
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Новая эра в исследовании остеопороза: гомогенный анализ резорбции костей in vitro.
В норме перестройку здоровых костей осуществляют два типа клеток: остеобласты, которые синтезируют новую костную ткань, и остеокласты, которые рассасывают старую. Вновь обнажаемая поверхность рассасывающейся кости притягивает остеобласты и инициирует синтез новой костной ткани. Среди ряда заболеваний костей остеопороз имеет наибольшее значение и наиболее широко изучается. Остеопороз – метаболическое заболевание костей, характеризуется истощением костной ткани и потерей массы костей. Признаки этого заболевания, прогрессирующие с его течением, – хрупкость костей, увеличение риска переломов. По мере старения населения разработка новых препаратов для лечения остеопороза стала важнейшей задачей фармацевтической промышленности. Планируется, что продажи лекарств от остеопороза на мировом рынке в 2006 г. достигнут 11,5 миллиардов долларов.
Фармацевтические разработки в области остеопороза далеко продвинулись с развитием культур остеобластов, остеокластов и их предшественников in vitro. Можно заставить дифференцироваться in vitro мезенхимные стволовые клетки в остеобласты, а предшественники остеокластов – в функциональные остеокласты. Тем не менее, анализ костной резорбции in vitro – узкое место клеточных методов разработки препаратов. Современные функциональные анализы остеокластов in vitro включают в себя цитохимическое окрашивание красителями типа TRAP (тартрат-устойчивая кислотная фосфатаза) на активность кислотной фосфатазы, иммуноферментный анализ (ИФА) для определения специфических продуктов деградации матрикса костей (например, C и N-концевые пептиды коллагена) анализ изображений следов, которые оставляют остеокласты при разрушении естественной или синтетической кости. Эти анализы требуют трудоёмкой пробоподготовки и обработки образцов, они либо дороги, либо необъективны.
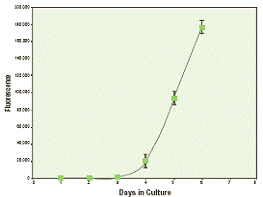
Мы с радостью представляем новый гомогенный анализ, который существенно облегчит измерение резорбции костей in vitro и повысит производительность анализа. Основой для разработки набора OsteoLyse (на человеческом коллагене) послужил тот факт, что костная масса примерно на 30% состоит из коллагена типа 1, который разрушают остеокласты в процессе резорбции. В этом методе предшественники остеокластов высевают на 96-луночный планшет для коллагена OsteoLyse, и они дифференцируются в присутствии ростовых факторов M-CSF и растворимого RANK-лиганда. Остеокласты начинают созревать примерно через 4 дня после посева и в это время приобретают способность разрушать меченный лантанидами человеческий коллаген типа 1 на поверхности планшета OsteoLyse (рисунок 1).
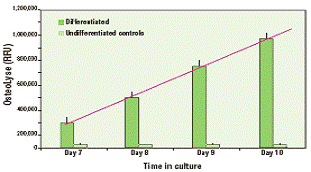
Выход фрагментов разрушения коллагена определяют, измеряя флуоресценцию в 10 мкл пробе супернатанта культуры клеток; пробу добавляют к 200 мкл выделяющего флуорофор реагента и анализируют с помощью флуориметра, измеряющего флуоресценцию с временным разрешением. Небольшой объём пробы, необходимый для анализа, позволяет многократно отбирать пробы культуры и ставить последовательные эксперименты во времени. Культуры предшественников остеокластов в присутствии M-CSF, но без RANK-лиганда, служат в качестве недифференцированного контроля (рисунок 2). Отношение образец-контроль (дифференцированные клетки/недифференцированные клетки) повышалось при замене культуральной среды; в течение 24 часов после замены среды это отношение могло быть >10 и могло достигать 35 при отборе супернатантов через 3 дня после замены среды. Методы могут различаться по влиянию тестируемого образца на дифференциацию предшественников и функцию зрелых остеокластов. Последняя требует присутствия образца в течение 5 дней, необходимых для дифференциации клеток; образец удаляется при замене среды. При поиске ингибиторов функции зрелых остеокластов образец в первый раз добавляют при замене среды на 5-7 день.
Рисунок 1. Для начала дифференцировки первичным человеческим предшественникам остеокластов требуется 4 дня культивации. Первичные человеческие предшественники остеокластов высевали на планшеты OsteoLyse по 10.000 клеток в лунку. Каждые 24 часа отбирали и анализировали пробы культурального супернатанта (10 мкл).
|
|
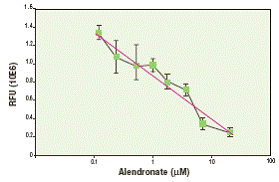
Как показано на рисунке 3, предшественники остеокластов дифференцировались либо в присутствии 1 нМ кальцитонина, либо в его отсутствие. Когда кальцитонин добавляли в культуру при замене среды, он ингибировал резорбцию костного матрикса только в тех культурах, в которые его не добавляли в день 0. Эти данные демонстрируют угнетение рецептора кальцитонина в остеокластах, подвергшихся обработке кальцитонином. В другом эксперименте с помощью набора OsteoLyse изучали влияние алендроната фосфоната на резорбцию костей in vitro. Значение IC50 (примерно 2 мкМ) рассчитывали на основе данных, показанных на рисунке 4.
Рисунок 2. Флуоресцентный выход коллагеновых пептидов при дифференцировке первичных человеческих остеокластов повышается со временем культивации.
|
 дни в культуре
дни в культуре
Рисунок 3. Ингибирование резорбции костного матрикса в анализе OsteoLyse. Первичные человеческие предшественники остеокластов высевали на планшет OsteoLyse и культивировали в среде для дифференциации, не содержащей кальцитонин, с добавлением кальцитонина только на 5 сутки или с добавлением кальцитонина между 0 и 5 сутками.
|
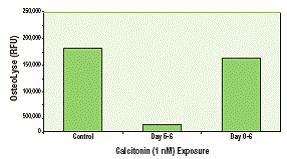
Рисунок 4. Анализ ингибирования функции остеокластов, опосредованного алендронатом. Первичные человеческие предшественники остеокластов высевали на планшет OsteoLyse с различными концентрациями алендроната или без него. Через 7 дней заменяли среду на свежую и ещё через 3 дня отбирали культуральные супернатанты.
|
Новый набор OsteoLyse для анализа коллагена требует минимума ручных операций с культивируемыми клетками или реагентами/образцами и даёт исследователям новый ценный метод поиска препаратов. Этот новый метод намного проще при том, что он позволяет с высокой чувствительностью и достоверностью исследовать функцию остеокластов in vitro.
Клетки на заказ: неограниченные возможности.
Мы предлагаем большое разнообразие типов клеток и сред для них, но Вам может понадобиться продукт, которого нет в нашем каталоге. С 1997 года у Cambrex действует Отдел клеток на заказ, который предлагает услуги по выделению на заказ типов клеток, не относящихся к стандартной продукции. За 20 лет работы в области культур клеток сотрудники наших лабораторий достигли больших успехов и приложат все усилия, чтобы обеспечить продукт на заказ под Ваши специфические потребности.
Отдел клеток на заказ Cambrex будет работать с Вами на индивидуальной основе, чтобы разработать методики, гарантирующие успех Вашего проекта. Благодаря обширным источникам человеческих и животных тканей, костного мозга и периферической крови, стимулированной гранулоцито-стимулирующим фактором (G-CSF), мы можем предложить Вам большую часть необходимых типов клеток. Мы обеспечиваем заказанное количество клеток, среды и услуги по упаковке. Предлагаются также услуги по контролю качества различными методами, чтобы Вы могли удостовериться в высоком качестве и жизнеспособности клеток.
Отдел клеток на заказ Cambrex предлагает полный набор человеческих клеток, включая астроциты, клетки эндотелия микрокапилляров мозга и сердечные фибробласты. Далее, мы можем Вам предложить животные клетки на заказ, включая клетки почек африканских зелёных обезьян, клетки гладкой мускулатуры радужной оболочки глаза кошек и кардиомиоциты крыс. С полным списком клеток и дополнительных услуг Вы можете ознакомиться на нашем сайте: www. /content/Bioscience/humananimalcells. asp.
Гелевый краситель для нуклеиновых кислот GelStar.
Часто задаваемые вопросы.
1) Каковы преимущества красителя GelStar перед этидий бромидом? Краситель GelStar как минимум в 5-10 раз более чувствителен к окрашиванию ДНК и РНК в геле, чем этидий бромид, и позволяет определять всего 20 пг двухцепочечной ДНК или 3 нг РНК. Гели, окрашенные GelStar, имеют чёткие полосы с очень низкой фоновой флуоресценцией.
2) Каковы преимущества красителя GelStar перед SYBR зелёным I? Краситель GelStar разработан специально для агарозных гелей и может использоваться для них непосредственно перед снятием результатов или использоваться для гелей после окрашивания. В отличие от красителя SYBR зелёного I, гели, предварительно окрашенные красителем GelStar, сохраняют высокую чувствительность и разрешение, чёткое проявление полос. Более того, краситель GelStar позволяет определять как ДНК, так и РНК, в то время как краситель SYBR зелёный I подходит в основном для ДНК.
3) Как обнаружить ДНК, окрашенную красителем GelStar? Краситель GelStar можно обнаружить с помощью УФ просвечивания, лазерных сканеров или просвечивания с помощью Dark Reader.
4) Совместим ли краситель GelStar с моей системой проявки изображения? Проявлять гели можно с помощью Поляроида, CCD или системы лазерного сканирования. Для максимально эффективной проявки Cambrex предлагает фотофильтры (50536) для гелевого красителя GelStar, используемые с фотоаппаратами Polaroid с желатиновыми фильтрами. Для аппаратов со стеклянными фильтрами подходят ярко-жёлтые фильтры, например, Tiffen #15. В камеры CCD обычно вставлены зелёные фильтры SYBR, оптимальные для красителя GelStar. Длины волн испускания и возбуждения для красителя GelStar – 527 и 493 нм соответственно. Для выбора фильтра, который лучше использовать с красителем GelStar, обратитесь к производителю Вашего оборудования.
5) Как утилизировать краситель GelStar? GelStar связывается с нуклеиновыми кислотами, и поэтому к нему следует относиться как к потенциальному мутагену. Следуйте Вашим инструкциям по утилизации опасного материала. Раствор красителя обычно выливают в активированный уголь, который удерживает краситель. Буферный раствор остаётся безопасным, а угольный фильтр следует сжигать.
Научное применение мезенхимных стволовых клеток.
Стволовые клетки имеют потенциал для самовосстановления, обладают недифференцированным фенотипом и способны дифференцироваться по разным направлениям (Marshak et al., 2000). Стволовые клетки имеются как в развивающемся, так и во взрослом организме и имеют значение как для формирования органов во время развития, так и для регенерации тканей при повреждении. Стволовые клетки всё больше используют для анализа на токсичность in vitro и функционального тестирования при поиске препаратов. Значение стволовых клеток в современной биотехнологии возрастает по мере того, как всё большее число типов клеток становятся доступными для изучения.
Потенциальные источники стволовых клеток – эмбрионы, новорожденные и взрослые организмы. Клетки эмбриона перед имплантацией – источник клеток, которые культивировали как плюрипотентные линии эмбриональных стволовых клеток. Валики половых желёз плода на ранних стадиях – источник очевидно плюрипотентных зародышевых клеток, а другие ткани эмбриона на более поздних стадиях – источники мультипотентных клеточных линий нервной ткани или печени. Ткани новорожденных, например, пуповины, – источники мультипотентных клеток. Взрослые соматические ткани – также источник стволовых клеток, например, костный мозг, периферическая кровь, кожа, жир, мускулатура, мозг и печень. Доступные и восстанавливающиеся взрослые ткани, такие как костный мозг, периферическая кровь, жир и кожа, являются наиболее ценным источником стволовых клеток.
Взрослый костный мозг – наиболее часто используемый источник мультипотентных стволовых клеток, которые дают начало различным тканям мезодермального происхождения. Гематопоэтические стволовые клетки дают начало линиям на основе крови, включая миелоидные, лимфоидные и эритроидные клетки. Гематопоэтические стволовые клетки, полученные из костного мозга напрямую или из мобилизованной периферической крови, способны восстанавливать клеточный пул крови после разрушающей мышечную ткань химиотерапии или действия радиации. Кроме того, внутренняя губчатая часть костного мозга – строма – содержит различные прикрепляющиеся клетки, включая недифференцированные – мезенхимные стволовые клетки. Мезенхимные стволовые клетки дают начало скелетной мускулатуре и соединительной ткани, и обширные исследования подтвердили, что эти клетки являются предшественниками костей, хрящей, мускулатуры, сухожилий и жировой ткани.
Рисунок 1. Мезенхимные стволовые клетки дифференцируются в жир, хрящи и кости in vitro, как описали Pittenger, M. F. et al. (1999).
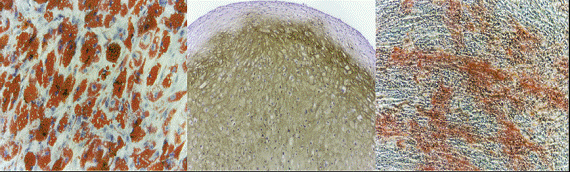
жир хрящ кость
В качестве компонента стромы костного мозга мезенхимные стволовые клетки играют важную роль в поддержке и регуляции продукции кровяных клеток. Они синтезируют белки внеклеточного матрикса и многочисленные цитокины и ростовые факторы. Регуляция мезенхимных стволовых клеток активно изучается в связи с трансплантацией гемопоэтических стволовых клеток и может существенно помочь при приживлении трансплантанта и восстановлении продукции клеток крови. Также было показано, что мезенхимные стволовые клетки почти не вызывают иммунного ответа при испытаниях на иммунологически чужеродных животных моделях.
Мезенхимные стволовые клетки – прекрасная модель для изучения клеточной дифференциации in vitro. Хорошо изучены три пути, по которым может идти дифференциация этих клеток in vitro, образование минерализованных костей, хрящей и жировой ткани (адипоцитов). Регуляция этих путей представляет большой интерес для изучения артрита, остеопороза и ожирения, а также многих других заболеваний. Использование мезенхимных стволовых клеток для восстановления и/или регенерации повреждённых связок и костей – ещё одна область активных фундаментальных и клинических исследований. Мезенхимные стволовые клетки также имеют потенциал для дифференцировки в мускулатуру, хотя этого труднее добиться in vitro. Тем не менее, исследования на животных показали способность этих клеток восстанавливать повреждённую сердечную мышцу и улучшать её работу.
Факторы, влияющие на дифференцировку стволовых клеток, изучались in vitro и in vivo. При определённых условиях культивации полипептидные ростовые факторы и цитокины влияют на выбор пути развития мезенхимных стволовых клеток. Также на него может влиять пространственная организация клеток, в том числе клеточная плотность. Отдельные виды клеток, такие как адипоциты и хондроциты, видимо, требуют специальной двух - или трёхмерной конфигурации. Механические силы могут повлиять на тип, сроки и объёмы дифференциации скелетной мускулатуры. Наконец, имеет значение для дифференциации состояние пролиферации клеток. Недавно Jaiswal et al. (2000) показали, что мезенхимным стволовым клеткам для запуска дифференциации в костную ткань требуется активная MAP-киназа. Если этот сигнальный путь заблокирован, дифференциация клеток идёт по пути образования жировой ткани. Из этого наблюдения следует, что дифференциация стволовых клеток регулируется сигнальными белками, включая протеинкиназы. Таким образом, возможности использования стволовых клеток для изучения механизмов клеточной дифференциации огромны. Более того, стволовые клетки могут служить мишенями для разрабатываемых фармпрепаратов, особенно при возрастных заболеваниях.
Также имеются огромные возможности использования мезенхимных стволовых клеток в генной терапии. Мезенхимные стволовые клетки здоровых доноров могут быть эффективны при замещении недостающего белка у пациентов с рядом заболеваний, особенно при отсутствии сильного иммунного ответа. Мезенхимные стволовые клетки также легко трансдуцировать, если требуется вставка или усиление экспрессии конкретного гена.
Рисунок 2. Влияние на дифференциацию мезенхимных стволовых клеток.
· Ростовые факторы и цитокины } например, TGF-b; IL-1; BMP |
· Основные питательные компоненты } например, кислород; глюкоза; сульфат; пролин |
· Плотность клеток/пространственная организация } например, однородность монослоя; кластеры |
· Механические силы } например, растяжение; сжатие; повторяемость |
· Состояние пролиферации } например, состояние покоя; циклы; старение |
Подводя итог, можно сказать, что мезенхимные стволовые клетки – это доступные стволовые клетки взрослых, обещающие как обширное научное применение, так и клиническое. Эти клетки могут пролиферировать как стволовые клетки в культуре, и также их можно заставить дифференцироваться по нескольким путям в зрелые функциональные клетки. Эта система обладает огромным потенциалом во многих областях клеточной биологии.
Набор для протеинкиназного анализа высокопроизводительного скрининга (HTS) PKLight: быстрый массовый протеинкиназный анализ HTS.
Введение.
Благодаря своей способности фосфорилировать белки протеинкиназы играют ключевую роль в механизмах передачи сигналов при многих заболеваниях, таких, как рак, артрит и диабет. Эта роль протеинкиназ делает из мишенями для многих групп HTS, производимых фармацевтической промышленностью. Современные технологии скрининга, такие как гомогенная флуоресценция с временным разрешением (HTRF), флуоресцентная поляризация (IMAP) и анализ совпадения сцинтилляции (SPA) включают использование специфичных к определённым состояниям фосфора антител или радиоактивных зёрен. Для нашей технологии не требуются антитела, зёрна, АТФ с радиоактивной меткой или специальные последовательности субстрата, и она подходит для широкого круга серин-/треонин - и тирозинкиназ.
Принципы.
Анализ PKLight – простая система детекции в одной точке, измерения потребления АТФ киназой при фосфорилировании субстрата (рисунок 1). В анализе используется фермент люцифераза, который светится при потреблении АТФ. В анализе PKLight поглощение АТФ определяют путём точного измерения АТФ, оставшейся в лунках после действия киназы. Интенсивность излучаемого света линейно зависит от концентрации АТФ и измеряется с помощью люминометра.
Установка.
Анализ PKLight – массовый киназный анализ in vitro, где единственным требованием является чистота киназы, субстрата и АТФ. Нет никаких специфических требований, например, антител или модифицированных субстратов, что делает анализ очень быстрым, простым и недорогим.
Рисунок 1. γ-фосфатный остаток АТФ вырезается киназой и вслед за этим вставляется в субстрат, превращая АТФ в АДФ.
|
|
|
|
|
|
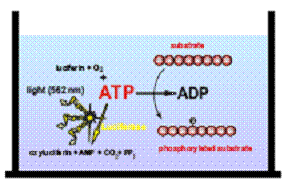
оксилюциферин + АМФ + CO2 + PP1
Рисунок 2. Калибровочная кривая протеинкиназного (PKA) анализа для 0.005-5.00 единиц PKA для фосфорилирования 5 мкМ субстрата кемптида. Вставка: ингибирование PKA H-9 дигидрохлоридом, (0.01 мкМ – 100 мкМ). 0.03 единицы PKA, 5.0 мкМ субстрата и 1 мкМ АТФ. Значение IC50 определено для 2.0 мкМ H-9 гидрохлорида.
|
|
|
|
|
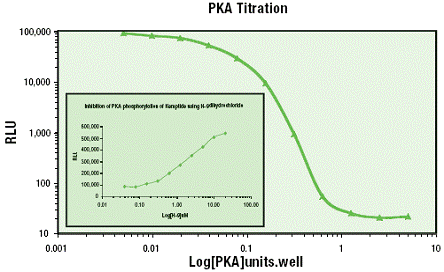
К примеру, был поставлен эксперимент для cАМФ-зависимой протеинкиназы (PKA) с её субстратом, кемптидом (рисунок 2). EC50 (эффективная концентрация PKA, дающая 50% максимального ответа) была определена как ~0.03 единицы. Эта величина определяет пределы достоверности анализа и, следовательно, максимальную разницу значения сигнала для минимальной разницы концентрации АТФ. Эту концентрацию киназы затем использовали в дальнейших экспериментах по ингибированию с известным ингибитором PKA, H-9 дигидрохлоридом.
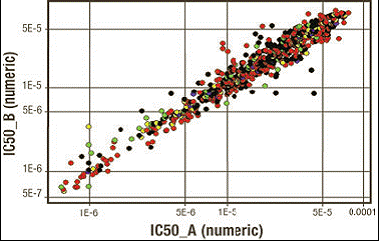
Рисунок 3. Корреляция значений IC50 при постановке экспериментов в двух повторах методом PKLight.
|
|
Таким образом, становится возможным точное измерение IC50 (концентрации ингибитора при 50% максимального ответа) интересующего нас вещества. Возрастающие концентрации H-9 дигидрохлорида дают стандартную кривую ингибирования со значением IC50 ~2.0 мкМ. И EC50, и IC50 просто измерить, чтобы провести сравнение препаратов, ферментов или разных методик.
Параметр | Значение |
Формат планшета | 384 |
[Вещество], мкМ | 10.0 |
Число компонентов | 444.321 |
Средний % ингибирования | 0.7 |
Предел положительных значений | 18% |
Число положительных значений | 3%) |
Средний Z-фактор | 0.69 (±0.09) |
Средний Z'-фактор | 0.74 (±0.09) |
Рисунок 4. Скрининг на ингибирование более чем 400.000 веществ с помощью анализа PKLight. Пороговое значение определено как >18% ингибирование.
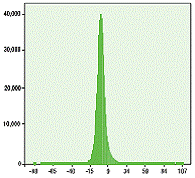
Накопительная диаграмма ингибирования в процентах (шаг 1%)
Скрининг на ингибирование.
Было показано, что анализ PKLight хорошо коррелирует с другими методиками, например, SPA и IMAP, давая возможность просто, с высокой точностью и воспроизводимостью определять IC50. На рисунке 3 получены значения IC50, полученные в экспериментах по два повтора методом PKLight, которые демонстрируют высокий уровень корреляции и воспроизводимости.
Анализ PKLight позволяет уменьшать объём пробы (96-, 384-, 1536-луночные планшеты и т. д.) и был разработан для применения в HTS. Этим методом в формате 384-луночного планшета было проанализировано на ингибирующий эффект по отношению к киназе более 400.000 веществ. Параметр скрининга был установлен на уровне 18% ингибирования (любое вещество, вызывающее ингибирование выше этого порога, учитывалось как положительный результат. Пользуясь этим пределом, 3.998 веществ (0.9%) определили как положительные, со средним значением Z' 0.74 (рисунок 4). Значение Z' – уровень качества анализа, относящийся к стандартному отклонению повторных измерений (для анализа HTS допустимым является значение более 0.5).
Заключение.
Анализ PKLight – гомогенная система детекции в одной точке для любой тирозин - или серин-/треонин - киназы. Анализ идеально подходит для HTS, где требуется определение АТФ in vitro. Методика легка для отработки и постановки, требует минимальной оптимизации и хорошо коррелирует с известными методами, даёт чёткие результаты с высоким уровнем достоверности.
Важные лабораторные методики для нормальной человеческой культуры клеток.
Нормальные человеческие клетки Clonetics (NHC) – прекрасная альтернатива традиционным клеточным линиям. Эти выделенные первичные клетки не иммортализованы, не трансформированы и являются идеальной заменой при невозможности работать in vivo. Тем не менее, необходимо помнить, что нормальные человеческие клетки чувствительны к колебаниям окружающей среды и условиям культивирования. Ниже приведены некоторые важные практические замечания и моменты, которые необходимо учитывать при работе с NHC.
Хранение и разморозка нормальных человеческих клеток Clonetics.
· Согласно инструкции, замороженные клетки необходимо немедленно поместить в жидкий азот и хранить там до начала разморозки и переноса в планшет. Хранение клеток при более высокой температуре, даже краткосрочное, может вызвать существенные повреждения и гибель клеток.
· Подготовьте культуральную посуду с заранее прогретой средой до разморозки NHC. Излишнее нахождение размороженных клеток вне культуры может привести к их повреждению.
· Погрузите размороженные ампулы в водяную баню при 37°C и держите, пока не останутся только маленькие кристаллики льда (обычно 1-2 минуты). Излишнее нагревание может вызвать необратимое повреждение клеток. Клетки не следует размораживать на рабочей поверхности стола или вращая ампулу в руках.
· Никогда не центрифугируйте клетки Clonetics при разморозке. Удаляйте остаточный DMSO, заменяя среду после первых 24 часов культивации клеток.
Культивирование нормальных человеческих клеток Clonetics.
· Рассчитайте количество культуральной посуды, пользуясь данными ниже уравнениями и информацией из Сертификата анализа, приложенного к каждой партии клеток Clonetics:
Число клеток в ампуле ´ процент жизнеспособности = максимум см2 для культуры
Рекомендуемая плотность рассева
Максимальное число занятых см2 = максимальное число культуральных флаконов
Эффективная ростовая поверхность флакона
Примечание: Клетки не следует помещать в один флакон T-25. Для каждого типа клеток рекомендуется своя плотность рассева, оптимальная для прикрепления и пролиферации клеток. Для роста в 96-луночном планшете клетки следует засевать по 10.000 клеток/см2.
· Заменяйте ростовую среду после первых 48 часов культивирования и затем каждые 48 часов, пока клетки не будут готовы к пересеву. Никогда не замораживайте и не нагревайте повторно дополнительную бутыль среды. Отбирайте и нагревайте только необходимый для добавления клеткам объём, предотвращая таким способом сокращение срока хранения ростовой среды. Когда прикрепление клеток достигнет 50%, увеличьте вдвое объём ростовой среды, чтобы избежать нехватки питательных веществ.
· Клетки следует пересевать при 70-90% прикреплении, чтобы избежать контактного ингибирования клеточной популяции и возможных трудностей при обработке трипсином.
· Для большего успеха во время пересева следует проводить подсчёт количества клеток и жизнеспособности, чтобы убедиться, что клетки пересеваются в рекомендуемой плотности.
Практические аспекты нормальных человеческих клеток Clonetics.
· Нормальные человеческие клетки не образуют сплошного слоя за один день. Через 24 часа в поле микроскопа будет немного прикрепившихся клеток. Отберите всю среду со взвешенными в ней клетками и замените на свежую среду. После этого прикрепившиеся к поверхности клетки начнут пролиферировать. Сплошной монослой формируется за 5-9 дней.
· Каждая система сред Clonetics оптимизирована для ведения и пролиферации типа клеток Clonetics, для которого она рекомендована. Эти уникальные системы нельзя заменить на RPMI или DMEM с сывороткой. Не следует использовать основную среду или добавки с истекшим сроком хранения. Готовую ростовую среду с добавками можно хранить 30 суток при 4°C.
· Если в систему сред входит эмбриональная телячья сыворотка, прогревать её не надо. Прогревание сыворотки может оказать неблагоприятное воздействие на состояние клеток и привести к денатурации необходимых компонентов.
· Важным моментом является разница между удвоением популяции и пассажем. Удвоение популяции – время, необходимое популяции клеток для количественного удвоения. Пассаж – отбор и перенос клеточной популяции из флакона в несколько флаконов или один более крупный. Для клеток Clonetics эту процедуру лучше всего выполнять с помощью культуральных реагентов ReagentPack (CC-5034). ReagentPack состоит из раствора HEPES в фосфатно-солевом буфере, трипсина/ЭДТА и трипсин-нейтрализующего раствора (TNS).
· Для клеток Clonetics важно использовать именно ReagentPack. Первичные человеческие клетки особо чувствительны к концентрации фермента. Трипсин в обычных концентрациях или при температуре выше комнатной может вызвать необратимое повреждение клеточных мембран. Для открепления клеток от поверхности флакона может потребоваться энергичное постукивание по нему. Как только клетки открепятся от поверхности, трипсин следует нейтрализовать с помощью TNS. Различные системы сред Clonetics оптимизированы для пролиферации клеток в отсутствие сыворотки, и для нейтрализации трипсина их недостаточно. Более того, настоятельно рекомендуется использовать TNS и трипсин в отношении 2:1. Нарушение этого соотношения может привести к значительному повреждению клеток.
С рекомендациями и инструкциями по обращению с нормальными человеческими клетками Clonetics можно ознакомиться по адресу www. /Content/bioscience/CatNav. oid.411 или обратившись к специалистам нашей технической службы по электронной почте *****@***com.
Правильный выбор агарозы для разделения малых фрагментов.
При большом выборе агарозы, которую мы предлагаем, нам часто задают вопрос: «Какая агароза подходит для моих задач или размеров фрагментов?» Мы предлагаем много возможностей для ответа на этот вопрос: Руководство по выбору агарозы в нашем каталоге, Брошюра по электрофорезу в агарозном геле, Настольная книга по гель-электрофорезу, веб-сайт Cambrex, отдел продаж и специалисты технической службы. Мы часто получаем запросы по выбору агарозы для разделения малых фрагментов. В этой статье разбираются три продукта, рекомендованных для разделения ДНК длиной менее 1 кБ: NuSieve 3:1, MetaPhor и агароза NuSieve GTG.
Агароза SeaKem LE, рекомендуемая для рутинного анализа нуклеиновых кислот, также подходит для разделения малых фрагментов. Можно ожидать удовлетворительного разделения малых фрагментов, но агароза SeaKem LE обладает широким спектром разделения (от 500 н. п. до 23 кБ), и может не позволять различать полосы ДНК малых размеров.
Агароза NuSieve 3:1 для ПЦР-анализа.
Уже в 1990 г., когда мы разработали в своих лабораториях агарозу NuSieve 3:1, её рассматривали как специальную агарозу для ПЦР-анализа. При стандартной температуре плавления агарозы (£90°C) агароза NuSieve 3:1 формирует простой в обращении гель, что делает её идеальной как для блота, так и для проверки выхода и/или чистоты ПЦР-продуктов. Она обеспечивает прекрасное разрешение для фрагментов от 50 н. п. до 1 кБ. Как и остальные продукты, упоминаемые в этой статье, агароза NuSieve 3:1 не обладает заметной РНКазной или ДНКазной активностью. К тому же в многочисленных публикациях агароза NuSieve 3:1 упоминается как агароза для ПЦР-анализа.
NuSieve 3:1 | Конечная концентрация агарозы, % (вес/объём) | |
Спектр размеров (н. п.) | 1´ TAE буфер | 1´ TBE буфер |
500-1.000 | 3.0 | 2.0 |
100-500 | 4.0 | 3.0 |
10-100 | 6.0 | 5.0 |
Агарозный гель для ПЦР-продуктов NuSieve 3:1.
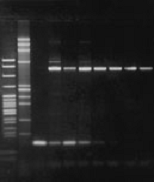 Последовательность l-ДНК длиной 550 н. п. амплифицировали 25 циклов в различных условиях. ПЦР-продукты подвергали электрофорезу на 4% агарозном геле NuSieve 3:1 в 1´ TAE буфере при 5 В/см в течение трёх часов. Дорожка 1: Msp I – сборная полоса ДНК pBR322; дорожка 2: Hae III – сборная полоса ДНК fX174; дорожка 3: праймер-димерный контроль; дорожки 4-9: ПЦР-продукты.
Последовательность l-ДНК длиной 550 н. п. амплифицировали 25 циклов в различных условиях. ПЦР-продукты подвергали электрофорезу на 4% агарозном геле NuSieve 3:1 в 1´ TAE буфере при 5 В/см в течение трёх часов. Дорожка 1: Msp I – сборная полоса ДНК pBR322; дорожка 2: Hae III – сборная полоса ДНК fX174; дорожка 3: праймер-димерный контроль; дорожки 4-9: ПЦР-продукты.
Агароза для совершенного разделения MetaPhor.
Агароза MetaPhor обладает наиболее высоким разрешением из всех доступных, её разрешение вдвое больше, чем у стандартной агарозы, и приближается к полиакриламиду. Агароза MetaPhor позволяет разделять фрагменты, отличающиеся по размеру только на 2% (к примеру, 200 н. п. и 204 н. п.). Существуют альтернативные методики, способные сократить эту разницу до 1%. Агароза MetaPhor рекомендуется для разделения очень маленьких фрагментов, от 20 до 800 н. п. Она оптимальна для множественного анализа ПЦР, STR или AMPFLP и позволяет выполнять эти анализы, не создавая опасностей, характерных для полиакриламида.
MetaPhor | Конечная концентрация агарозы, % (вес/объём) | |
Спектр размеров (н. п.) | 1´ TAE буфер | 1´ TBE буфер |
150-800 | 2.0 | 1.8 |
100-600 | 3.0 | 2.0 |
50-250 | 4.0 | 3.0 |
20-130 | 5.0 | 4.0 |
<80 | - | 5.0 |
Сравнение разрешения для коротких цепей ДНК.


3,5% агароза MetaPhor Горизонтальный формат | 8% полиакриламид Вертикальный формат |
Цепи ДНК с шагом 4 н. п. (дорожка 3) или 8 н. п. (дорожка 1), приготовленные с помощью сшивания линкеров BgI II. Дорожка 2: Msp I – сборная полоса ДНК pBR322. Анализ в агарозе MetaPhor проводили при 6.7 В/см в течение 4 часов при 15°C. Анализ в полиакриламидном геле проводили при 8 В/см в течение 2 часов.
Агароза для восстановления NuSieve GTG.
Агароза NuSieve GTG – агароза класса для генных технологий (GTG), качество которой проверяли относительно устойчивых стандартов, чтобы удостовериться в её оптимальных свойствах. Низкая температура плавления агарозы (£65°C) обеспечивает отличное разрешение для малых фрагментов, её свойства проверены в наших лабораториях на надёжность подготовки и сшивания фрагментов нуклеиновых кислот. Агароза NuSieve GTG идеальна для восстановления малых фрагментов. Помимо стандартных методик восстановления, эта агароза подходит также для реакций внутри геля, что экономит время и позволяет избежать возможных потерь при восстановлении.
NuSieve GTG | Конечная концентрация агарозы, % (вес/объём) | |
Спектр размеров (н. п.) | 1´ TAE буфер | 1´ TBE буфер |
500-1.000 | 2.5 | 2.0 |
150-700 | 3.0 | 2.5 |
100-450 | 3.5 | 3.0 |
70-300 | 4.0 | 3.5 |
10-100 | 4.5 | 4.0 |
8-50 | 5.0 | 4.5 |
Совершенное разделение малых фрагментов ДНК в агарозных гелях NuSieve GTG.


2% гель | 4% гель |
Фрагменты ДНК разделяли в 2% и 4% агарозных гелях NuSieve GTG в 1´ TBE буфере. Дорожка 1: Hae III – сборная полоса ДНК fX174, дорожка 2: Msp I – сборная полоса ДНК pBR322. Анализ проводили при 5 В/см.
Мы всегда готовы Вам помочь.
При выборе из нескольких вариантов агарозы понимание различий характеристик поможем Вам принять более сознательное решение. Будучи пионерами агарозного электрофореза, мы обобщили огромный опыт и данные как по подбору агарозы, так и по новым задачам и методам. Если у Вас есть ещё вопросы по агарозе или любым другим продуктам – пожалуйста, связывайтесь с нами.
Новости.
Доставка белков непосредственно в клетки.
Откройте для себя доставку заряженных молекул, включая белки, пептиды и другие макромолекулы, в живые клетки с помощью реактива для транспорта белков Passport. Этот совершенно новый пептид (патент находится на рассмотрении), не имеющий аналогов по эффективности, не является цитотоксичным, при добавлении в культуральную среду самостоятельно перемещается вместе с прикреплёнными к нему заряженными молекулы внутрь культивируемых клеток. Эффективность доставки достигает 100% трансфекции культивируемых первичных клеток или клеточных линий.
Реагент для транспорта белков Passport:
Скорость – доставка макромолекул в клетки менее чем через 1-2 часа. Быстрый анализ активности.
Эффективность – доставка биологически активных белков, пептидов и других макромолекул достигает 100% клеток.
Широкое применение – подходит для многих типов клеток, включая первичные клетки, которые сложно трансфецировать.
Отсутствие цитотоксичности – можно анализировать живые клетки.
Рисунок 1. Клетки NIH3T3, трансфецированные реагентом Passport и b-галактозидазой.

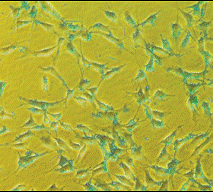
Связывание реагента Passport с заряженными молекулами – простой процесс, в котором принимает участие 2-пиридил-тиогруппа реагента, образующая дисульфидную связь с любым свободным тиолом (цистеином), расположенным на заряженной молекуле. Просто инкубируйте Ваше макромолекулярное соединение с реагентом Passport при комнатной температуре в течение 30 минут, разведите культуральной средой и используйте для культивируемых клеток.
Реагент для транспорта белков Passport поставляется в виде 400 мкг лиофилизированного реагента, достаточного для 40-80 экспериментов в 6-луночных планшетах или >1.000 лунок при трансфекции в 96-луночных. Для проверки и оптимизации экспериментальных параметров поставляется контроль – b-галактозидаза. Чтобы получить более подробную информацию по реагенту Passport, посетите веб-страницу
http://www. /Content/bioscience/Cat/Nav. oid.1032.prodoid. Passport.
