
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
 БИЛЕТ 18
БИЛЕТ 18
•Структура и функции нуклеотидов, дезоксинуклеотидов и нуклеозидов. Адениловые
динуклеотиды (НАД, НАДФ, ФАД). Связи, формирующие первичную структуру ДНК и РНК-
5'-фосфатный и 3'- гидроксильный концы полинуклеотидных цепей. Вторичная структура ДНК
и РНК. Денатурация и ренатурация ДНК.
 Основания – пуриновые (аденин (А) гуанин (G). Важные интермедиаты - Гипоксантин (ГКс) и ксантин (Кс)Пиримидиновые цитозин (C), Тимин (Т) урацил (U) [Nu1] Нуклеозиды – основание + рибоза/ или дезоксирибоза. [Nu2] Тимин – только дезоксирибоза! Нуклеотид – нуклеозид с фосф. гр. на ОН рибозы.
Основания – пуриновые (аденин (А) гуанин (G). Важные интермедиаты - Гипоксантин (ГКс) и ксантин (Кс)Пиримидиновые цитозин (C), Тимин (Т) урацил (U) [Nu1] Нуклеозиды – основание + рибоза/ или дезоксирибоза. [Nu2] Тимин – только дезоксирибоза! Нуклеотид – нуклеозид с фосф. гр. на ОН рибозы.
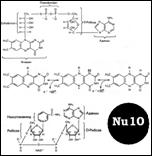
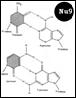
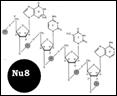
Дезоксинуклеотид – нуклеозид с фосф гр. на ОН дезоксирибозы. Структура ДНК – [Nu8]. Формы спиралей: В-форма. Шаг спирали – 3,4 нм, по 10 п. о. При ↑ Na и К возникает А-форма, с бо’льшим диаметром. Похожа по структуре на двухцепочечную РНК. С-Е формы правоспиральные, только in vitro, Z-форма: левозакрученная, наименее скрученная (12 п. о.) и самая тонкая. Только с 1 желобком. Сущ-ет в повторяющихся последовательностях GC и АС при усл: 1. ↑ Na и К 2. выс содерж отрицат-х супервитков в молекуле ДНК 3. связывание z-специфичных белков 4. метилирование С5 некоторых дезоксицитидинов.
Эта форма рег-ет экспрессию генов. Денатурация при ↑ температуры и ↓ соли. Цепи расходятся. Гиперхромный эффект - ↑ оптич-го поглощения пуринов и пиримидинов. Т плавления – от типа оснований. Зависит от количества Н-Н связей. [Nu9] [Nu10]
•Общая схема источников и путей расходования аминокислот в тканях. Динамическое состояние белков в организме. Основные этапы катаболизма азот-содержащих соединений. Реакции переаминирования. Пиридоксальфосфат. Глутаматдегидрогеназа
 Обновление белков – 1-2% в сутки. 80% высвободившихся АК Þсинтез новых белков. Сут. деградация белков 30-40г (т. е.5-7г азота) Þнужно 30-60 г белка (+!незаменимые АК). !Избыток АК не запасается (подверг. катаболизму). [Азотистый баланс].
Обновление белков – 1-2% в сутки. 80% высвободившихся АК Þсинтез новых белков. Сут. деградация белков 30-40г (т. е.5-7г азота) Þнужно 30-60 г белка (+!незаменимые АК). !Избыток АК не запасается (подверг. катаболизму). [Азотистый баланс].
Метаболизм АК: угл. скелет Þамфибол. интермедиаты; амино. группа Þпереаминирование/ окисл. дезамин. Þ1.5NH3 (рыбы; аммонотелические), 2.5Моч. к-та (птицы; урикотелические), 3.5Мочевина (млеки; уреотелические).
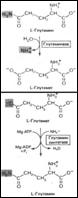 Катаболизм АК: I.Переаминирование. трансаминазы (аминотрансферазы). ВзаимоQ α-АК и α-кетокислоты. В акт. центре фермента – пиридоксальфосфат. 2 вида ферм.: 1.аланин-пируват - (обр-ся аланин) и 2.глутамат-α-кетоглутарат-трансаминазы (обр-ся глутамат). Аланин – субстрат еще и 2ого фермента Þазот всех АК может в кон. итоге переходить в L-глутамат (единст. АК, которая оч. быстро ~>окисл. дезаминирование). Субстратами для трансаминаз не явл-ся: Lys, Tre, Pro и гидрокси-Pro.
Катаболизм АК: I.Переаминирование. трансаминазы (аминотрансферазы). ВзаимоQ α-АК и α-кетокислоты. В акт. центре фермента – пиридоксальфосфат. 2 вида ферм.: 1.аланин-пируват - (обр-ся аланин) и 2.глутамат-α-кетоглутарат-трансаминазы (обр-ся глутамат). Аланин – субстрат еще и 2ого фермента Þазот всех АК может в кон. итоге переходить в L-глутамат (единст. АК, которая оч. быстро ~>окисл. дезаминирование). Субстратами для трансаминаз не явл-ся: Lys, Tre, Pro и гидрокси-Pro.
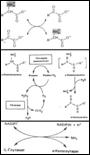
II.Окислительное дезаминирование. L-глутаматдегидрогеназы и оксидаза L- и D-аминокислот (роль???). Ферменты =окисляемые флавопротеины (FMN & FAD окисл-ся непоср. О2 ÞН2О2 Þрасщепление каталазой. Оксидазы АК Þα-иминок-та Þ+Н2О Þсоотв. α-кетокислота (потеря α-NH2-группы в виде иона аммония).
III.L-глутаматдегидрогеназа. Ингибиторы: ATP, GTP, NADH; стимуляторы: ADP. Гормон. регуляция. Косубстрат: NAD или NADP +р-ция обратима Þused и для синтеза, и для биосинтеза АК.
IV.Образование аммиака. В тканях +много бактериями в киш-ке из белка и мочевины Þv. porta Þhepar (!NH3 ~>в кровоток при циррозе Þколлатерали с v. cava inf. Þотравление ЦНС).
Отдельный трёп: 5NH3 в поч. канальцах (поддерж. КОС) из амидного-азота глутамина (а не из мочевины; не α-аминогруппа used!; глутманаза).
V.Транспорт аммиака. Глутаматдегидрогеназа, обр-ие глутамина: глутаминсинтетаза (мх. фермент, тратим АТР). Обр. процесс – за счет глутаминазы. Аналогичный механизм – L-аспарагиназа.
Обмен аминокислот – гл. роль – у мышц (тут освобожд-ся 50% аминок-т, половина образующегося в мышцах азота – Ala и Gln) и печени (осн. поглощ. Ala) и кишечника (осн. поглощ. Gln). Почки – гл. источник серина, поглощ. Gln, Pro & Gly), Мозг – поглощ. Val (в мозге проще окисл-ся АК с разветвл. бок. цепью).
 !Аланин – глюкогенная АК (Þглюконеогенез в печени).
!Аланин – глюкогенная АК (Þглюконеогенез в печени).
БИОСИНТЕЗ МОЧЕВИНЫ
За сутки выделяем 16,5 г азота (85% ~>мочевина Þэкскреция).
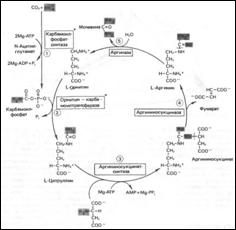
Цикл мочевины: 1.Синтез карбамоилфосфата (карбамоилфосфатсинтаза, в мх., тратим 2 АТР, нужен Mg2+ и дикарб. к-та); 2.Синтез цитруллина (L-орнитин-карбамоилтрансфераза, мх. печени, высокоспециф.); 3.Синтез аргининосукцината (аргининосукцинат-синтаза, тратим АТР); 4.Расщепление аргининосукцината на Arg и фумарат (аргининосукциназа; фумарат далее Þ*оксалоацетат, или аспартат при переаминировании); 5.Расщепление Arg на орнитин и мочевину (аргиназа).
НАРУШЕНИЯ ЦИКЛА МОЧЕВИНЫ
•Гипераммониемия I типа (недостаток карбамоилфосфатсинтазы; наслед.); •Гипераммониемия II типа (недостаток орнитин-карбамоилтрансферазы, сцепл. с Х-хромосомой, ↑[Gln] в крови Ü↑синтеза глутамина); •Цитруллинемия (много цитрул. в моче, плазме, ликворе; дефект аргининосукцинат-синтазы); •Аргининосукцинатная ацидурия (наслед. рецесс. тип; ↑арг.-сукцината в крови, Qроста волос; недостаток аргининосукциназы).
•Генетический код.
Больш часть генетич инф-ции, содержащ в ДНК, код-ет последоват-ть ам. к-т. Процесс экспрессии генетич инф-ции включ транскрипцию«текста»,записан на«языке нуклеин к-ты»,в текст, записан на«языке белков».Таково происхож-е термина трансляц, испол-мого для обознач процесса биосинт белков. Правила, кот следует трансляция, назыв генетич кодом. В биосинт участв20 ам. к-т, наз-ых протеиногенными,«язык» нуклеиновых к-т должен содерж по крайней мере 20 слов(кодонов)Но в ам. к-тном«алфавите»им-ся только 4«буквы» (A, G, С и U или Т*]),так что для получ-я20 различ слов каждое должно сост-ть по крайней мере из 3букв. Кодоны действит-но включают 3азотистых основ-я(триплет нуклеотидов).Изобр-ный в виде круга. Схема чит-ся от центра наружу, так что, напр, триплет CAT кодирует ам. к-ту His. ДНК-кодоны идентичны таковым в мРНК, за исключ того, что в мРНКвместо урацила (U),хар-ого для ДНК, стоит тимин(Т).Примера прочтения кода на сх 2 показаны коротк участ нормал-го и мутантного гена β-глобина вместе с соответств-ми последоват-ми мРНК и ам. к-т. Показаны относ-но часто встреч-ся точковые мутации, в резул кот остаток глутаминовой к-ты в полож 6 β-цепи заменен на валин. Такой мутантный Hb в дезоксиформе склонен к агрегации, что вызыв деформац ЭЦ и уменьшает эффек-ть транспорта кислорода(серповидноклеточная анемия).В триплетном генетич коде для 20 ам. к-т потенциально существует43=64 кодона. Т.обр, бол-во ам. к-т записыв-ся нескол кодонами, т.е. генетич код явл вырожденным. Так же им 3триплета, кот обознач конец транскрипции(стоп-кодоны).Еще 1спец кодон, стартовый(инициирующий)кодон, маркирует начало трансляц.
