
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ЗОЛОТУХИН Вадим Викторович
Система и фауногенез коконопрядов
мировой фауны (Lepidoptera: Lasiocampidae)
03.02.05 – энтомология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
доктора биологических наук
Санкт-Петербург, 2011
Работа выполнена на кафедре зоологии ФГБОУ ВПО
«Ульяновский государственный педагогический университет имени »
Научный консультант:
доктор биологических наук (ЗИН РАН)
Официальные оппоненты:
доктор биологических наук (СПбГУ),
доктор биологических наук (ИПЭЭ РАН),
доктор биологических наук, профессор (СПбГЛТА)
Ведущая организация:
Саратовский государственный университет им. .
Защита состоится «13» октября 2011 в 16:00 часов на заседании совета Д 212.232.08 по защите докторских и кандидатских диссертаций при Санкт-Петербургском государственном университете Санкт-Петербург, Университетская наб, д. 7/9, ауд. 133.
С диссертацией можно ознакомиться в научной библиотеке им. Санкт-Петербургского государственного университета.
Автореферат разослан «……....» 2011 г.
Ученый секретарь
диссертационного совета,
кандидат биологических наук
e-mail: katrin. *****@***ru
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследований. Коконопряды (Lasiocampidae) – семейство шелкопрядообразных чешуекрылых (Bombycomorpha), иногда выделяемое в самостоятельное надсемейство Lasiocampoidea. Объем семейства до настоящего времени не выяснен; ежегодно описывается до 100 новых видов, в том числе и из таких хорошо исследованных регионов, как страны Юго-Восточной Азии или острова Тихого океана. В некоторых родах до 60% видов остаются неописанными, а до 20% используемых видовых эпитетов должны быть заменены старейшими валидными названиями. Общепринятой системы группы также не существует; разные авторы значительно расходятся в понимании объема и взаимосвязей между родами и обычно используют слабо обоснованную и уже устаревшую систему 1927 года, предложенную Х. Ауривиллиусом. Начатая ревизия коконопрядов мировой фауны показала, что большинство афро - и неотропических родов представляют собой сложные пара - или полифилетические комплексы, включающие несколько самостоятельных эволюционных ветвей. По-прежнему не разработаны вопросы фауногенеза коконопрядов, изучение которых может способствовать пониманию общих закономерностей эволюции группы.
Изучение коконопрядов актуально и в чисто практическом аспекте, поскольку они являются одними из основных дефолиаторов многих лиственных и хвойных пород. Кроме того, гусеницы Gloveria и Quadrina вызывают серьезные кожные раздражения, а пыль коконов Malacosoma аллергенна (Stehr, 1987). Некоторые виды коконопрядов используют для получения шелка, более прочного и долговечного, чем шелк тутового шелкопряда. Коконы некоторых видов в различных мировых культурах используются также для изготовления украшений и являются своеобразной валютой (Peigler, 1993, 1994), а куколки и гусеницы могут использоваться в пищу.
Цель и задачи исследования. Целью данной работы является изучение таксономической структуры семейства и вопросов его фауногенеза. Для этого были поставлены следующие задачи:
1) провести обзор существующих систем коконопрядов мировой фауны;
2) на основе результатов сравнительно-морфологического, функционально-морфоло-гического и биохимического анализов модернизировать систему семейства в объеме мировой фауны;
3) проанализировать видовой и родовой состав фаун коконопрядов всех континентов, установить уровень их эндемизма;
4) определить вероятные пути фауногенеза группы на каждом из континентов;
5) изучить видовой и родовой состав коконопрядов крупных островов Старого Света и выявить особенности становления островных фаун.
Основные положения, выносимые на защиту:
1. На основе анализа признаков внешней морфологии и генитального аппарата создана оригинальная классификация семейства Lasiocampidae, протестированная с помощью молекулярно-генетических методов. В составе семейства выделены пять крупных монофилетических групп, рассматриваемых в ранге подсемейств; самое крупное подсемейство Lasiocampinae Harris, 1841 включает 9 триб.
2. Основные события фауногенеза семейства приходятся на позднемеловой период и связаны с активными рельефообразовательными процессами этого времени. Специализированные эремиальные роды коконопрядов возникли относительно недавно, предположительно в плейстоцене, в связи с крайней аридизацией соответствующих территорий.
3. Фауны коконопрядов отдельных континентов слагаются как за счет видов автохтонного происхождения, так и за счет вселенцев. Неарктическая и неотропическая фауны коконопрядов формируются в результате 3 основных волн миграции предковых форм через Берингию из Евразии. Наиболее богаты эндемиками крупные океанические острова и континенты, на которых развитие местных фаун шло в условиях изоляции.
Научная новизна. Ревизована система семейства Lasiocampidae, ее основные положения опубликованы (Zolotuhin, 1998, 2009). В ходе проделанной работы определен таксономический состав семейства в рамках Старого Света; в частности, выявлен видовой состав ряда региональных фаун (Кавказа, европейской части бывшего СССР, Дальнего Востока России и Российской Федерации в целом, республик Средней Азии, Вьетнама, Филиппин, Таиланда, Сулавези, Флореса, Лаоса, Непала). Установлен видовой состав всех родов Палеарктической и Индо-Малайской фаун и частично ревизована Афротропическая фауна. Описаны в качестве новых для науки 139 видов и 43 подвида, а также 27 родов; подготовлены к описанию 2 новые трибы. Установлено более 100 новых синонимов видового и родового ранга.
Определено положение Lasiocampidae среди родственных семейств, и предложенная гипотеза проверена молекулярными методами. Предложены пути становления фаун различных материков и проанализирован фауногенез некоторых таксономических (Chionopsychinae, Chondrosteginae, Selenepherini и др.) и экологических (хвоегрызущие и эремиальные коконопряды) групп.
Теоретическая и практическая значимость. Результаты проведенной ревизии дают основу для последующих обобщений как в части построения надродовой системы семейства, так и определения его места в пределах бомбикоидного комплекса (Bombycoidea sensu lato). Разработка вопросов становления фаун коконопрядов различных материков вносит вклад в изучение исторического развития наземной биоты. Составленные региональные списки видов могут быть использованы службами лесного и сельского хозяйства различных стран для мониторинга вредных видов, а также природоохранными организациями для определения редких и требующих особого внимания видов. Переданные в Академию Наук Вьетнама списки хвоегрызущих видов (Zolotuhin, Ta, Tran, 2009) уже официально используются для корректировок сроков обработок сосновых посадок. Основные результаты и положения работы включены в курсы отдельных дисциплин, читаемых в УлГПУ, Самарском государственном университете и Саратовском государственном университете.
Апробация работы и публикации. Результаты работы докладывались на ежегодных конференциях преподавателей УлГПУ, Конгрессе по проблемам энтомологии Средней Европы (Жигулевск, 1993), X съезде Всесоюзного (Ленинград, 1989) и XIII съезде Русского энтомологического общества (Краснодар, 2007), XX международном энтомологическом конгрессе (Фиренце, 1996), XV Европейском лепидоптерологическом конгрессе (Эркнер, 2007), Зоологических чтениях Баварии (Мюнхен, 2007) и Конференции по биоресурсам (Ханой, 2009).
По теме диссертации опубликовано 84 работы, из которых 4 – монографии и коллективные монографии, 9 опубликованы в списке изданий, рекомендованных ВАК, из 71 прочих – 11 русскоязычные, 54 в иностранных изданиях и 6 как тезисы.
Структура и объем работы. Диссертация состоит из Введения, 7 глав, Выводов, Списка литературы и Приложения. Список цитируемой литературы включает 262 источника, среди которых 202 на иностранных языках. Основной текст диссертации изложен на 202 страницах, включает 23 таблицы и 34 рисунка. В Приложение вынесены база данных представителей семейства, отобранных для анализа ДНК; матрица нуклеотидных последовательностей фрагмента гена EF-1α, использованных в работе; каталог видов коконопрядов мировой фауны и перечень выделов для кластерного анализа распространения коконопрядов.
Благодарности. Выражаю свою глубочайшую благодарность всем родным, друзьям и коллегам, без дружеской поддержки, разнообразной помощи и ценных советов которых данная работа была бы невозможной.
Глава 1. Материал и методика исследований
1.1. Материал. В основе работы лежат собственные сборы коконопрядов и близких групп чешуекрылых, выполненные в ходе полевых исследований в Европейской части России (1984–2008 гг.), Средней Азии (1989–1994), на Дальнем Востоке РФ, включая Сахалин и Кунашир (1991–1993), в Германии (1995), Турции (1995), Таиланде (2005), Вьетнаме (2006, 2008, 2009), ЮАР (2007), Демократической Республике Конго (2008). Обработаны материалы большинства ведущих научных учреждений мира и крупных частных коллекций России и Европы. За время исследований было обработано более 600 000 экземпляров и изготовлено более 5 900 препаратов различных морфологических структур, в том числе преимагинальных стадий – микроскульптуры хориона яиц, хетотаксии гусениц и куколок.
1.2. Методика. Методика сбора и выращивания живого материала была традиционной. Изготовление генитальных препаратов осуществлялось по общепринятой методике, с использованием в качестве уплотнителя Эупарала; инвертирование везики самца проводилось с помощью тонких инсулиновых шприцев. Электронно-микроскопические исследования выполнены на базе Zoologisches Forschungsinstitut und Museum Alexander Koenig, Bonn, с использованием сканирующего электронного микроскопа HITASHI S 450.
Материал для изучения ДНК фиксировался в 96% этаноле; дальнейшая его обработка проводилась на базе специализированных лабораторий (СарГУ и Canadian Centre for DNA Barcoding, Canada). Всего исследовано 2090 образцов.
Глава 2. Морфологический обзор семейства
Дан обзор морфологических признаков, служащих для целей диагностики и построения системы, рассмотренный по следующим разделам: Взрослое насекомое; Генитальный аппарат; Яйцо; Гусеница; Куколка и кокон. Выделено несколько диагностических черт семейства, часть которых являются его аутапоморфиями, а другие должны быть рассмотрены как гомоплазии: боковая или нижняя часть первого членика щупиков несет особую сенсорную структуру, напоминающая хетозему; пилиферы отсутствуют; в переднем крыле дистальная часть радиального ствола тесно сближена с субкостальным стволом; ареола отсутствует; CuA2 переднего крыла отходит от основания радиокубитальной ячейки на значительном удалении от основания М3; соединение крыльев осуществляется по амплексиформному механизму (Minet, 1998) с участием особой мелкошипиковой зоны переднего края заднего крыла, называемой спинареей; мускульный набор генитального аппарата самцов сокращен с исходных 8 до 2-3 пар мышц с полным выпадением m1, m3, m20 у большинства групп; хетотаксия гусениц первого возраста с обильным развитием подпервичных и вторичных щетинок; мандибулы гусениц с несколькими вторичными щетинками; брюшные ноги гусениц изнутри с двумя зонами отчетливой склеротизации, разделенными вертикальной мембраной; футляр хоботка у куколки сильно укорочен или полностью отсутствует.
Вместе с тем, семейство обладает несколькими явными плезиоморфиями: радио-кубитальная ячейка крыльев со следами медиального ствола; на крыльях часто сохраняется рудиментарная А1; гусеницы первого возраста несут полный или почти полный венец крючков на подошве брюшных ног; хетотаксия гусениц исходного типа без значимых модификаций; дендробионтность у подавляющего большинства родов; наличие эпикраниального шва у куколок; кокон (у архаичных родов) тесно связан с почвой и инкрустирован ее частицами.
Глава 3. Биология
В главе рассматриваются образ жизни коконопрядов, пищевые связи гусениц и их региональные особенности. Отсутствие доказанных случаев монофагии и редкость узкой олигофагии делают затруднительным анализ сопряженной эволюции коконопрядов и их кормовых растений. Можно лишь констатировать, что эволюция группы была преимущественно связана с освоением покрытосеменных, и прежде всего двудольных, растений. При проникновении в новые для них биотопы, коконопряды переходили к использованию и новых для них кормовых растений. В условиях аридного климата ими становились ксерофитные злаки и осоки или специализированные формы бобовых. В зависимости от условий обитания и исходных кормовых предпочтений развивалась полифагия на травянистых растениях (все Chondrostegini) или узкая олигофагия/монофагия на бобовых (Selenepherini аридных ландшафтов) или миртовых (австралийские представители). В горных местностях наиболее легко доступным кормовым субстратом для коконопрядов оказались голосеменные растения; приспособление к ним привело к формированию специализированных групп семейства, не способных использовать в пищу лиственные породы.
Глава 4. Система коконопрядов
4.1. Обзор имеющихся систем семейства. Самая ранняя попытка классифицировать коконопрядов была предпринята Дж. Таттом (Tutt, 1902), который рассмотрел представителей Палеарктической фауны в составе 3 отдельных семейств Chondrostegidae, Lachneidae и Eutrichidae, и предложил очень дробный вариант системы, включающей 23 трибы. Ауривиллиус (Aurivillius, 1927), основываясь на особенностях расположения субкостальной жилки и радиального ствола на заднем крыле, предложил разделение семейства на 7 подсемейств: Chondrosteginae, Malacosomatinae, Archaeopachinae, Gastropachinae, Chionopsychinae, Lasiocampinae и Gonometinae. Все последующие предлагавшиеся варианты системы в той или иной степени опирались на схему, предложенную Ауривиллиусом.
4.2. Филогенетический анализ. Для тестирования выдвинутых ранее гипотез о взаимоотношениях различных родов был проведен филогенетический анализ семейства Lasiocampidae на основе изучения нуклеотидных последовательностей генов EF-1α и СО1 представителей его европейской, индо-малайской и афротропической фаун.
4.2.1. Объекты и методика исследования. Для 49 видов коконопрядов (64 экз.) с территории Евразии и Африки были установлены нуклеотидные последовательности гена EF-1α. Также использованы 6 образцов, депонированных в GenBank. В качестве внешней группы выбран Lemonia dumi (семейство Lemoniidae).
Анализ нуклеотидных последовательностей проведен на автоматическом секвенаторе Beckman в лаборатории молекулярной биологии СарГУ. Коррекция полученных последовательностей выполнена в программе BioEdit Sequence Alignment Editor. Для множественного выравнивания использована программа ClustalW. Построение филогенетических деревьев произведено с использованиием методов максимального правдоподобия, ML, минимальной эволюции, ME, максимальной парсимонии, MP и Байеса, BI в пакетах прикладных программ MEGA 4, MEGA 5 и MrBayes 3.1.2.
Матрица нуклеотидных последовательностей составлена из фрагментов длиной от 585 до 1019 п. н. Всего в анализе были использованы 72 нуклеотидные последовательности. На исследуемом участке гена EF-1α с 164 по 1182 п. н. выявлено 345 вариабельных сайтов, что составило 34%. Доля парсимониально значимых сайтов составила 28% (287 сайтов). Среднее отношение транзиций к трансверсиям составило 2,57.
4.2.2. Исследование генетического родства представителей Lasiocampidae.
Топологии полученных кладограмм ML, ME, MP, и BI в целом оказались сходными и в основных узлах имели высокие значения поддержки (более 50% бутстреп-поддержки для методов ME, MP и ML и постериорная вероятность более 0.5 для BI). Метод Байеса дал высокие значения поддержки (0,99-1) для большинства клад (рис. 1), поэтому в качестве оптимального было выбрано именно дерево BI. По результатам проведенного анализа можно выделить несколько четко очерченных групп коконопрядов.
Две ветви, представленные родами Malacosoma и Chionopsyche, во всех вариантах дендрограмм оказываются базальными и обособленными, что позволяет рассматриватьотреть их в качестве отдельных подсемейств Malacosominae и Chionopsychinae. Остальные роды образуют особую кладу с высоким уровнем поддержки (66-98%), для которой существует валидное название Lasiocampinae. Все подчиненные ей крупные клады можно рассматривать в ранге триб Selenepherini, Gastropachini, Pinarini и Argudini tribus nova (Arguda + Syrastrena (+ Hallicarnia).
Род Euthrix представляется на настоящий момент парафилетическим. Вероятно, в дальнейшем будет целесообразно выделить вид E. isocyma в отдельный монотипический род.
Группа родов, объединяемая здесь в составе Argudini (Arguda, Syrastrena, Odontocraspis и Leipoxais), ранее сближалась с Odonestis в рамках трибы Odonestini. Однако все построенные дендрограммы широко разводят эти роды, которые, вероятно,
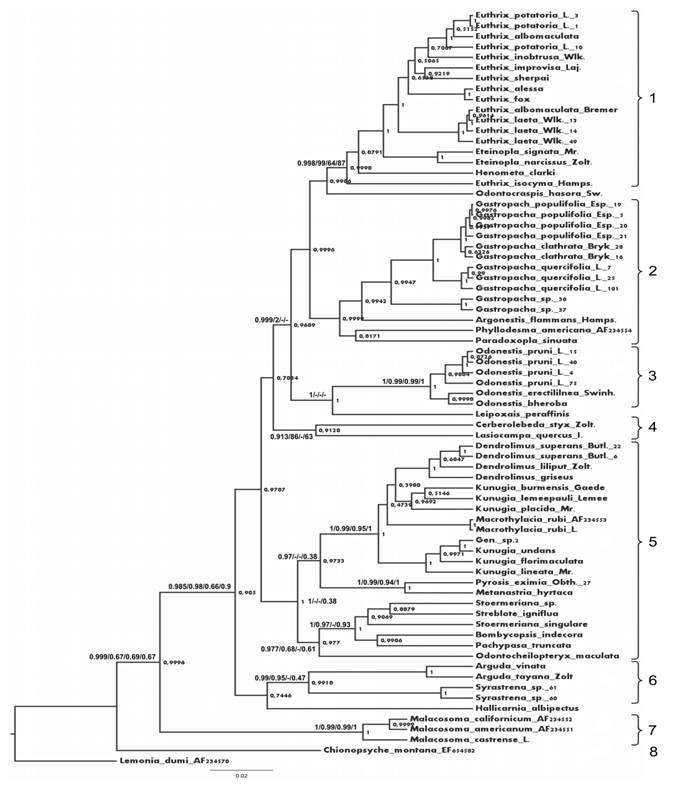
Рис. 1. Схема филогенетических отношений родов коконопрядов на основании изучения полиморфизма нуклеотидных последовательностей гена EF-1α с использованием метода Байеса.
Значения поддержки (постериорная вероятность для метода BI и бутстреп-поддержка для методoв ML, ME и MP) приведены у основания ветвей в последовательности BI/ME/MP/ML; если в анализе клада не выявлялась, то вместо значения поддержки указан прочерк. Цифрами обозначены: 1 – Selenepherini; 2 – Gastropachini; 3 – Odonestini; 4 – Lasiocampini; 5 – Pinarini; 6 – Argudini; 7 – Malacosomini: Malacosominae; 8 – Chionopsychini: Chionopsychinae; Lemonia dumi – внешняя группа.
не являются столь близкими. Роды Odontocraspis и Leipoxais в каждом из 4 вариантов анализа располагаются в различных кладах и не включаются ни в одну трибу. Однако морфологически оба рода близки к представителям трибы Argudini и, вероятно, должны быть отнесены именно к ней.
Обособленную позицию занял род Hallicarnia, ранее провизорно сближавшийся с представителями ветви Argudini; такая точка зрения не поддержана молекулярно-генетическим анализом, и положение этого рода в системе семейства требует уточнения. Неопределенным остается и положение рода Macrothylacia. Признаки строения гениталий и особенности морфологии гусениц сближают этот род с Lasiocampini, но его включение в состав последней трибы пока несколько провизорно.
Представители рода Stoermeriana оказались разнесенными на схеме в удаленные ветви одной клады, что неудивительно, поскольку этот пара - или полифилетический род распадается на несколько морфологически четко очерченных видовых групп.
Несмотря на то, что в BI род Argonestis включается в Gastropachini, морфологически он явно близок к Odonestis. Такое родство с высокой бутстреп-поддержкой отражено также в ML, что позволяет рассматривать его в составе Odonestini.
4.3. Обсуждение.
Полученная кладограмма, в целом, поддерживает идею о синонимизации Pinarinae с Lasiocampinae, рассматриваемых как отдельные трибы Pinarini и Lasiocampini. Вместе с тем, морфологически представители обеих этих групп хорошо отличаются друг от друга, обладая собственными аутапоморфиями. С другой стороны, большинство родов, традиционно рассматривающихся различными авторами в составе Lasiocampini (Anadiasia, Arguda, Euthrix, Pachypasa и др.), в действительности являются типичными членами пинароидной линии. Это подтверждается как признаками строения генитальных структур, так и особенностями развития преимагинальных стадий.
Нет оснований поддержать ранг подсемейства для неотропических Macromphaliinae, которые явно близкородственны Poecilocampinae. Для последних отмечено 28 (у Trichiura crataegi) или 36 (у Poecilocampa populi) хромосом в гаплоидном наборе, против типичных 30-31 хромосом у прочих родов семейства. Недавние исследования показали, что Macromphaliinae также обладают гаплоидным набором, равным 28. Наличие такой важной синапоморфии служит основанием для понижения статуса подсемейства Macromphaliinae до трибы Macromphaliini, stat. nov.
До настоящего времени молекулярно-генетические исследования не затронули виды подсемейства Chondrosteginae, но крайняя специализация его представителей позволяет сохранить за группой статус подсемейства.
На основании сопоставления результатов морфологического и молекулярно-генетического анализа нами предлагается следующий вариант системы коконопрядов мировой фауны:
Lasiocampidae Harris, 1841
1. Chondrosteginae Tutt, 1902
= Chondrostegidae Tutt, 1902
1.1. Chondrostegini Tutt, 1902
2. Poecilocampinae Tutt, 1902
= Trichiurinae Tutt, 1902
= Macromphaliinae Franclemont, 1973
2.1 Poecilocampini Tutt, 1902
= Trichiuridi Tutt, 1902
2.2. Macromphaliini Franclemont, 1973
3. Chionopsychinae Aurivillius, 1927
3.1. Chionopsychini Aurivillius, 1927
4. Malacosominae Tutt, 1902
= Malacosomatinae Aurivillius, 1927
4.1. Malacosomini Tutt, 1902.
5. Lasiocampinae Harris, 1841
= Lachneidae sensu Tutt, 1902
= Eutrichidae sensu Tutt, 1902
= Pachygastriinae Tutt, 1902
= Metanastriinae Tutt, 1902 (partim)
= Pinarinae Kirby, 1892
= Eutrichidae Tutt, 1902 (partim)
= Diplurinae Tutt, 1902
= Dendroliminae Tutt, 1902
= Pachypasinae Tutt, 1902
= Trabaliinae Tutt, 1902
= Chileninae Tutt, 1902
= Cosmotrichinae Tutt, 1902
= Eutrichinae Tutt, 1902
= Nadiasinae Tutt, 1902
= Gastropachinae Newm. & Dyar, 1893
= Gonometinae Aurivillius, 1927
5.1. Lachneini Grote, 1888
5.2. Lasiocampini Harris, 1841
= Pachygastriidi Tutt, 1902
= Lambessidi Tutt, 1902
= Metanastriidi Tutt, 1Macrothylaciini Tutt, 1902
5.4. Selenepherini Tutt, 1902
= Dipluridi Tutt, 1902
= Chilenidi Tutt, 1902
= Cosmotrichidi Tutt, 1902
= Routledgiidi Tutt, 1902
= Nadiasidi Tutt, 1902
5.5. Trabaliini Tutt, 1902.
5.6. Pinarini Kirby, 1892.
= Paralebedidi Tutt, 1902
= Dendrolimidi Tutt, 1902
= Pachypasidi Tutt, 1902
= Bhimidi Tutt, 1902
5.7. Gastropachini Newm. & Dyar, 1893
= Eutrichidi Tutt, 1902
5.8 Odonestini Tutt, 1902
5.9. Argudini trib. nov.
Диагнозы всех надродовых таксонов с указанием типовых родов даны в тексте диссертационной работы.
Глава 5. Особенности распределения и фауногенеза коконопрядов
по материкам
5.1. Видовое разнообразие коконопрядов различных зоогеографических областей рассмотрено на основе схемы, предложенной (2002). По фауне коконопрядов, как и медведиц (Дубатолов, 2007), Чилийско-Патагонская область не обнаруживает отличий от Неотропической области ни на видовом, ни на родовом уровне и потому рассматривается в составе Неотропического, а не Нотогейского царства, а Папуасская область, близкая к Австралийской, переносится из Палеотропического царства в Нотогейское.
5.1.1. Голарктическое царство. Известно 56 палеарктических и 17 неарктических родов, а также 389 и 66 видов, соответственно. Общими для Неарктики и Палеарктики являются роды Phyllodesma и Malacosoma, а общие виды для них не известны. Уровень видового эндемизма высок, и около 90% видов не выходят за пределы царства. Для Голарктического царства известен лишь один эндемичный таксон надродового ранга – триба Malacosomini; вероятно, триба Lasiocampini также строго эндемична для хорона.
Бореальная область сильно обеднена родами (28) и видами (75), которые связаны в основном с хвойными и мелколиственными лесами. Доля аркто-альпийских видов ничтожна; единственный вид с аркто-альпийским ареалом (Eriogaster arbusculae) эндемичен для Палеарктики.
Область Древнего Средиземья. Известно только 26 родов с высоким уровнем эндемизма (9); среди эндемиков Psilogaster, Pachypasa, Gufria. Также высок уровень видового эндемизма: 92 вида из 112 известных (82%) – эндемики; многие с точечными ареалами (Malacosoma laurae, M. mixtum, M. squalorum, Macrothylacia digramma, Eriogaster philippsi, Phyllodesma kermesifolia, Ph. suberifolia, Ph. priapus и др.).
Восточноазиатская область. В целом из области известны 41 род и 210 видов с уровнем эндемизма соответственно 24,4% и 52,4%. Среди эндемичных родов Lenodora, Cosmeptera, Eteinopla, Pachypasoides, Similodora, Somadasys, Syrastrenopsis, Notogroma, Amurilla. Условно субэндемичны хозяйственно важные Cosmotriche и Dendrolimus. Своеобразие хорона объясняется его разнообразными ландшафтами и складчатостью в виде горных систем, среди которых особо значимыми являются Гималаи.
Сонорская область. Фауна области насчитывает 17 родов и 66 видов, причем 2 рода общие с фауной Палеарктики (Malacosoma и Phyllodesmа). Эндемичных родов всего 4 (Apotolype, Hypopacha, Heteropacha и Edwardsimemna), но большинство видов эндемичны для хорона, и в Неотропику проникает лишь 6-8 видов.
5.1.2. Палеотропическое царство. Наиболее богатое родами и видами царство, различные области которого имеют специфическую фауну коконопрядов. При этом часть видов в ходе эволюции проникали на территории смежных областей, сформировав современный сложный таксономический состав каждого хорона. Общих видов, распространенных по всей территории царства, нет, общих рода 2 (Streblote и Estigena). Эндемичных надродовых таксонов нет.
Ориентальная (Индо-Малайская) область. Высок уровень видового эндемизма из-за большого количества океанических островов. Известно 32 рода (2 эндемичных – Voracia и Eleganteda) и 208 видов, среди которых более 100 эндемичных. В родах Trabala, Paralebeda, Lebeda, Metanastria, Estigena, Arguda наблюдается целый веер аллопатрических островных видов.
Афротропическая область. Отмечено 77 родов и около 600 видов. Все виды эндемичны. Из надродовых таксонов, помимо эндемичных Chionopsychinae, для области известны автохтонные трибы Chondrostegini, Pinarini и Selenepherini и проникшие из Индо-Малайской области Trabaliini и Gastropachini. Ареал большинства родов жестко ограничен на севере пустыней Сахара, и лишь единичные специализированные виды (из родов Braura, Beralade, Sena, Streblote, Anadiasa) проникают в ее оазисы.
Мадагаскарская область. Для острова отмечено 27 родов и 95 видов коконопрядов; при этом 25 родов эндемичны, и лишь Lechriolepsis и Odontocheilopteryx представлены также в тропической Африке. Впрочем, родство африканских и мадагаскарских представителей рода не бесспорно и отражено лишь в сильном внешнем сходстве. Все виды эндемичны.
5.1.3. Неотропическое царство. Фауна коконопрядов насчитывает 496 видов из 16 родов, причем ее ядро составляют виды 6 близких родов (30 видов Artace, 351 вид Euglyphus, 16 видов Macromphalia, 31 вид Nesara, 29 видов Titya и 98 Tolype), относящихся к трибе Macromphaliini.
Неотропическая область. Отмечено около 450 видов из 16 родов; 1 род (Malacosoma) проникает из Сонорской области. Большинство видов эндемичны, но эндемичных родов нет; родовая система для хорона разработана слабо и нуждается в пересмотре. Все коконопряды Неотропики являются дендробионтами; равнинные виды широко распространены в тропических лесах Амазонии, но горные виды Nesara, Titya, Laruma, Norapidia, Pompeja, Tolitia и Prorifrons узко локальны.
Чилийско-Патагонская область. Фауна коконопрядов слабо изучена, известна только для Чили (Ureta, 1957), откуда указаны 2 рода Macromphalia и Euglyphus с 12 видами. Эндемичных родов нет; степень эндемичности видов не выяснена – считается, что они субэндемичны для хорона.
5.1.4. Нотогейское царство. Несмотря на небольшое число отмеченных здесь родов (17) и видов (94), фауна коконопрядов Нотогеи крайне специфична и полностью эндемична и на родовом и на видовом уровнях.
Австралийская область. Фауна коконопрядов Австралии насчитывает 71 вид из 12 родов из триб Pinarini и Argudini; все они эндемичны, не имеют прямых связей с родами Ориентальной подобласти, и лишь восточноавстралийские Digglesia и Cyclophragma обладают сродством к папуасским Pararguda и Pseudophyllodes. Эндемичных таксонов надродового уровня нет.
Новозеландская область. Коконопрядов нет (Dugdale, 1988).
Папуасская область. Крайне специализированная фауна коконопрядов хорона насчитывает 23 вида из 5 родов (Isostigena, Pseudophyllodes, Pararguda, Sporostigena и новый род для “Taragama” proserpina), относящихся к трибам Pinarini и Argudini; часть видов до сих пор еще не описаны. Все виды и роды являются эндемиками Новой Гвинеи, не обнаруживают родства с видами Индо-Малайской области, но близки к таковым Австралии, что позволяет рассматривать Папуасскую область внутри Нотогейского царства. Несмотря на высокий уровень родового и видового эндемизма, эндемичных таксонов надродового уровня нет.
5.2. Анализ неоднородности фауны коконопрядов. Для выявления неоднородности фауны коконопрядов Земного шара, территории всех континентов и острова были рассмотрены в рамках 149 относительно небольших участков, для которых оказалось возможным выявить достоверный набор обитающих там видов (Приложение 3). При типологическом анализе локальных фаун на уровне родов использован коэффициент сходства Жаккара-Наумова для количественных признаков. Анализ проведен с помощью метода факторной классификации (1976). В результате было получено 23 типа фаун (рис. 2).
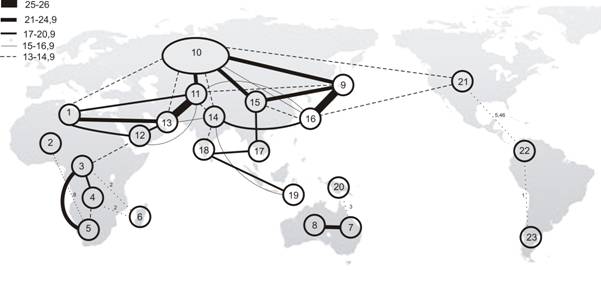
Рис. 2. Граф неоднородности фауны коконопрядов на уровне родов. Сходство фаун между регионами указано линиями пропорциональной толщины (в %%).
На родовом уровне довольно четко выражены 4 группы фаун: Евразиатская, Афротропическая, Новогвинейская и Американская, которые соответствуют крупным зоогеографическим хоронам. При этом фауны Северной Африки и аридных зон Сахеля обладают высоким уровнем сходства с Палеарктической фауной; отдельные фауны Евразии, имея высокий уровень сходства (19-21% и выше) по своему родовому составу, дают четкое разделение на Палеарктическую и Ориентальную области. Фауна Приморья (9), в отличие от Среднеамурской, оказалась в одном кластере с фауной Ориентальной области, и уровень связи по градиенту Китай–Приморье–Индокитай оказался высоким, более 19%. Среднеамурско-Восточноазиатско-Тибетская смешанная фауна (15) имеет высокий уровень сходства как с палеарктической Европейско-Кавказско-Переднеазиатско-Сибирской фауной (10), так и с Южнокитайско-Индийско-Зондской фауной (17) – соответственно 21% и 17%, и является переходной между фаунами Палеарктики и Ориентальной области. Гималайско-Гиндукушская горная фауна (14) обособлена от собственно Ориентальной, имея с ней сходство всего 11%. Крайне обособленными оказались и фауны Малых Зондских островов, что связано с обедненностью их видового состава.
Фауна Афротропической области образовала три тесно связанных между собой кластера (3, 4, 5). Сходство фаун пустынь Намибии и Сахели невелико из-за бедного видового состава. Уровень сходства континентальной фауны с фауной Мадагаскара (6) очень низкий – 2%, всего 2 общих рода. Через Аравию с ее сильно обедненным вариантом аридных фаун Эфиопии и Сомали прослеживается связь фауны Афротропики с фаунами Загроса (13) и Северной Африки (1), которая может отражать историческое расселение афротропических элементов с образованием изолятов для видов, вытесняемых на край ареалов в ходе плейстоценовой аридизации климата.
Отсутствие коконопрядов на Молуккских островах делает возможным провести границу между Ориентальной и Нотогейской фаунами по линии Лайдеккера. Фауна Новой Гвинеи имеет очень слабые связи (3%) с фауной Австралии, но не обнаруживает связей с Патагонией (23). Последняя, хоть и объединяется с фаунами Неарктики и Неотропики (21, 22), но при крайне низком уровне сходства (1%); вероятно, это является следствием ее слабой изученности.
Сходство между фаунами коконопрядов Палеарктики и Неарктики незначительно, причем последняя оказывается значительно ближе к палеарктической (13%), чем к центральноамериканской (5,5%). Предполагается, что в результате комплексной ревизии Неарктической фауны уровень сходства между ними существенно возрастет.
5.3. Эндемизм и его особенности у коконопрядов. Распределение коконопрядов жестко связано с границами материков, и виды-космополиты среди них не известны (табл. 1).
Низкая летная активность, а у некоторых видов (Chondrostega, Mesocelis, отдельные Lasiocampa и Tolype) и бескрылость самок накладывают ограничения на распространение видов, и коконопряды часто образуют узколокальные таксоны. Практически все высокие горные системы населены эндемичными
Табл. 1
Особенности видового и родового эндемизма
коконопрядов различных хоронов
Хорон | Родов | Видов | ||||
Всего | Эндемичных | %% | Всего | Эндемичных | %% | |
Голарктическое царство | 73 | 17 | 23,3 | 455 | 390 | 85,7 |
Бореальная область | 28 | 2 | 7,1 | 75 | 8 | 10,7 |
Область Древнего Средиземья | 26 | 9 | 34,6 | 112 | 92 | 82 |
Восточноазиатская область | 41 | 10 | 24,4 | 210 | 110 | 52,4 |
Сонорская область | 17 | 4 | 23,5 | 66 | 60 | 90,9 |
Палеотропическое царство | ||||||
Ориентальная область | 32 | 2 | 6,3 | 208 | 102 | 48 |
Афротропическая область | 86 | 80 | 93 | 600 | 600 | 100 |
Мадагаскарская область | 26 | 24 | 92,3 | 94 | 94 | 100 |
Неотропическое царство | 16 | 3 | 18,8 | 496 | 470 | 94,8 |
Неотропическая область | 16 | 0 | 0 | 450 | 398 | 88,9 |
Чилийско-Патагонская область | 2 | 0 | 0 | 13 | 11 | 84,6 |
Нотогейское царство | 17 | 17 | 100 | 94 | 94 | 100 |
Австралийская область | 12 | 12 | 100 | 71 | 71 | 100 |
Новозеландская область | 0 | 0 | 0 | 0 | 0 | 0 |
Папуасская область | 5 | 5 | 100 | 23 | 23 | 100 |
видами; к ним же привязаны и центры эндемизма семейства. В Евразии наибольшее число локальных эндемиков приходится на Гималайско-Тибетский регион (60 видов и 3 рода – Baodera, Similodora, Phachebia). Их число уменьшается по направлению к северу, и в горах Центрального и Восточного Китая эндемичными являются хвоегрызущие представители родов Cosmotriche, Dendrolimus, Kunugia, Paradoxopla, Takanea и Pachypasoides (20 видов). Два вида эндемичны для островов Японии, 1 – для Алтая. На Урале и в Альпах эндемичные виды не отмечены.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
