin Nutr 2000; 72: 618S.
51. Romijn J. A., Coyle E. F., Sidossis L. S. et al. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am J Physiol 1993; 265: E380.
52. Hiatt W. R., Regensteiner J. G., Wolfel E. E., Ruff L., Brass E. P. Carnitine and acylcarnitine metabolism during exercise in humans. Dependence on skeletal muscle metabolic state. J Clin Invest 1989; 84: 1167.
53. Brass E. P., Hiatt W. R. The role of carnitine and carnitine supplementation during exercise in man and in individuals with special needs. J Am Coll Nutr 1998; 17: 207.
54. Giamberardino M. A., Dragani L., Valente R., Di Lisa F., Saggini R., Vecchiet L. Effects of prolonged L-carnitine administration on delayed muscle pain and CK release after eccentric effort. Int J Sports Med 1996; 17: 320.
55. Dragan IG., Vasiliu A., Georgescu E., Eremia N. Studies concerning chronic and acute effects of L-carnitine in elite athletes. Physiologie 1989; 26: 111.
56. Dragan A. M., Vasiliu D., Eremia N. M., Georgescu E. Studies concerning some acute biological changes after endovenous administration of 1 g L-carnitine, in elite athletes. Physiologie 1987; 24: 231.
57. Dragan G. I., Wagner W., Ploesteanu E. Studies concerning the ergogenic value of protein supply and L-carnitine in elite junior cyclists. Physiologie 1988; 25: 129.
58. Vecchiet L., Di Lisa F., Pieralisi G. et al. Influence of L-carnitine administration on maximal physical exercise. Eur J Appl Physiol Occup Physiol 1990; 61: 486.
59. Arenas J., Ricoy J. R., Encinas A. R. et al. Carnitine in muscle, serum, and urine of nonprofessional athletes: effects of physical exercise, training, and L-carnitine administration. Muscle Nerve 1991; 14: 598.
60. Marconi C., Sassi G., Carpinelli A., Cerretelli P. Effects of L-carnitine loading on the aerobic and anaerobic performance of endurance athletes. Eur J Appl Physiol Occup Physiol 1985; 54: 131.
61. Angelini A., Imparato L., Landi C., Porfido F. A., Ciarimboli M., Marro A. Variation in levels of glycaemia and insulin after infusion of glucose solutions with or without added L-carnitine. Drugs Exp Clin Res 1993; 19: 219.
62. Colombani P., Wenk C., Kunz I. et al. Effects of L-carnitine supplementation on physical performance and energy metabolism of endurance-trained athletes: a double-blind crossover field study. Eur J Appl Physiol Occup Physiol 1996; 73: 434.
63. Nuesch R., Rossetto M., Martina B. Plasma and urine carnitine concentrations in well-trained athletes at rest and after exercise. Influence of L-carnitine intake. Drugs Exp Clin Res 1999; 25: 167.
64. Trappe S. W., Costill D. L., Goodpaster B., Vukovich M. D., Fink W. J. The effects of L-carnitine supplementation on performance during interval swimming. Int J Sports Med 1994; 15: 181.
65. Decombaz J., Deriaz O., Acheson K., Gmuender B., Jequier E. Effect of L-carnitine on submaximal exercise metabolism after depletion of muscle glycogen. Med Sci Sports Exerc 1993; 25: 733.
66. Soop M., Bjorkman O., Cederblad G., Hagenfeldt L., Wahren J. Influence of carnitine supplementation on muscle substrate and carnitine metabolism during exercise. J Appl Physiol 1988; 64: 2394.
67. Oyono-Enguelle S., Freund H., Ott C. et al. Prolonged submaximal exercise and L-carnitine in humans. Eur J Appl Physiol Occup Physiol 1988; 58: 53.
68. Casas H., Murtra B., Casas M. et al. Increased blood ammonia in hypoxia during exercise in humans. J Physiol Biochem 2001; 57: 303.
69. Yuan Y., So R., Wong S., Chan K. M. Ammonia threshold-comparison to lactate threshold, correlation to other physiological parameters and response to training. Scand J Med Sci Sports 2002; 12: 358.
70. Parolin M. L., Spriet L. L., Hultman E., Hollidge-Horvat M. G., Jones N. L., Heigenhauser G. J. Regulation of glycogen phosphorylase and PDH during exercise in human skeletal muscle during hypoxia. Am J Physiol Endocrinol Metab 2000; 278: E522.
71. Gani F., Passalacqua G., Senna G., Mosca Frezet M. Sport, immune system and respiratory infections. Allerg Immunol (Paris) 2003; 35: 41.
72. Gleeson M., Pyne D. B., Austin J. P. et al. Epstein-Barr virus reactivation and upper-respiratory illness in elite swimmers. Med Sci Sports Exerc 2002; 34: 411.
73. Gleeson M., McDonald W. A., Pyne D. B. et al. Immune status and respiratory illness for elite swimmers during a 12-week training cycle. Int J Sports Med 2000; 21: 302.
74. Gleeson M. Interleukins and exercise. J Physiol 2000; 529(pt 1): 1.
75. Gleeson M. Mucosal immune responses and risk of respiratory illness in elite athletes. Exerc Immunol Rev 2000; 6: 5.
76. Gleeson M., Bishop N. C. Special feature for the Olympics: effects of exercise on the immune system: modification of immune responses to exercise by carbohydrate, glutamine and anti-oxidant supplements. Immunol Cell Biol 2000; 78: 554.
77. Venkatraman J. T., Feng X., Pendergast D. Effects of dietary fat and endurance exercise on plasma cortisol, prostaglandin E2, interferon-gamma and lipid peroxides in runners. J Am Coll Nutr 2001; 20: 529.
78. Venkatraman J. T., Pendergast D. Effects of the level of dietary fat intake and endurance exercise on plasma cytokines in runners. Med Sci Sports Exerc 1998; 30: 1198.
79. Adlouni H. A., Katrib K., Ferard G. Changes in carnitine in polymorphonuclear leukocytes, mononuclear cells, and plasma from patients with inflammatory disorders. Clin Chem 1988; 34: 40.
80. Demirkol M., Sewell A. C., Bohles H. The variation of carnitine content in human blood cells during disease – a study in bacterial infection and inflammatory bowel disease. Eur J Pediatr 1994; 153: 565.
81. De Simone C., Famularo G., Tzantzoglou S., Trinchieri V., Moretti S., Sorice F. Carnitine depletion in peripheral blood mononuclear cells from patients with AIDS: effect of oral L-carnitine. AIDS 1994; 8: 655.
82. De Simone C., Tzantzoglou S., Famularo G. et al. High dose L-carnitine improves immunologic and metabolic parameters in AIDS patients. Immunopharmacol Immunotoxicol 1993; 15: 1.
83. Franceschi C. Cell proliferation, cell death and aging. Aging (Milano) 1989; 1: 3.
84. Matsumura M., Hatakeyama S., Koni I., Mabuchi H. Effect of L-carnitine and palmitoyl-L-carnitine on erythroid colony formation in fetal mouse liver cell culture. Am J Nephrol 1998; 18: 355.
85. Matsumoto Y., Amano I., Hirose S. et al. Effects of L-carnitine supplementation on renal anemia in poor responders to erythropoietin. Blood Purif 2001; 19: 24.
86. Moretti S., Alesse E., Di Marzio L. et al. Effect of L-carnitine on human immunodeficiency virus-1 infection-associated apoptosis: a pilot study. Blood 1998; 91: 3817.
87. Pastorino J. G., Snyder J. W., Serroni A., Hoek J. B., Farber J. L. Cyclosporin and carnitine prevent the anoxic death of cultured hepatocytes by inhibiting the mitochondrial permeability transition. J Biol Chem 1993; 268: 13791.
88. Di Marzio L., Moretti S., D’Alo S. et al. Acetyl-L-carnitine administration increases insulin-like growth factor 1 levels in asymptomatic HIV-1-infected subjects: correlation with its suppressive effect on lymphocyte apoptosis and ceramide generation. Clin Immunol 1999; 92: 103.
89. Wang L., Ma W., Markovich R., Chen J. W., Wang P. H. Regulation of cardiomyocyte apoptotic signaling by insulin-like growth factor I. Circ Res 1998; 83: 516
90. Pugazhenthi S., Miller E., Sable C. et al. Insulin-like growth factor-I induces bcl-2 promoter through the transcription factor cAMP-response element-binding protein. J Biol Chem 1999; 274: 27529.
91. Wolff J. M.D., Strack E. Toxicity of L(-)-carnitine and various 0-acylcarnitines (in German). Acta Biol Med Ger 1971; 26: 1237.
92. Sulkers E. J., Lafeber H. N., Degenhart H. J., Przyrembel H., Schlotzer E., Sauer P. J. Effects of high carnitine supplementation on substrate utilization in low-birthweight infants receiving total parenteral nutrition. Am J Clin Nutr 1990; 52: 889.
93. Li B., Lloyd M. L., Gudjonsson H., Shug A. L., Olsen WA. The effect of enteral carnitine administration in humans. Am J Clin Nutr 1992; 55: 838.
