
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
Изменение пространственной организации хромосомы 6 в ядрах трофоцитов и клеток слюнных желез Calliphora erythrocephala Mg. (Diptera: Calliphoridae) в процессе политенизации
03.03.04 - клеточная биология, цитология, гистология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Томск 2012
Работа выполнена в лаборатории эволюционной цитогенетики Научно-исследовательского института биологии и биофизики Томского государственного университета, г. Томск.
Научный руководитель: доктор биологических наук,
Официальные оппоненты:
доктор биологических наук, заведующий лаборатории хромосомной инженерии, Федеральное государственное бюджетное учреждение науки Институт молекулярной и клеточной биологии Сибирского отделения Российской академии наук, г. Новосибирск
кандидат биологических наук, заведующий лаборатории механизмов клеточной дифференцировки, Федеральное государственное бюджетное учреждение науки Институт цитологии и генетики Сибирского отделения Российской академии наук, г. Новосибирск.
Ведущее учреждение: Федеральное государственное бюджетное учреждение науки Институт биологии развития им. Российской академии наук, г. Москва
Защита диссертации состоится «__»____________2012 г. на заседании диссертационного совета Д.003.011.01 по защите диссертаций на соискание ученой степени кандидата наук, на соискание ученой степени доктора наук в ИЦиГ СО РАН в конференц-зале института г. Новосибирск, пр. ак. Лаврентьева, д. 10. тел/, ; е-mail: *****@.
С диссертацией можно ознакомиться в библиотеке ИЦиГ СО РАН.
Автореферат разослан «__»____________2012 г.

Ученый секретарь
диссертационного совета,
доктор биологических наук
ВВЕДЕНИЕ
Актуальность работы
Пространственная организация хромосом в ядре является одной из ключевых проблем современной клеточной биологии. Утверждение, что хромосомы занимают в интерфазном ядре определенные хромосомные территории в настоящий момент не подвергается сомнению и экспериментально доказано для клеток большинства животных (Cremer, 2001), растений (Pecinka et al., 2004; Berr et al., 2006), а также одноклеточных эукариот, таких как почкующиеся и делящиеся дрожжи (Bystricky et al., 2005; Molnar, Kleckner, 2008). В основе трехмерной организации интерфазного ядра эукариот лежит дифференциальное позиционирование различных районов хромосом относительно друг друга и ядерной оболочки. Известно, что гены, хромосомные сегменты и геном как целое, трехмерно упорядочены в пространстве ядра, и их специфические ассоциации, очевидно, позволяют регулировать позиции определенных генных продуктов внутри ядра и их транспорт в цитоплазму. Жёсткая пространственная организация и функциональность ядра обеспечиваются наличием двух типов связей - межхромосомных взаимодействий (Куличков, Жимулев, 1976) и хромосомно-мембранных (Стегний, 1979).
Муха Calliphora erythrocephala является интересным объектом с точки зрения изучения пространственной организации хромосом в ядре. У C. erythrocephala политенные хромосомы обнаружены в ядрах клеток слюнных желез (Handa et al., 1981), клетках эпидермиса (Pearson, 1974), трофоцитах (Ribbert, Bier, 1969), трихогенных клетках (Ribbert, 1967), клетках мальпигиевых сосудов (Thomson, Gunson, 1970). Выявляются четкие тканеспецифичные особенности в степени политении и типе организации политенных ядер, что вероятно связано с разным функциональным значением этих тканей. В ядрах клеток слюнных желез C. erythrocephala на всех этапах политенизации выявляются политенные хромосомы. Синапсис хроматид у таких хромосом ослаблен, хромосомы рыхлые. Поперечный рисунок сохраняется только в области наиболее крупных дисков. Организация ядер трофоцитов существенно отличается от стандартных типов организации политенного ядра. В ядрах трофоцитов политенные хромосомы в ходе эндоредупликации претерпевают ряд морфологических изменений (Bier, 1957). На начальных этапах политенизации ядра трофоцитов организованы в первичную ретикулярную структуру, далее в ходе эндоредупликации происходит формирование политенных хромосом, затем, помпоновидных хромосом. Помпоновидные хромосомы декомпактизуются и происходит формирование ядра с ретикулярной структурой, но только с высоким уровнем политении.
|
Ранее было показано, что в ядрах трофоцитов C. erythrocephala взаиморасположение хромосом подчиняется определенным закономерностям. Центромерные районы хромосом трофоцитов не объединены в хромоцентр и рассредоточены в пространстве ядра. Хромосома 6 всегда связана тонким тяжом с хромосомой 2. Хромосома 4 располагается рядом с хромосомой 3 и 5, а 5 – с хромосомой 1. Таким образом, зачастую можно обнаружить следующее ассоциированное распределение хромосом: 6 – 2, 4 – 3, 5 – 1 (Стегний и др., 1999). Было показано, что хромосомы 3 и 6 сохраняют локальные хромосомные территории в объеме ядра на протяжении всего процесса политенизации ядер трофоцитов (Ананьина, 2005). Эти результаты по пространственной организации ядер трофоцитов С. erythrocephala были получены на давленых препаратах ядер на стадии с политенными хромосомами. Как известно, ядро трехмерно и поэтому наиболее полное представление о расположении хромосом может быть получено только при изучении трехмерного пространства ядра.
В работах Бойса (Boyes at al., 1975) было показано, что хромосома 6 C. erythrocephala содержит гены рРНК, транскрипция которых сопровождается образованием ядрышка. Однако, до сих пор не было получено данных относительно того, только ли хромосома 6 является ядрышкообразующей.
Ядрышко - эволюционно-консервативный и наиболее крупный структурный домен клеточного ядра, который принято называть «фабрикой рибосом» (Olson et al., 2002; Andersen et al., 2005). За последние годы получены убедительные доказательства того, что большое количество биологических процессов, таких как старение клеток, модификация РНК, контроль клеточного цикла и ответ на стрессирующие факторы регулируется ядрышком (Olson, 2002; Tschochner, 2003; Grummt, 2005; Boisvert, 2007; Mayer, 2005; Sirri, 2008; McKeown, Shaw, 2009). Эти наблюдения позволяют говорить о многофункциональности ядрышка и его основных белков. Кроме того, было показано, что в клетках человека ядрышко и ядерная оболочка являются своеобразным каркасом для организации ядра (Chubb et al. 2002). Не вызывает сомнения, что ядрышко играет важную роль в пространственной организации ядра. Однако, как происходит влияние ядрышка на архитектуру ядра все же остается не известно. Понимание законов организации хромосом в пространстве ядра и роли ядрышка в этом процессе существенно приблизит к разгадке принципов реализации генетического материала в ядре.
|
Изучение организации хромосомы 6 и ядрышка в пространстве ядер трофоцитов и клеток слюнных желез C. erythrocephala имеет большое значение для понимания особенностей организации и функционирования генома в ядрах соматических клеток и клеток генеративной системы, клеток с разной морфологической структурой политенных хромосом, выполняющих разные функции.
Цель работы
Целью исследования является анализ организации ядрышкообразующей хромосомы 6 и ядрышка в пространстве ядер трофоцитов и клеток слюнных желез С. erythrocephala с разным уровнем политенизации. Для достижения цели исследования были поставлены следующие задачи:
1. В пространстве ядер трофоцитов C. erythrocephala изучить организацию ядрышкообразующей хромосомы 6 и генов рРНК в ходе политенизации с помощью метода 3D флуоресцентной in situ гибридизации (3D FISH).
2. В пространстве ядер клеток слюнных желез C. erythrocephala изучить организацию ядрышкообразующей хромосомы 6 и генов рРНК в ходе политенизации с помощью метода 3D флуоресцентной in situ гибридизации (3D FISH).
3. Определить локализацию кластера генов рРНК в пространстве ядер трофоцитов и клеток слюнных желез C. erythrocephala.
4. Провести анализ пространственной организации ядрышка в ядрах трофоцитов и клеток слюнных желез C. erythrocephala с разным уровнем политенизации с помощью окрашивания недавленных препаратов ядер азотнокислым серебром.
Научная новизна
Установлено, что ядрышкоорганизующей хромосомой у C. erythrocephala является только хромосома 6, так как кластер генов рРНК локализован исключительно в пределах хромосомной территории хромосомы 6. Впервые показано, что в ходе политенизации ядер происходит изменение организации ядрышкоорганизующей хромосомы 6 как в ядрах соматических клеток, так и клеток генеративной системы C. erythrocephala (клетки слюнных желез и трофоциты). Показано, что распад ядрышка, занимающего центральное положение в ядре на начальных этапах политенизации на микроядрышки, распределенные в пространстве ядра в высокополитенных ядрах, коррелирует с изменением организации ядрышкоорганизующей хромосомы 6.
Научно-практическая значимость работы
Результаты проведенного исследования позволяют расширить представления о закономерностях пространственной организации хромосом в ядре эукариотических организмов. Результаты данной работы могут быть использованы в курсах лекций по клеточной биологии для студентов биологических факультетов.
|
Положения, выносимые на защиту
1. Ядрышкоорганизующей хромосомой у C. erythrocephala является хромосома 6;
2. Процесс политенизации ядер трофоцитов и клеток слюнных желез сопровождается перемещением хромосомы 6 и рибосомных генов на периферию ядра и распадом ядрышка на микроядрышки.
Апробация работы
Полученные результаты исследования были представлены: на международной научной студенческой конференции «Студент и научно-технический прогресс» (Новосибирск, 2009); на международной конференции «Хромосома-2009» (Новосибирск, 2009); на международной конференции молодых ученых "Биология: от молекулы до биосферы" (Харьков, Украина, 2009); на международной конференции по кариосистематике «KARYO V» (Новосибирск, 2010); на международной научной конференции «Молодежь и прогресс в биологии» (International scientific conference «Youth and progress of biology»), (Львов, Украина, 2011); По теме диссертации опубликовано 14 печатных работ, из них 5 статей в журналах перечня ВАК.
Вклад автора
Основные результаты работы были получены автором самостоятельно. Анализ ультраструктуры ядер трофоцитов C. erythrocephala проводился совместно с (НПО «Вирион», г. Томск).
Структура и объем диссертации
Диссертация состоит из введения, трех глав (обзор литературы, материалы и методы, результаты и обсуждение), выводов и списка литературы. Работа изложена на 118 страницах, содержит 16 рисунков. В списке литературы приведено 198 источников.
МАТЕРИАЛЫ И МЕТОДЫ
1.Объект исследования
Объектом настоящего исследования является муха Calliphora erythrocephala Mg. (Diptera: Calliphoridae). Для исследования использовали слюнные железы личинок разных возрастов и трофоциты яичников имаго от 2 до 5 дней после выхода из пупария. Материал был взят из лабораторной культуры.
2. Получение хромосомоспецифичной ДНК хромосомы 6
ДНК хромосомы 6 была получена ранее методом микродиссекции материала политенных хромосом (Рубцов и др., 1999). FISH ДНК хромосомы 6 с политенными хромосомами показал, что зонд является хромосомоспецифичным (Вассерлауф и др., 2003; Ананьина и др., 2005).
3. Приготовление ДНК-зонда хромосомы 6 С. erythrocephala
|
4. Амплификация кластера рибосомных генов С. erythrocephala
Рибосомная ДНК С. erythrocephala была получена с помощью ПЦР геномной ДНК в присутствии олигонуклеотидных праймеров (СибЭнзим-Нск): 18 Sai F (5'-CCTGAGAAACGGCTACCACATC-3’); 18 Sbi R (5’-GAGTCTCGTTCGTTATCGGA-3’). Праймеры фланкируют ген 18S рРНК (Whiting et al., 1997). Геномная ДНК была получена из яичников С. erythrocephala с помощью набора для выделения ДНК Invisorb Spin Tissue Mini Kit (Invitek) по протоколу, рекомендованному производителем. Условия ПЦР включали: денатурацию ДНК 95 °C - 3 минуты, затем 25 циклов: 95 °C – 1 минута, 55 °C – 45 секунд, 72 °C – 2 минуты. Терминальная элонгация 72 °C - 8 минут.
5. Получение ДНК-зонда рДНК С. erythrocephala
ДНК-зонд рДНК С. erythrocephala готовили в реакции ПЦР с использованием в качестве модифицированного нуклеотида биотин-11-дУТФ (Fermentas). Условия ПЦР были идентичны описанным в разделе 4.
6. Двуцветная 3D флуоресцентная in situ гибридизация (3D FISH) ДНК хромосомы 6 и генов рРНК с политенными хромосомами трофоцитов и клеток слюнных желез С. erythrocephala
За основу методики 3D FISH, которая использовалась в работе, был взят описанный в литературе протокол (http://www. /researchtools/ protocol. php? protid=5#reagents). В протокол были внесены модификации, позволяющие проводить 3D FISH на ядрах трофоцитов яичников и клеток слюнных желез С. erythrocephala.
7. Получение недавленных препаратов ядер трофоцитов и клеток слюнных желез С. erythrocephala, окрашенных азотнокислым серебром
За основу была взята стандартная методика окрашивания ядрышка азотнокислым серебром (Рубцов, 2006). Внесение модификаций позволило использовать ее для окрашивания недавленых ядер трофоцитов и клеток слюнных желез С. erythrocephala.
8. Приготовление препаратов для изучения ультраструктуры ядер трофоцитов С. erythrocephala
Ультратонкие срезы трофоцитов С. erythrocephala готовили по методике Б. Уикли (Уикли, 1975) с модификациями. Изучение ультраструктуры проводили методом трансмиссионной электронной микроскопии (Карупу, 1984).
9. Микроскопический анализ
|
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В данной работе проведен анализ расположения хромосомы 6 и рибосомных генов С. erythrocephala в пространстве ядер трофоцитов и клеток слюнных желез С. erythrocephala. Для определения организации в пространстве ядра кластера рибосомных генов использовалась ДНК специфичная к гену 18 S РНК. Также изучен процесс формирования ядрышка в процессе политенизации хромосом в ядрах трофоцитов и клеток слюнных желез С. erythrocephala.
1. Пространственная организация хромосомы 6 в ядрах трофоцитов C. erythrocephala
|

|

Рисунок 2 - Изменение пространственной организации хромосомы 6 и рибосомных генов, связанное с декомпактизацией помпоновидных хромосом и образованием ретикулярной структуры в ядрах трофоцитов C. erythrocephala. а1,б1 – давленые препараты ядер, окрашенные орсеином; а2,б2 - графическая реконструкция оптических срезов целого ядра; а3,б3 - графическая реконструкция оптических срезов сигналов ДНК-зондов хромосомы 6 и рибосомных генов в пространстве. Хроматин окрашен DAPI (синий), ДНК-зонд хромосомы 6 (красный), ДНК-зонд генов рРНК (зеленый). Масштабная линейка 5 мкм.
|
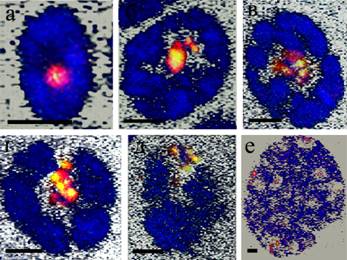
Рисунок 3 – Оптические срезы ядер трофоцитов C. erythrocephala на разных стадиях политенизации. а – стадия политенного ядра гермария, б – стадия ядра с первичной ретикулярной структурой, в – стадия ядра с политенными хромосомами, г – стадия ядра с помпоновидными хромосомами, д – стадия ядра с вторичной ретикулярной структурой. Масштабная линейка 5 мкм.
2. Ядрышки в ядрах трофоцитов C. erythrocephala на стадии с вторичной ретикулярной структурой
Для подтверждения того, что распределение хромосомы 6 в пространстве ядра на стадии с вторичной ретикулярной структурой связано с распределением микроядрышек был проведен анализ ядрышка в ядрах трофоцитов на стадии с вторичной ретикулярной структурой с помощью окрашивания азотнокислым серебром недавленых препаратов ядер трофоцитов C. erythrocephala (NOR-окрашивание). AgNO3-окрашивание выявляет белки ядрышка, связанные с транскрипционно активной частью ДНК ядрышкоорганизующих районов (Рубцов, 2006).
|
В ядрах трофоцитов C. erythrocephala на стадии с первичной ретикулярной структурой начинается формирование крупного ядрышка, ассоциированного с хромосомой 6 в центральной части ядра. Центральное положение и структура ядрышка сохраняются на стадиях, где хромосома 6 занимает центральное положение т. е. стадии политенных хромосом и стадии помпоновидных хромосом. При переходе ядра вновь к ретикулярной структуре, но уже с высоким уровнем политении процесс перемещение хромосомы 6 на периферию ядра сопровождается распадом крупного ядрышка на микроядрышки. Микроядрышки распределяются в пространстве ядра, сохраняя связь с ядрышкоорганизующими районами хромосомы 6.
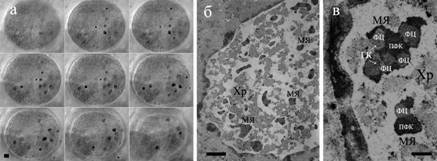
Рисунок 4 - Распределение микроядрышек в ядрах трофоцитов C. erythrocephala на стадии с вторичной ретикулярной структурой. а - серия оптических срезов ядра, окрашенного азотнокислым серебром. Микроядрышки (черный) на темно сером фоне (хроматин ядра); б, в - электронно-микроскопический анализ. Хр – хроматин, МЯ – микроядрышки, ФЦ – фибриллярный центр, ПФК – компактный фибриллярный компонент, ГК – гранулярный компартмент. Масштабная линейка 5 мкм в а и б, 1 мкм в в.
3. Пространственная организация ядер клеток слюнных желез C. erythrocephala
|
1. Хромосома 6 занимает компактное положение в центральной части ядер клеток слюнных желез C. erythrocephala.(Рисунок 5 а);
2. Хромосома 6 рассредоточена в пространстве ядер клеток слюнных желез C. erythrocephala (Рисунок 5 б).

Рисунок 5 – Варианты пространственной организации хромосомы 6 и рДНК в объеме ядер клеток слюнных желез C. erythrocephala. а1, б1 - оптические срезы ядер клеток слюнных желез; а2, б2 - графическая реконструкция оптических срезов половины ядра; а3, б3, - графическая реконструкция оптических срезов сигналов ДНК-зондов хромосомы 6 и рибосомных генов в пространстве. Хроматин окрашен DAPI (синий), ДНК-зонд хромосомы 6 (красный), ДНК-зонд генов рРНК (зеленый). Масштабная линейка 5 мкм.
В ходе эндоредупликации ядер клеток слюнных желез морфология ядер меняется не столь существенно как в ядрах трофоцитов и проявляется лишь в увеличении объема ядра. Исходя из этого, при анализе зависимости распределения материала хромосомы 6 в пространстве ядра от степени политенизации в качестве критерия оценки уровня политении нами использовался такой показатель как размер ядра. Было проанализировано 336 ядер клеток слюнных желез личинок C. erythrocephala взятых произвольно из разных районов слюнных желез разных особей. У всех ядер был измерен диаметр, который являлся критерием группировки данных. Каждое ядро оценивалось по характеру распределения хромосомы 6 и рДНК в объеме, т. е. было отнесено к одной из групп вариантов пространственной организации хромосомы 6.
|
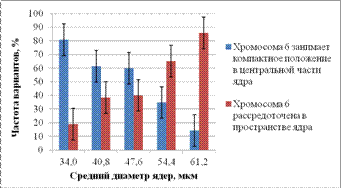
Рисунок 6 - Частоты ядер с разными вариантами распределения хромосомы 6 в пространстве ядер клеток слюнных желез C. erythrocephala.
Показано, что в ядрах клеток слюнных желез C. erythrocephala наряду с ядрами трофоцитов C. erythrocephala происходят изменения пространственной организации хромосомы 6 в ходе политенизации. Для ядер клеток слюнных желез C. erythrocephala на начальных этапах политенизации наиболее характерен вариант компактного расположения хромосомы 6 в центральной части ядра. С увеличением степени политении частота встречаемости варианта компактного расположения хромосомы 6 в пространстве ядра снижается. А вот частота встречаемости варианта распределения хромосомы 6 по всему объему ядра в ходе политенизации возрастает и на завершающих этапах политенизации ядер клеток слюнных желез C. erythrocephala значительно превосходит частоту встречаемости варианта компактного расположения хромосомы 6 в пространстве ядра.
4. Организация ядрышка в пространстве ядер клеток слюнных желез C. erythrocephala
|
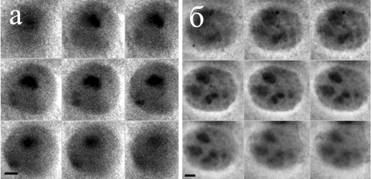
В ядрах клеток слюнных желез C. erythrocephala, как и в ядрах трофоцитов, в ходе политенизации происходит изменение организации ядрышка в объеме ядра. Однако процесс формирования ядрышка и его организации несколько отличается. Показано, что на начальных этапах политенизации, когда материал хромосомы 6 занимает компактную область в центральной части ядра происходит формирование ядрышка, ассоциированного с хромосомой 6 (Рисунок 7 а). Разделение хромосомы 6 на блоки и распределение их в пространстве ядра сопровождается разделением ядрышка на микроядрышки, расположенными ассоциировано с активно транскрибируемыми рибосомными генами на значительном расстоянии друг от друга в пространстве ядра (Рисунок 7 б).
Результаты исследования показали, что в ядрах клеток слюнных желез C. erythrocephala в ходе политенизации, несмотря на более стабильную структуру ядра, чем в ядрах трофоцитов так же происходит перераспределение хромосомы 6 в пространстве ядра. Подобная динамика хромосомы 6 вероятно связана с изменением общей экспрессионной активности ядра. Распределяясь по всему объему ядра материал хромосомы 6 активно функционирует. Вокруг активно транскрибируемых ядрышкоорганизующих районов формируются микроядрышки.
|
ЗАКЛЮЧЕНИЕ
В результате проведенного исследования показано, что в ядрах, относящихся к двум типам клеток с разным функциональным значением для организма (трофоциты и клетки слюнных желез C. erythrocephala) в ходе политенизации происходит изменение пространственной организации ядра. Было выявлено, что организация ядрышкообразующей хромосомы 6 и ассоциированного с ней ядрышка в объеме ядра существенно изменяется в ходе политенизации. На начальных этапах политенизации, как в ядрах трофоцитов, так и клеток слюнных желез хромосома 6 расположена в центральной части ядра. На этих стадиях происходит формирование крупного ядрышка, ассоциированного с хромосомой 6 и занимающего центральное положение в ядре. В ходе политенизации происходит изменение организации хромосомы 6 в пространстве ядра. В ядрах трофоцитов на стадии с вторичной ретикулярной структурой, как и высокополитенных ядрах клеток слюнных желез, ядрышкоорганизующие районы хромосомы 6 распределены в пространстве ядра и ассоциированы с микроядрышками. Кроме того, гены рРНК, расположенные в объеме высокополитенных ядер, являются активно экспрессирующимися, так как выявленные с помощью окрашивания азотнокислым серебром микроядрышки являются белковым компонентом ядрышка, связанным с транскрипционно активной рДНК.
Наличие схожих механизмов реорганизации ядер, которые происходят в клетках с совершенно разным функциональным значением, может объясняться особенностями функционирования рибосомных генов и ядрышка в высокополитенных ядрах. Вероятно, описанное явление распределения ядрышкоорганизующей хромосомы и ассоциированного с ним ядрышка в пространстве ядра является характерным для всех политенных ядер.
Однако, наряду с описанными закономерностями в изменении организации хромосомы 6 в ходе политенизации существуют и различия в пространственной организации хромосомы 6 в ядрах трофоцитов и клеток слюнных желез C. erythrocephala. В ядрах трофоцитов наблюдается перемещение хромосомы 6 на периферию ядра. В ядрах на стадии с вторичной ретикулярной структурой на периферии ядра расположена часть хромосомы 6, в составе которой находится так же и рибосомные гены. В ходе политенизации ядер клеток слюнных желез подобного перемещения хромосомы 6 на периферию ядра не происходит. В высокополитенных ядрах клеток слюнных желез хромосома 6 распределяется во всем пространстве ядра.
|
Были описаны различия в локализации рибосомных генов на блоках хромосомной территории хромосомы 6 в ядрах трофоцитов и клеток слюнных желез. На начальных этапах политенизации в ядрах обоих органов рибосомные гены располагаются в центральной части блоков хромосомной территории хромосомы 6. В ходе политенизации ядер трофоцитов происходит перемещение рибосомных генов на периферию блоков хромосомной территории. Подобное расположение сохраняется и на завершающих этапах политенизации. В ядрах клеток слюнных желез рибосомные гены на всем протяжении политенизации сохраняют центральное положение блоков хромосомной территории хромосомы 6.
Вероятно, в ядрах трофоцитов в ходе политенизации экспрессия генов рРНК существенно выше, чем в ядрах клеток слюнных желез. В результате активной и, все возрастающей транскрипции рибосомных генов, происходит синтез огромного количества белков ядрышка. Ядрышко увеличивается в размерах, что приводит к смещению ядрышкоорганизующих районов на периферию блоков хромосомной территории хромосомы 6. Однако, в высокополитенных ядрах экспрессия генов рРНК в ядрах трофоцитов ниже, чем в высокополитенных ядрах клеток слюнных желез, что связано с перемещением рДНК в ядрах трофоцитах на периферию ядра.
В целом существуют общие закономерности расположения хромосомных территорий внутри ядра. Расположение хромосомных территорий в активно транскрибируемых клетках динамично, что указывает на возможный механизм генной регуляции через внутриядерное позиционирование отдельных локусов хромосом. Кроме того, ядрышко, как значительный домен ядра, играет важную роль в пространственной организации ядра.
Описанная динамика хромосомной территории ядрышкоорганизующей хромосомы и ассоциированного с ней ядрышка в пространстве ядра, вероятно, связана с изменением общей экспрессионной активности ядра. Кроме того, вероятно, описанное распределение ядрышкоорганизующих районов и микроядрышек в пространстве высокополитенных ядер является особенностью функционирования ядрышка на завершающих этапах политенизации. Таким образом, изменения транскрипционной активности коррелируют с изменением пространственной организации ядра.
|
ВЫВОДЫ
1. В ядрах трофоцитов C. erythrocephala в ходе политенизации и морфологических преобразований ядра происходит изменение организации хромосомы 6 в пространстве ядра. На ранних этапах политенизации хромосома 6 занимает центральную часть ядра; на поздних этапах часть хромосомы 6 перемещается на периферию, а по всему ядру распределяются ядрышкоорганизующие районы хромосомы 6.
2. В ядрах клеток слюнных желез C. erythrocephala происходит изменение пространственной организации ядрышкоорганизующей хромосомы 6 в ходе политенизации. На ранних этапах политенизации хромосома 6 занимает компактную область в центральной части ядра. На поздних этапах политенизации происходит распределение хромосомы 6 на блоки в пространстве ядра.
3. Ядрышкоорганизующей у C. erythrocephala является только 6 хромосома, так как кластер генов рРНК локализован в пределах хромосомной территории хромосомы 6.
4. В ядрах трофоцитов и ядрах клеток слюнных желез C. erythrocephala распад крупного ядрышка, занимающего центральное положение в ядре, на микроядрышки, распределенные в пространстве ядра, коррелирует с изменением организации ядрышкоорганизующей хромосомы 6 в пространстве ядра.
|
СПИСОК ОСНОВНЫХ РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
1. Kokhanenko A. A., Anan’ina T. V., Stegniy V. N.The changes in chromosome 6 spatial organization during chromatin polytenization in the Calliphora erythrocephala Mg. (Diptera: Calliphoridae) nurse cells // Protoplasma. – 2012. - DOI 10.1007/s385-7.
2. Коханенко А.А., , Стегний динамика хромосомы 6 в трофоцитах Calliphora erythrocephala Mg. (Diptera: Calliphoridae) // ГенетикаТ. 46. - № 9. - С. .
3. , , , Стегний гомологии половых хромосом представителей семейства Calliphoridae // ГенетикаТ. 46, - № 9. – С. .
4. , Коханенко А.А., , Стегний строения яйцевых трубок яичников Calliphora erythrocephala (Mg.) (Diptera: Calliphoridae) // Вестник Томского государственного университета. – 2007. - № 000. – С. 175-180.
5. , , Стегний пространственного расположения хромосом питающих клеток Calliphora erythrocephala Mg. (Diptera: Calliphoridae) // В мире научных открытий№ 5(11). – С. 41-45.
