
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
БИЛЕТ 9
 •Внутри и межмолекулярные силы, обеспечивающие формирование третичной и четвертичной
•Внутри и межмолекулярные силы, обеспечивающие формирование третичной и четвертичной
структуры белка. Функциональная роль третичной и четвертичной структуры белка.
Особенности строения и функционирования олигомерных белков на примере гемсодержащих
белков - гемоглобина и миоглобина. Представления о коопертивности и аллостерии.
ТРЕТИЧНАЯ структура. Фибриллярные белки. •α-кератин – 2 типа белков, но центр. участки одинаковые. Сам белок – coiled coil из 4 α-спиралей + попер. Cys-Cys-связи. Пример с хим. завивкой волос (перераспред. дисульфид. мостиков), мокрые волосы – просто перераспред. водор. связей. В составе волос, ногтей и т. д. •Фиброин и β-кератин – из антипараллельных β-складок (каждая вторая АК – Gly; складка идет «волнами», а бок. цепи АК выступают на пов-ти Þс одной стороны складки выступают только глицины, а с другой – преим. Ala и Ser Þсвязи Gly-Gly и Ala/Ser-Ala/Ser Þ↑прочности). В составе шелка, перьев. •Коллаген. 3 типа: I – кость, сухожилия, кожа; II – хрящи; III – стенки сосудов. Субъединица – тропоколлаген: каждая 3я АК – Gly, много Pro +необычн. АК – 4-гидроксипролин, 3-гидроксипролин, 5-гидроксилизин. !Pro и Lys подверг-ся гидроксил-ия после синтеза полипептидной цепи (посттрансляционные модификации). Тройная спираль Þоч. плотная упаковка (поэтому «полезна» маленькая АК – Gly). +доп. стабилизация за счет лизил-косидазы Þреакция между остатками Lys [Б9] Длина волокна – 300 нм, в местах соединения волокон друг с другом – промежуток 40 нм Þместо связывания с сахарами и! гидроксиапатитом Са5(РО4)3ОН.
Глобулярные белки (на примере мио - и гемоглобина). Есть гем – циклический тетрапиррол (Þкр. цвет): 4 молекулы пиррола +4 α-метиленовых мостика. [Б10] !NB: В отличие от цитохрома, хлорофилл-содерж. белков и пр. дряни, в мио - и гемоглобине окисление Fe2+ ведет к потере их биол. активности. •Миоглобин (красные мыщцы, при кисл. голодании отдает О2, не способен к транспортированию О2). На пов-ти – полярн. АК-остатки, внутри – неполярные. Если сильным понижение рН удалить гем Þпотеря трет. и втор. структуры. Снова ввели гем Þвсе восстановилось. Гем образ-ет плоскость (4 координационные положения), 5-ое (перп. ось) положение занято проксим. гистидином, 6-ое – свободно, но рядом «висит» дистальный гистидин. Неоксигенированный белок Þатом Fe выступает в сторону прокс. His (His F8) на 0,03 нм, оксигенир. белок Þна 0,01 Þпри насыщении О2 изм-ся и положение дист. His (His E7) ÞQконформации. Молекула О2 обр-ет с плоскостью гема угол 120о. Молекула угарного газа СО более прочно связ-ся, но выгодное для нее положение под углом 90о занять не может (мешает дист. His) Þтоже становится под углом Þ↓прочность связи Þ«ест. защита» от перенасыщения белка организма угарным газом. См. кривую насыщение белка О2 – при выс. парц. давлении О2 и Mb и Hb одинаково хорошо связ-ют газ, но при низк. значения парц. давл. (как в тканях) Hb проще его отдает ÞHb больше подходит для транпорта. •Гемоглобин. Есть четвертичная структура. Разные типы полипепт. цепей (α,β,γ,δ,S). Одна молекула – две цепи двух разных типов. Разн. цепи кодир-ся разными генами. ш: 1.HbA (норма, α2β2); 2.HbF (фетальный, α2γ2); 3.HbS (серповидно-клеточная анемия); 4.HbA2 (минорный гемоглобин взрослого человека, α2δ2). Аллостерические св-ва. Связ-ет 4 молекулы О2 на тетрамер, кривая насыщение сигмовидной формы. !NB: оксигенирование Hb Þразрыв концевых солевых связей (между N - и С-концами разных полипепт. цепей Þоблегчение послед. присоединения О2 Þповорот α/β-субъединиц друг отн. друга Þкомпактизация тетрамера ÞFe втягивается в плоскость гема. Част. оксигенир. структура – Т (taut – напряженая), полностью – R (relaxed). 2,3-бисфосфоглицерат стабилизирует Т-форму.
 •Окислительное декарбоксилирование пировиноградной кислоты. Пируватдегидрогеназный
•Окислительное декарбоксилирование пировиноградной кислоты. Пируватдегидрогеназный
комплекс. Строение, свойства и механизм функционирования и регуляции. Сопряжение
гликолиза с циклом Кребса. Пируват как ключевой метаболит в превращениях углеводов,
аминокислот и жирных кислот.
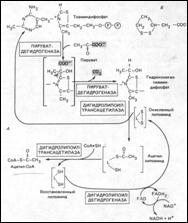
Окислительное декарбоксилирование пирувата является связующим звеном между двумя последовательными этапами окисления глюкозы в клетке – гликолизом и ЦК. Но, т. к. гликолиз протекает в цитозоле клетки, а ЦК в матриксе МХ, то пируват предварительно должен быть транспортирован внутрь МХ. Процесс обеспечивается специальными переносчиками, расположенными на внутренней мембране МХ, которые осуществляют симпорт П. вместе с протоном (наружную мембрану МХ пируват проходит через поры).Внутри МХ происходит ОДП и образование ацетил – КоА, состоящее из 4 последовательных реакций. Они катализируются тремя ферментами, которые работают в определенной последовательности и входят в состав одного большого ферментного комплекса – пируватдегидрогеназного. Поэтому промежуточные продукты способны достаточно быстро взаимодействовать друг с другом. Он аналогичен α – кетоглутаратдегидрогеназному. Комплекс состоит из: •Пируватдегидрогеназы (кофактором является тиаминпирофосфат = Тиаминдифосфат, ТРР); •Дигидролипоилтрансацетилазы (кофактор – липоевая кислота); •Дигидролипоилдегидрогеназы (кофактор - ФАД) = Е3.
!!! Также в состав комплекса - 2 регуляторных белка: протеинкиназа и фосфопротеин фосфатаза
Субъединицы дигидролипоилтрансферазы образуют его ядро, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
Последовательность превращений: 1. На первой и стадии пируват декарбоксилируется в присутствии ТРР, при этом происходит перенос гидроксиэтильной группы (остаток пирувата) на тиальное кольцо кофермента. Образуется гидроксиэтилтиаминпирофосфат. (катализируется Е1). 2.Далее он вступает в реакцию с окисленным липоамидом с образованием ацетиллипоамида. Р-я катализируется дигидролипоидтрансферазой. 3. Ацетиллипоамид взаимодействует с коферментом А, образуя ацетил – КоА (тоже дигидролипоилтрансфераза). При этом высвобождается восстановленный липоамид, который необходимо перевести в окисленную форму, для осуществления следующего цикла реакций. Это происходит во время 4 стадии. 4.Восстановленный липоамид окисляется дигидролипоилдегидрогеназой при участии ФАД, который связан с ферментом и является как бы «первичным» акцептором протонов. Образуется ФАДН2, который в свою очередь окисляется с помощью НАД+ и образуется НАДН+Н+ (второй и последний акцептор протонов).
Таким образом, итоговая реакция ОДП будет выглядеть так: Пируват + НАД+ + КоА = Ацетил-КоА + СО2 + НАДН+Н+.
 Регуляция ПДГК: Аллостерическая модификация:
Регуляция ПДГК: Аллостерическая модификация:
Комплекс ингибируют АТР, также как и ацетил-КоА и NADH, являющиеся продуктами реакции (ингибирование конечным продуктом). Т. е. активность комплекса снижается когда увеличиваются соотношения: [ATP]/[ADP], [NADH]/[NAD+], И [acetylCoA]/(CoA]. Увеличение соотношения свидетельствует о достаточном количестве энергии в клетке, о ее положительном энергетическом статусе. Поэтому ПДГК и гликолиз ингибируются при окислении жирных кислот (особенно длинноцепочечных). Напротив, AMP, CoA, и NAD+, аллостерически активируют ПДГК (соотношения уменьшаются). Таким образом, ПДГК выключается, когда есть доступное «топливо» в виде жирных кислот и ацелит-Коа и когда концентрация АТР и NADH в клетке высока и включается когда энергетические запросы возрастают и требуется большое количество ацетил-КоА для цикла Кребса.
Ковалентная модификация. В регуляции активности ПДГК позвоночных алллостерическая модуляция дополняется также и вторым уровнем регуляции - ковалентной модификацией. Ферментный комплекс ингибируется при обратимом фосфорилировании специфического остатка серина в одной из 6-ти субъединиц Е1. Как было замечено ранее, помимо ферментов Е1, Е2 и Е3 в состав ПДГК входят также и регуляторные белки, единственная цель которых - регуляция активности комплекса. Специфическая протеинкиназа фосфорилирует и, таким образом, инактивирует Е1, а специфическая фосфатаза удаляет фофатные группы путем гидролиза, активируя Е1. Киназа аллостерически активируется с помощью АТР: когда уровень АТР высок (что отражает достаток энергии в клетке), ПДГК инактивируется путем фосфорилирования Е1. Когда уровень АТР снижается, снижается и активность киназы и фосфатаза удаляет остаток фосфата от Е1, активируя комплекс. (Киназа и фосфорилаза также регулируются и другими в-вами. См. рисунок).
Пируват как ключевой метаболит: пируват Þв синтез аминокислот, лактата, глюконеогенез, различные виды брожений (молочно-, маслянокислое) у бактерий. Может (при окисл. декарбоксилировании) Þацетил-СоА Þсинтез ЖК или Цикл Кребса.
•ДНК-гликозилазы и ДНК-инсертазы. Эксцизионная репарация.
3:в репарац N-алкилирован пуринов и др модфицирован осн-ий вДНК приним ключев роль специфичN-гликозилазы -ферм расшепл-ие гликозид св м/ду модифирован-ми основ-ми и дезоксирибозой. N-гликозилазы испол-ся и при коррекци наруш-ий, сост-щих в спотан дезаминировании цитозина или аденина и превращ-ии их в соответствен в урацил или гипоксантин. Ферменты оставляют вДНК пробелы на месте модифицирован осн-ий. Потом с пом мех-ма выщепления-ресинтеза восстан-тся исходная последов-сть. Урацил-N-гликозила играет важную роль в антимутагенном действии и при исправлении ошибок, происходящих при репликации когда вставл-ся dUTP вместо dTTP. Данная ошибка почти не распознается pol 1 pol 3,поэт без репарац изменный нуклеотид может включ-ся в новосинтезирован цепь при дальнейшей репликации. DNA insertase - ДНК-инсертаза. Ферм эукариот, способн во вр репарацииДНК непосредствен присоед-ть к дезоксирибозе пуриновое основ-е в соответствии с правилом комплементарности
