
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
МЕТАБОЛИЗМ
Обмен веществ (метаболизм) - все биохимические процессы - от поступления веществ в организм до выделения конечных продуктов. У животных организмов метаболизм состоит из нескольких последовательных этапов: поступления веществ, пищеварения, всасывания в кровь, проникновения в клетку, превращения их в клетке, образования конечных продуктов, обезвреживания токсических метаболитов и выделения их во внешнюю среду. Аналогичная последовательность процессов обмена прослеживается с некоторыми вариациями у растений и микроорганизмов.
Обмен веществ всегда сопровождается обменом энергии. При распаде веществ, как правило, энергия высвобождается, а при синтезе веществ энергия используется. Обмен веществ и энергии - единый общебиологический процесс.
Функции метаболизма
1. превращение экзогенных веществ в строительные блоки клетки, например: превращение углекислого газа и воды растениями в глюкозу, превращение липидов пищи в глицерол и жирные кислоты и др.;
2. синтез из строительных блоков биомолекул клетки: белков, жиров, полисахаридов, нуклеиновых кислот;
3. обеспечение организма энергией, необходимой для поддержания двух выше перечисленных функций и всех остальных проявлений жизни.
Поступление углерода и азота
Углерод составляет 80% массы липидов, более половины массы белков и около половины - углеводов.
Живые организмы, которые используют углерод углекислого газа для синтеза органических соединений называют автотрофами, а те, которые усваивают углерод только из органических соединений – гетеротрофами.
К автотрофам относятся зеленые растения, сине-зеленые, серные, водородные и другие фотосинтезирующие бактерии. К гетеротрофам - все животные, нефотосинтезирующие клетки растений, некоторые паразитирующие цветковые растения, грибы и микроорганизмы, не способные к фотосинтезу.
Основная масса биогенного азота на земле находится в круговороте (рис. 1):
1) Растения используют нитрат, восстанавливая его до аминокислот. Также они могут усваивать аммоний. Переход неорганического азота (типа нитрата) в органическую форму азота как, например, аминокислоты называется ассимиляцией.
2) Животные организмы и большинство микроорганизмов способны усваивать лишь восстановленный азот, в форме сложных соединений.
Человек нуждается примерно в двадцати сложных азотсодержащих веществах, незаменимых факторах питания. Это незаменимые аминокислоты и витамины.
Животные поедают растения, выделяют азот в виде мочевины и мочевой кислоты или аммиака. Этот процесс перевода азота из органической формы в неорганическую называется минерализацией.
3) Мочевая кислота, мочевина, а также азот, включённый в состав живых существ, после их гибели подвергается аммонификации (разложение содержащих азот сложных соединений с выделением аммиака).
4) Аммиак нитрифицируется бактериями нитрозомонас до нитрита, а бактериями нитробактер - до нитрата. Этот процесс носит название нитрификация.
5) Нитратный азот вновь поглощается растениями или исключается из круговорота денитрифицирующими микроорганизмами, которые используют нитрат в качестве конечного акцептора электронов и протонов в дыхании. Эти микроорганизмы восстанавливают нитрат до молекулярного азота. Процесс – денитрификация.
6) Количество обращающегося в живой природе азота пополняется за счет усвоения его из воздуха азотфиксирующими микроорганизмами родов азотобактер, клостридиум, ризобиум и др. Процесс – азотфиксация.

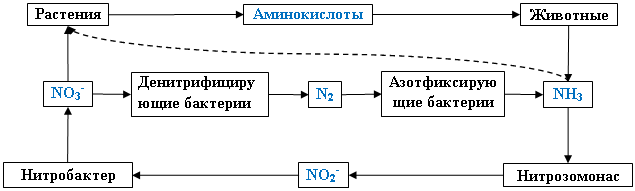
Рисунок 1 – Круговорот азота в природе
Поступление энергии и классификация организмов
Организмы, в зависимости от способа получения энергии делятся на 2 группы:
фототрофы - используют энергию Солнца;
хемотрофы - используют энергию химических связей химических соединений.
Каждая из названых групп делится еще на 2 группы в зависимости от источника углерода и доноров электронов:
литотрофы - используют углерод углекислого газа, а в качестве доноров электронов используют неорганические вещества: воду, сероводород, аммиак, водород, серу и др;
органотрофы - потребляют углерод органических веществ, органические вещества являются и донорами электронов и протонов.
Конечным акцептором электронов в окислительно-восстановительных реакциях у организмов могут быть: кислород, нитраты, сульфаты, органические вещества и др. Если конечным акцептором электронов является кислород, то организмы относят к аэробам. Если не кислород - анаэробам.
В результате вышеприведенных рассуждений, все организмы делятся: фотолитотрофы, фотоорганотрофы, хемолитотрофы и хемоорганотрофы. Среди них могут быть аэробы и анаэробы.
Типичным представителем фотолитотрофов следует считать зеленые клетки высших растений. Они в качестве источника углерода используют углекислый газ, в качестве источника энергии - свет, донором электронов является вода.
К хемоорганотрофам относятся животные. У них источник углерода - органические вещества, источник энергии - окислительно-восстановительные реакции, донором электронов являются также органические соединения. К двум остальным группам: фотоорганотрофам и хемолитотрофам относятся различные представители бактерий.
Катаболизм и анаболизм
Принято различать две стороны обмена веществ - катаболизм и анаболизм.
Катаболизм (диссимиляция) - это расщепление веществ с последующим превращением, происходящим с высвобождением энергии. В этом процессе преобладают реакции гидролиза и окисления.
В катаболизме различают три стадии (рис. 2).
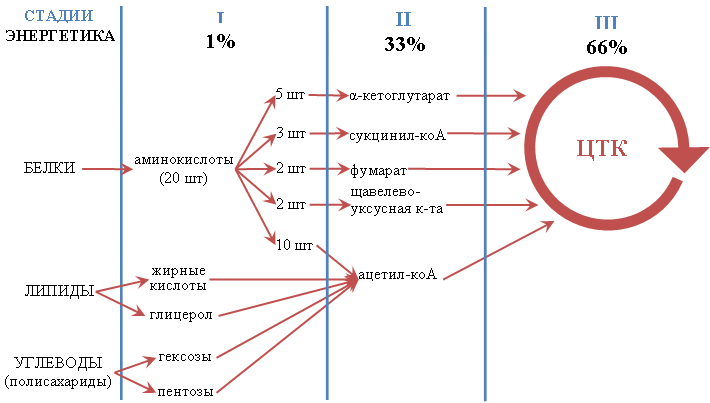

Рисунок 2 – Три стадии катаболизма
На первой стадии макромолекулы распадаются на строительные блоки. Белки распадаются на аминокислоты, полисахариды - на моносахара, липиды - на глицерол и жирные кислоты. На этой стадии высвобождается всего около 1% энергии, заключенной в химических связях веществ.
На второй стадии все многообразие образовавшихся на первой стадии веществ превращается в 5 соединений. Моносахара, жирные кислоты, глицерол и десять аминокислот превращаются в ацетил-КоА. Аргинин, гистидин, пролин, глутаминовая кислота и глутамин - в α-кетоглутарат; аспарагиновая кислота и аспарагин - в щавелево-уксусную кислоту (оксалоацетат); углеродные скелеты изолейцина, валина и метионина - в сукцинил-КоА. Часть углеродного скелета тирозина и фенилаланина превращается в фумарат. На второй стадии высвобождается 33% энергии.
На третьей стадии образовавшиеся на второй стадии 5 соединений окисляются в цикле трикарбоновых кислот. На этой стадии высвобождается 66% энергии химических связей веществ, вступивших в катаболизм. Это самая энергоёмкая стадия катаболизма.
Анаболизм (или ассимиляция) - синтез из простых соединений более сложных с затратой энергии. Здесь преобладают реакции химического синтеза и фотосинтеза.
Анаболизм в общем плане повторяет катаболизм в обратном направлении. Не все реакции катаболизма обратимы, следовательно, пути анаболизма несколько отличаются от путей катаболизма. Так, из 10 реакций гликолиза необратимы три. В цикле трикарбоновых кислот из 9 реакций необратима одна. Обратимые реакции метаболизма называются амфиболическими. Другим отличием катаболизма и анаболизма является различная локализация этих процессов в клетке. Например, катаболизм жирных кислот (β-окисление) протекает в митохондриях, а синтез жирных кислот - в растворимой фракции цитоплазмы.
Локализация метаболических процессов в клетке
Каждая клеточная структура выполняет в метаболизме вполне определенную роль.
1) В цитоплазматической мембране осуществляется направленный перенос из клетки и в клетку.
2) В митохондриях протекают многие важнейшие процессы аэробного окисления веществ: цикл трикарбоновых кислот, окисления жирных кислот. Здесь протекает конечный путь электронов к кислороду: дыхательная цепь и др.
3) В ядре синтезируется ДНК – репликация и РНК - транскрипция.
4) В рибосомах синтезируется белок - трансляция.
5) Лизосомы - ставшие ненужными белки, полисахариды и другие вещества расщепляются до мономеров (гидролиз).
6) В микросомах вещества окисляются кислородом, а в специализированных микросомах - глиоксисомах протекает глиоксилатный цикл.
7) Цитозоль - это отсек цитоплазмы, где сосредоточены ферменты, например, синтеза гликогена, гликолиза, синтеза жирных кислот и др.
8) В аппарате Гольджи идёт синтез липидов, полисахаридов
9) В хлоропластах протекает фотосинтез
Привязанность ферментных систем к определенной структуре клетки носит название компартментализации. Благодаря этому достигается высокая скорость и упорядоченность процессов метаболизма.
Утилизация энергии и её переносчики
Образующаяся в процессе катаболизма энергия используется для различных целей: механическая работа скелетных и гладких мышечных волокон и других двигательных систем, транспорт веществ в клетку и из клетки, биосинтез веществ в процессе анаболизма, теплопродукция и др. Все перечисленные энергозатраты, за исключением теплопродукции, возможны лишь при участии специальных посредников, в качестве которых выступают химические соединения двух типов: нуклеотидные коферменты и нуклеозидтрифосфаты.
1. Нуклеотидные коферменты: НАД, НАДФ, ФМН, ФАД. Важнейшим из них является НАДФ. В процессе катаболизма НАДФ востанавливается в НАДФН2. В реакциях анаболизма НАДФН2 используется как восстановитель, например, в синтезе жирных кислот.
2. Нуклеозидтрифосфаты: АТФ, ГТФ, ЦТФ, УТФ. Наиболее распространённым посредником среди них является аденозинтрифосфат (АТФ). Система АДФ - АТФ - важнейший переносчик энергии в клетке. В физиологических условиях последние две фосфоангидридные связи АТФ заключают в себе значительно больший запас энергии (12 ккал), чем первая связь (3 ккал). Связи, при гидролизе которых высвобождается много энергии в полезной форме, называются макроэргическими. В схемах они обозначаются значком ~ (тильда).
Кроме нуклеозидтрифосфатов, макроэргические связи имеются и в других соединениях. Большинство из них - эфиры фосфорной кислоты, но есть и другие, например, тиоэфир а ацил-коА.
Энергия АТФ используется в самых разнообразных процессах: мышечном сокращении, реакциях синтеза, процессах ионного переноса, передаче генетической информации, т. е. практически везде. АТФ - хорошо растворимое вещество, хорошо проникает через мембраны, легко транспортируется.
Другие нуклеозидтрифосфаты (ГТФ, УТФ, ЦТФ, ТТФ) также участвуют в переносе энергии. Они образуются в процессе катаболизма, например, ГТФ образуется в шестой реакции ЦТК, или из АТФ и соответствующих нуклеозиддифосфатов по уравнению:
АТФ + УДФ àУТФ + АДФ;
АТФ + ЦДФ àЦТФ + АДФ;
АТФ + ГДФà ГТФ + АДФ.
Приведенные реакции катализируются нуклеозиддифосфаткиназами.
Имеется специализация при использовании энергии нуклеозидтрифосфатов для биосинтетических целей. Так, УТФ принимает участие в синтезе полисахаридов, ГТФ - в синтезе белков, ЦТФ в синтезе липидов.
Синтез и распад АТФ
Аденозинтрифосфорная кислота (АТФ) образуется при взаимодействии аденозиндифосфорной кислоты (АДФ) с фосфорной кислотой. Существует два способа синтеза АТФ: субстратное фосфорилирование и окислительное фосфорилирование.
Субстратное фосфорилирование заключается в том, что макроэргическая связь переносится от макроэргических соединений на АДФ. В этом случае на АДФ переносится с субстрата макроэргическая связь вместе с фосфатом. Например: субстратное фосфорилирование происходит в одной из реакций цикла трикарбоновых кислот, где ГТФ синтезируется из ГДФ и фосфата за счет энергии макроэргической тиоэфирной связи в сукцинил-КоА. В результате реакции высвобождается свободный КоА и сукцинат. Образовавшийся в этом примере ГТФ нуклеозидфосфаткиназой в присутствии АДФ легко превращается в АТФ по схеме:
ГТФ + АДФ à ГДФ + АТФ.
Окислительное фосфорилирование - это синтез АТФ из АДФ и Н3Р04, сопряженный с перемещением электронов по дыхательной цепи.
Высвобождение энергии из АТФ происходит двумя путями. Наиболее распространённый путь - ортофосфатное расщепление, когда от АТФ отщепляется ортофосфат:
АТФ + Н2О à АДФ + Ф (Н3РО4)
Такой способ освобождения энергии из АТФ наблюдается при мышечном сокращении, при фосфорилировании глюкозы, фруктозо-6-фосфата и др.
Второй путь высвобождения энергии из АТФ - пирофосфатное расщепление - идет по схеме:
АТФ + Н2О à АМФ + 2Ф (Н4Р2О7)
В этом случае АТФ распадается на АМФ и пирофосфат. Затем пирофосфат расщепляется на 2 остатка фосфорной кислоты с высвобождением энергии. Пирофосфатное расщепление наблюдается в реакциях активации субстрата, например, активации жирных кислот и аминокислот.
Образовавшийся при пирофосфатном расщеплении АМФ может превратиться в АТФ по схеме:
АМФ + АТФà2 АДФ
2 АДФ + 2 Н3РO4à2 АТФ.
Образование и использование АТФ в клетке происходит очень быстро. В организме человека за 1 сутки образуется и распадается 190 кг АТФ. При интенсивной работе еще больше. Однако всего в теле человека не более 50 г АТФ.
Фосфагены
При благоприятных условиях в клетке может создаваться запас энергии. Вещества, способные депонировать энергию в фосфатной связи, называются фосфагенами. Простейшее из них - полиметафосфат (волютин) - обнаруживается в виде хорошо растворимых гранул в клетках микроорганизмов.
 |
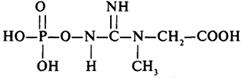
Креатинфосфат
У позвоночных животных таким соединением является креатинфосфат, а у беспозвоночных - аргининфосфат. Эти соединения аккумулируют энергию. Образуются они из АТФ и креатина или аргинина соответственно. В условиях дефицита АТФ фосфагены отдают макроэргический фосфат на АДФ.
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ
Цикл трикарбоновых кислот, цикл лимонной кислоты, цикл Кребса и "метаболический котел" - это все синонимы одного процесса. Цикл трикарбоновых кислот (ЦТК) - последовательность реакций, занимающих центральное место в метаболизме клетки. В метаболическом котле окисляются соединения, образующиеся из липидов, углеводов и белков. Благодаря обратимым реакциям в ЦТК образуются соединения, которые используются в синтезе липидов, углеводов и белков. Большинство ферментов ЦТК локализуется в матриксе митохондрий.
Реакции ЦТК
Цикл трикарбоновых кислот включает 9 реакций (рис.3).
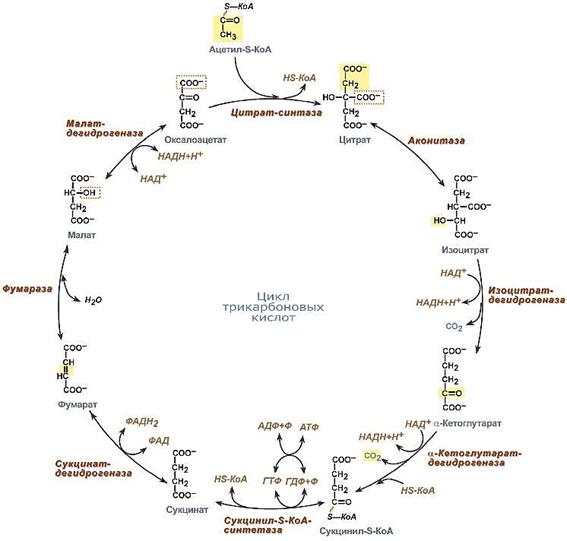
1. Синтез лимонной кислоты. Щавелево-уксусная кислота (оксалоацетат) конденсируется с поступающей в цикл активной уксусной кислотой - ацетил-КоА. Необходимая для синтеза энергия поставляется за счет разрыва тиоэфирной связи в ацетил-КоА. Катализируется реакция цитратсинтазой.
2 и 3. Изомеризация лимонной кислоты в изолимонную. Обе реакции катализируются одним ферментом - аконитазой.
4. Окисление и декарбоксилирование изоцитрата. Катализируется комплексом ферментов, среди которых - НАД-зависимая дегидрогеназа. В результате образуется α-кетоглутарат. При этом НАД восстанавливается до НАДН2. Шестиуглеродная изолимонная кислота теряет один атом углерода в виде СО2.
5. Окислительное декарбоксилирование α-кетоглутарата осуществляется сложным комплексом ферментов, включающим дегидрогеназы. Конечными продуктами здесь являются сукцинил-КоА и СО2. Таким образом, теряется еще один атом углерода и образуется четырёхуглеродная кислота. На этом этапе также высвобождается энергия в виде восстановленного НАДН2.
6. Тиокиназная реакция. Сукцинил-коА гидролитически теряет HS-KoA. Энергия тиоэфирной связи используется для синтеза ГТФ из ГДФ (субстратное фосфорилирование). Высвобождается свободная янтарная кислота - сукцинат. Реакция катализируется тиокиназой.
7. Окисление сукцината осуществляется ФАД-зависимой дегидрогеназой, которая прочно связана с внутренней мембраной митохондрий, в отличие от всех остальных ферментов ЦТК, которые, локализуются в матриксе. ФАД восстанавливается. Образуется фумаровая кислота.
8. Гидратация фумарата катализируется фумаразой. Образуется яблочная кислота - малат.
9. Окисление малата. НАД-зависимая малатдегидрогеназа превращает яблочную в щавелево-уксусную кислоту. Восстанавливается НАД. Цикл замыкается. Щавелевоуксусная кислота готова вновь прореагировать с очередной молекулой ацетил-КоА.
Суммарная реакция ЦТК выглядит следующим образом:
Ацетил-коА+2 Н2O + 3 НАД + ГДФ + Н3РO4 + ФАД à
à 2 СO2 + HS-кoA + 3 НАДН2 + ГТФ + ФАДН2
Таким образом, результатом всех реакций ЦТК является окисление одной молекулы двууглеродной уксусной кислоты до двух молекул СO2 с высвобождением энергии.
Энергетика ЦТК. Окисление трех молекул НАДН2 в дыхательной цепи сопровождается образованием девяти молекул АТФ. Окисление там же одной молекулы ФАДН2 приводит к образованию двух молекул АТФ. Энергетическая ценность ГТФ идентична АТФ. В результате окисления одной молекулы CH3-CO-S-кoA в ЦТК образуется энергия, эквивалентная 12 молекулам АТФ.
В ЦТК имеется два регуляторных фермента: цитратсинтаза (1-ая реакция) и изоцитратдегидрогеназа (2-ая реакция). Оба фермента ингибируются АТФ и НАДН2, изоцитратдегидрогеназа активируется АДФ и НАД, а цитратсинтаэа активируется своим субстратом - щавелево-уксусной кислотой.
Все реакции цикла трикарбоновых кислот, за исключением окислительного декарбоксилирования α-кетоглутарата, обратимы. Следствием этого является то, что ЦТК "работает" не только в процессе катаболизма, но и в анаболизме. Так щавелево-ук-сусная кислота после аминирования становится аспарагиновой кислотой, сукцинил-коА служит предшественником в синтезе гема, α-кетоглутарат после аминирования превращается в глутаминовую кислоту.
Восполняющие реакции ЦТК
1. Ацетил-КоА - основное "топливо" метаболического котла - образуется при окислении жирных кислот, глицерола, углеводов и при окислении большинства аминокислот. Из пирувата путём декарбоксилирования.
2. Другим компонентом, необходимым для запуска ЦТК, является щавелевоуксусная кислота (оксалоацетат). Она образуется из пирувата путем его карбоксилирования:
СН3-СО-СООН + СО2 + АТФ à НOОС-СO-СН2-СOOН + АДФ + Н2РO4
Пируват Оксалоацетат
Также пируват образуется в результате переаминирования или дезаминирования аминокислот (аспарагиновой кислоты) и ряда других реакций.
Исходя из выше изложенного, запустить все реакции ЦТК может одна пировиноградная кислота, поскольку из неё образуется и щавелевоуксусная кислота, и ацетил-КоА. Реакции, ведущие к образованию пирувата, следует также считать восполняющими ЦТК. Это гликолиз, превращение ряда аминокислот в пируват и др.
3. Восполняющими реакциями являются также биохимические процессы, ведущиек образованию других субстратов ЦТК: α-кетоглутарата, сукцинил-коА и фумарата, которые образуются при катаболизме аминокислот.
Цикла трикарбоновых кислот является универсальным механизмом окисления ацетил-коА и других веществ, образующихся из липидов, углеводов и белков. В ЦТК выделяется большое количество энергии. Благодаря амфиболическим реакциям, в нём образуются соединения, необходимые для синтеза белков, углеводов и липидов.
Глиоксилатный цикл
Глиоксилатный цикл - это несколько изменённый ЦТК, свойственный растениям и микроорганизмам. Он протекает в том случае, когда ацетил-КоА используется не только как энергетический, но и как пластический материал. Первые и заключительные реакции у ЦТК и глиоксилатного пути общие. Но при глиоксилатном цикле образовавшийся изоцитрат выводится из митохондрий и поступает в микротельца - глиоксиомы, в которых протекают две реакции:
1) расщепление изоцитрата на глиоксилевую кислоту и сукцинат. Образовавшийся сукцинат в дальнейших реакциях цикла участия не принимает.
2) конденсация глиоксилевой кислоты и ацетил-коА с образованием малата. Малат вновь поступает в митохондрии и окисляется, как в ЦТК, в щавелево-уксусную кислоту.
Таким образом, в реакциях глиоксилатного цикла из двух молекул ацетил-коА образуется одна четырёхуглеродная молекула янтарной кислоты. При этом на две молекулы ацетил-КоА образуется всего одна НАДН2. То есть глиоксилатный цикл не столько катаболический процесс, сколько анаболический, так как в его последовательных реакциях из ацетил-КоА образуется более сложное соединение - янтарная кислота.
Биологическое окисление. Дыхательная цепь
Биологическое окисление - это совокупность окислительно-восстановительных реакций, протекающих в живых организмах. Есть два способа окисления веществ в клетке. Первый начинается с отщепления с помощью НАД - или ФАД-зависимых дегидрогеназ электронов и протонов от субстрата. В дальнейшем протоны связываются кислородом. Этот способ позволяет аккумулировать высвобождающуюся в процессе окисления энергию в виде АТФ. Он преобладает в клетке.
Второй способ - присоединение кислорода к субстрату, он катализируется оксигеназами. В этом случае энергия высвобождается в виде тепла.
Заключительным этапом в первом способе окисления является дыхательная цепь. Дыхательная цепь (электрон-транспортная цепь дыхания, ЭТЦ) - система связанных белков и переносчиков электронов и протонов. ЭТЦ позволяет запасти энергию, выделяющуюся в ходе окисления НАДН2 и ФАДН2. Энергия запасается в форме разности потенциалов протонов на мембране за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану. Высвобожденная энергия идёт на синтез макроэргических связей АТФ. Конечным акцептором электронов и протонов при аэробном дыхании является кислород. Локализуется дыхательная цепь на внутренней мембране митохондрий.
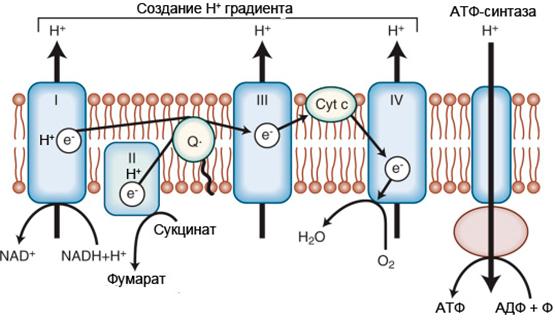
Рисунок 4 - Электрон-транспортная цепь митохондрий
Ферменты и вещества дыхательной цепи
Все ферменты дыхательной цепи относятся к оксидоредуктазам и являются сложными белками.
Пиридинзависимые дегидрогеназы в качестве простетической группы содержат НАД. Эти ферменты осуществляют перенос электронов и протонов (водорода) от субстрата.
Флавинзазисимые дегидрогеназы в качестве кофермента содержат ФАД или ФМН. К флавинзависимым дегидрогеназам относятся сукцинатдегидрогеназа.
Убихинон (кофермент Q) - небелковый переносчик электронов и протонов. Структура убихинона включает хинон и изопреноидную цепь. Присоединяя два водорода, хинон превращается в гидрохинон.
Цитохромы - переносчики электронов, по строению это гемопротеины. Цитохромы b и с имеют небольшие различия в строении радикалов гема. Цитохромы переносят электроны, которые присоединяются к железу гема. Железо при этом меняет валентность Fe3+ à Fe2+. Каждый цитохром переносит один электрон. Протоны цитохромами не транспортируются.
Цитохромоксидаза (Сu2+) - единственный фермент в цитохромной системе, способный отдавать электроны на кислород.
Железосерные белки обнаружены в составе мембран митохондрий. Железо в железосерных белках (FeS-комплексов) удерживается с помощью серы (SH-групп) аминокислоты цистеина. Эти белки участвуют в окислительно-восстановительных процессах и являются компонентами дыхательной цепи. Наиболее известный белок из этого семейства, называется ферредоксином.
Последовательность окисления в ЭТЦ
Перенос электронов и протонов по ЭТЦ начинается с окисления НАДН2 или ФАДН2 дегидрогеназами (рис. 4).
НАДН2 окисляется НАДН2 дегидрогеназой (комплекс I), которая отбирает у него два электрона и протона и переносит их на растворимый в липидах убихинон. Во время этого процесса комплекс I перекачивает протоны из матрикса в межмембранное пространство митохондрий.
ФАДН2 окисляется сукцинатдегидрогеназой (комплекс II). При этом процессе протоны не перекачиваются через мембрану, но убихинон получает дополнительные электроны и протоны.
Восстановленный убихинон (гидрохинон) отдаёт электроны на цитохромный комплекс (комплекс III), а он в свою очередь переносит электроны на два водорастворимых цитохрома с, расположенных на внутренней мембране митохондрии. При этом процессе также переносятся протоны убихинона, которые перекачиваются комплексом.
Конечным акцептором электронов в дыхательной цепи является кислород. Передачу электронов на кислород осуществляет цитохромоксидаза (комплекс). При этом комплексом IV перекачиваются протоны в межмембранное пространство. Цитохромоксидаза отдаёт электроны на кислород и активирует его. В заключение активированный кислород с двумя протонами образует воду.
В итоге при переносе электронов и протонов в ЭТЦ образуется протонный потенциал на внутренней мембране митохондрий.
В рассмотренной дыхательной цепи конечным акцептором электронов и протонов является кислород. У некоторых микроорганизмов конечным акцептором протонов и электронов выступает нитрат, который восстанавливается до азота или даже до аммиака. Дыхательная цепь в анаэробном "нитратном" дыхании отличается от цепи аэробного (кислородного) дыхания только последним ферментом. Вместо цитохромоксидазы последним ферментом цепи в этом типе дыхания является нитратоксидаза. У некоторых микроорганизмов обнаружено "сульфатное " дыхание, в котором сульфат восстанавливается до серы или до сероводорода.
Окислительное фосфорилирование
Общепризнаной теорией синтеза АТФ в ЭТЦ является теория хемиоосмотического сопряжения, выдвинутая П. Митчелом. Сущность ее следующая: процесс окислительного фосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий — протонной АТФ-синтазой. В процессе транспорта электронов по дыхательной цепи образуется градиент ионов водорода по разные стороны внутренней мембраны митохондрий. Возникший градиент концентрации заставляет протоны проходить через АТФ-синтазу на ту сторону мембраны, где концентрация протонов ниже. На АТФ-синтазе удерживаются АДФ и Фн. Проход протонов через АТФ-синтазу приводит к её конформационным изменениям и соответственно взаимодействию АДФ и Фн. Реакция синтеза такова:
АДФ + Фн → АТФ + H2O
Это наиболее распространенный способ окисления субстрата в клетках, осуществляемый путем дегидрирования. Эффективность этого способа характеризуется высоким коэффициентом полезного действия. Примерно 40% энергии, заключенной в химической связи окисляемых веществ, концентрируется в макроэргических связях АТФ, а остальная энергия рассеивается в виде тепла.
Оксигеназы
Второй способ окисления веществ - присоединение кислорода к субстрату. Такое окисление имеет место в микросомах и катализируется оксигеназами.
В процессе окисления к субстрату может присоединяться атом кислорода, и образуется гидроксильная группа. Ферменты, осуществляющие эти реакции, называются монооксигеназами. В других случаях к субстрату присоединяется молекула кислорода. Ферменты, катализирующие присоединение молекулы кислорода, называются диоксигеназами.
Оксигеназы - сложные ферменты, содержащие гем, рибофлавин или негеминовое железо. Такое окисление не сопровождается синтезом АТФ. Это редкий способ окисления, не более 10% кислорода, поступающего в организм, используется оксигеназами. 90% кислорода восстанавливается с участием цитохромоксидазы, т. е. в дыхательной цепи.
Оксигеназы участвуют при генерации тепла, для обезвреживания токсических веществ.
