
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Структура глутаминовой тРНК из организма Escherichia coli
Автор:
Аннотация: были изучены вторичная структура глутаминовой тРНК (pdb код – 1exd) и ее внутримолекулярные контакты, поддерживающих третичную структуру.
Ключевые слова — глутаминовая тРНК, 1exd.
Введение
Рибонуклеиновые кислоты (РНК) представляют собой полимеры из нуклеофосфатных звеньев, соединенных фосфодиэфирной связью. В качестве азотистых оснований в РНК присутствуют урацил, цитозин, аденин и гуанин.
Транспортные РНК участвуют в процессе трансляции в качестве промежуточного связующего звена между нуклеиновыми кислотами и белками. Это небольшие молекулы из 70-90 нуклеотидов, которые с помощью своих антикодонов «узнают» за счет спаривания оснований определенные кодоны мРНК. На 3’-конце (CCA-конец) они несут тот активный остаток аминокислоты, который согласно генетическому коду соответствует очередному кодону мРНК [1].
Известно, что вторичная структура тРНК по своему внешнему виду похожа на «клеверный лист». Это изображение не передает особенностей строения молекулы в пространстве. Структура молекулы в пространстве называется L-формой. Заметим, что для молекул тРНК характерно присутствие большого числа разнообразных модифицированных нуклеозидов, часто называемых минорными [2].
Глутаминовая тРНК является одной из 45 тРНК кишечной палочки (Escherichia coli).
Цели работы
Структура глутаминовой тРНК была исследована биоинформатическими методами, на основании чего были сделаны выводы о вторичной и третичной структуре молекулы.
Материалы и методы
Модель пространственной структуры глутаминовой тРНК, полученная кристаллографическими методами, была получена из базы данных PDB (http://www. pdb. org). Структура молекулы изучалась с помощью программы RasMol. Для более подробного изучения комплементарных взаимодействий и особенностей глутаминовой тРНК использовались программы mfold, реализующая алгоритм Зукера, и find_pair пакета 3DNA.
Результаты
· С помощью программы find_pair был получен список комплементарных пар нуклеотидов в структуре глутаминовой тРНК.
Из Таблицы 1 Приложения видно, что данная тРНК содержит четыре участка, образующие спираль.
Из записи PDB файла можно сделать вывод, что нестандартных оснований молекула не содержит. По результатам программы find_pair видно, что в структуре существует 9 неканонических взаимодействий: 6 из них не соответствуют парам A-U, C-G, еще 3, возможно, характеризуются необычнымb свойствами связи (хотя, казалось бы, являются каноническими). Помимо этого из записи PDB файла получаем таблицу с не-уотсон-криковскими взаимодействиями (см. Таблицу 2 Приложения). Заметим, что пара 914 (A) – 921 (A), выделенная программой find_pair как неканоническая, не отмечена в списке не-уотсон-криковских взаимодействий списка PDB файла. Придумать объяснение этому факту мы не смогли.
На основании информации о спиральных участках была составлена Схема 1 Приложения, иллюстрирующая вторичную структуру глутаминовой тРНК.
· При работе с программой RasMol были выявлены следующие взаимодействия, поддерживающие стабильность пространственной структуры глутаминовой тРНК:
1. Водородные связи.
Некомплементарные взаимодействия: 914 (AA);
915 (GG);
918 (GU).
Комплементарные взаимодествия: 919 (GC);
954 (UA);
913 (AG).
2. Неспиральный стекинг:
976 (A), 975 (C), 973 (G) - взаимодействие внутри 3’ конца;
945 (G), 909 (C), 946 (G) – взаимодействие с дополнительной петлей;
957 (G), 919 (GC), 918 (GU) - взаимодействия между D - и T-петлями.
Схема 1 Приложения показывает приведенные выше взаимодействия.
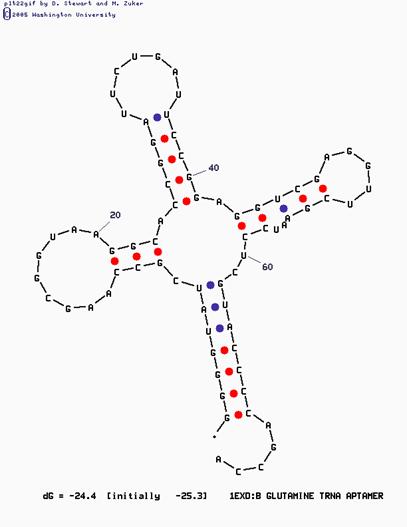
· С помощью программы mfold, по алгоритму Зукера было сделано предсказание о возможной вторичной структуре глутаминовой тРНК. Для получения всех возможных предсказаний варьировался параметр программы P=N (то значение свободной энергии, на которое энергия полученной структуры может превысить лучшее предсказание). Наиболее близкое изображение (Схема 2 Приложения) было получено уже при P=15.
Обсуждение
Пространственная структура тРНК поддерживается в основном некомплементарными водородными взаимодействиями, а также неспиральным стекингом между различными частями молекулы. За счет того, что D - и T-петли плотно взаимодействуют получается L-форма тРНК в пространстве.
Предсказание по алгоритму Зукера сильно отличается от пространственной структуры молекулы, восстановленной по кристаллографическим данным. Программа Зукера при построении предсказания по возможности минимизирует нестандартные взаимодействия, которые, как показано в исследовании, поддерживают L-форму. В нашем случае по алгоритму Зукера акцепторный стебель выходит на две пары нуклеотидов длиннее, а акцепторный стебель – на три пары нуклеотидов короче. D-петля предсказана верно, но строение Т-петли практически полностью не соответствует изображению, восстановленному по кристаллографическим данным. Структура, предсказанная по алгоритму Зукера, не может выполнять функций тРНК.
тРНК выполняет две основные функции: акцепторную - способность ковалентно связываться с аминоацильным остатком, превращаясь в аминоацил тРНК, и адаптерную - способность узнавать триплет генетического кода, соответствующий, транспортируемой аминокислоте (в нашем случае глутамин), и обеспечить поступление аминокислоты на нужное место в белковой цепи. Ключевыми для выполнения этих функций являются структуры антикодон (CUG для глутаминовой тРНК) и CCA-конец акцепторного стебля, которые не участвуют во взаимодействиях с другими частями молекулы.
Сопроводительные материалы
В файле 1exd. spt содержится скрипт для Rasmol, позволяющий визуализировать основные элементы структуры тРНК из PDB записи 1exd.
Приложение
Таблица 1. Список комплементарных взаимодействий в глутаминовой тРНК.
Желтым цветом выделены участки, образующие спираль. Синим шрифтом выделены неканонические пары.
1 | 902 | G | — | C | 971 |
2 | 903 | G | — | C | 970 |
3 | 904 | G | — | C | 969 |
4 | 905 | G | — | C | 968 |
5 | 906 | U | — | A | 967 |
6 | 907 | A | — | U | 966 |
7 | 949 | C | — | G | 965 |
8 | 950 | G | — | C | 964 |
9 | 951 | A | — | U | 963 |
10 | 952 | G | — | C | 962 |
11 | 953 | G | — | C | 961 |
12 | 954 | U | — | A | 958 |
13 | 955 | U | — | G | 918 |
14 | 937 | A | — | U | 933 |
15 | 938 | U | — | U | 932 |
16 | 939 | U | — | A | 931 |
17 | 940 | C | — | G | 930 |
18 | 941 | C | — | G | 929 |
19 | 942 | G | — | C | 928 |
20 | 943 | G | — | C | 927 |
21 | 944 | A | — | A | 926 |
22 | 910 | G | — | C | 925 |
23 | 911 | C | — | G | 924 |
24 | 912 | C | — | G | 923 |
25 | 913 | A | — | G | 946 |
26 | 914 | A | — | A | 921 |
27 | 915 | G | — | U | 948 |
28 | 919 | G | — | C | 956 |
Таблица 2. Таблица, отражающая не-уотсон-криковские взаимодействия, полученная из данных PDB записи глутаминовой тРНК. Синим цветом выделены пары, совпадающие с результатами счета программы find_pair.
903 | G | - | C | 970 |
904 | G | - | C | 969 |
908 | U | - | A | 914 |
910 | G | - | G | 945 |
913 | A | - | G | 946 |
915 | G | - | U | 948 |
918 | G | - | U | 955 |
919 | G | - | C | 956 |
926 | A | - | A | 944 |
928 | C | - | G | 942 |
930 | G | - | C | 940 |
932 | U | - | U | 938 |
933 | U | - | A | 937 |
954 | U | - | A | 958 |
Схема 1. Схема вторичной структуры глутаминовой тРНК, а также взаимодействий, поддерживающих ее третичную структуру.
Синим шрифтом выделены неканонические пары. Красным шрифтом обозначен антикодон. Желтой заливкой отмечены нуклеотиды, участвующие в неспиральном стекинге. Подчеркиванием выделены водородные связи, отвечающие за стабильность тРНК.
AGCCA
5’ G–C 3’
|
![]() G–C
G–C
G–C
 GAA U–A UA
GAA U–A UA
![]()
|
|
![]()
![]()
![]() G ||| ||||| G
G ||| ||||| G
 G GGC CGAGG C
G GGC CGAGG C
UAA U UU
G
![]() G
G
|
C–G
|
![]() G–C
G–C
G–C
A–U
U–U
U–A
|
![]() U
U
Схема 2. Изображение втоичной структуры глутаминовой тРНК, построенной по алогоритму Зукера.
Благодарности
Благодарю Ришу-тян за хорошее настроение и моральную поддержку, старшее поколение за то, что они уже прошли эту пытку и за то, что иногда отвечал прямо на заданные вопросы.
Литература
1. -Г., 2004. Наглядная биохимия. Издательство «МИР».
2., 1998. Строение транспортных РНК и их функция на первом (предрибосомальном) этапе биосинтеза белков. Соросовский образовательный журнал, №11, 71-77 (http://www. *****/nauka/Soros/pdf/9811_071.pdf).
