
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Лекция 4
СТРОЕНИЕ БЕЛКОВ
В строении белков различают четыре уровня организации молекулы: первичную, вторичную, третичную и четвертичную структуры. Первые два уровня характерны для всех белков. Третичная и четвертичная структуры имеют место только у глобулярных белков.
Первичная структура белков
Образование пептидной связи |

Первичная структура белков - это последовательность остатков аминокислот в полипептидной цепи. Порядок аминокислот в белке предопределен генетически последовательностью нуклеотидов в ДНК. Полипептид образуется путем взаимодействия карбоксильной группы одной аминокислоты с аминогруппой другой аминокислоты - пептидная связь.
"Голова" (NH2-) одной аминокислоты присоединяется к "хвосту" (-СООН) другой аминокислоты. Между аминокислотами замыкается пептидная связь (-CO-NH-), являющаяся единственным типом связи в первичной структуре белка. Как видно из приведенной схемы, при образовании пептидной связи высвобождается вода. Разрыв пептидной связи при гидролизе сопровождается присоединением воды по месту расщепляемой связи. Конечный продукт гидролиза белков и полипептидов - свободные аминокислоты.
Пептидная связь прочнее одинарных связей между углеродом и азотом, так как в результате таутомерии на 40% является двойной. По той же причине в полипептидной цепи вращение возможно только вокруг углеродных атомов, связанных с радикалом
Скелет у всех полипептидов одинаковый. Различаются полипептидные цепи характером и последовательностью радикалов. Называют полипептид по числу составляющих его остатков аминокислот: дипептидом, трипептидом и т. д.
Белками называют полипептиды, содержащие более 50 остатков аминокислот. Простейший белок - инсулин. Он содержит всего 51 остаток аминокислот. Рибонуклеаза включает 124 остатка, гемоглобин 574.
В белках последовательность аминокислот, т. е. первичная структура, строго определенная. Замена остатка одной аминокислоты на другую дает уже новый белок. Так, в инсулине быка в девятом положении находится остаток серина, а в инсулине барана - глицина. В инсулине человека и лошади различия касаются трех остатков аминокислот - восьмого, девятого и десятого. Все перечисленные инсулины имеют различную первичную структуру. Белки разных организмов с одинаковой функцией называются гомологичными.
Вторичная структура белков
Различают два основных типа вторичной структуры белков: спираль и складчатый слой.
| |
β-складчатый слой | α-спираль |
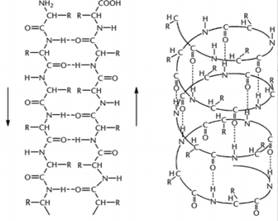
Спирали. Благодаря свободному вращению связей вокруг α-углеродного атома в полипептидной цепи, нарушается линейность полипептидной цепи. Это приводит к образованию спиралей. Существует 3 разновидности спиралей.
1. Для кератина характерна α-спираль. Полипептидная цепь кератина как бы накручена на воображаемый цилиндр. Витки друг возле друга удерживаются водородными связями между кислородом одной пептидной связи и водородом другой пептидной связи. Водородные связи в 20 раз слабее ковалентных связей между кислородом и водородом, но благодаря их многочисленности они довольно прочно, удерживают спираль.
2. β-спираль обнаружена в белке бактерий. Один виток р-спирали состоит из 22 остатков аминокислот, β-спираль - полая труба, α-спираль - заполненный цилиндр.
3. Ломаная спираль характерна для коллагена. Такая разновидность спирали является следствием высокого содержания в коллагене глицина и пролина с гидроксипролином - аминокислот, нарушающих "правильность" спирали.
Складчатый слой характерен для белка шелка - фиброина. Направление рядом лежащих цепей в складчатом слое противоположное (антипараллельны) Друг возле друга цепи удерживаются водородными связями.
Спирали и складчатые слои в фибриллярных белках часто дают сверх-вторичные структуры или суперспирали. Так, 7 α-спиралей кератина дают суперспираль. В свою очередь, 11 суперспиралей кератина образуют микрофибриллу волоса.
Вторичная структура глобулярных белков не столь однообразна как у фибриллярных белков. Так, в молекуле миоглобина спирализовано 77% полипептидной цепи и 23% не спирализовано. Степень спирализации инсулина - 60%, яичного альбумина - 40%, пепсина - 28%. Полипептидная цепь химотрипсина почти не содержит спирализованных участков, однако здесь есть складчатые, слои, петли, изгибы и т. д.
В структуре глобулярных белков с молекулярной массой свыше 20 тыс. Да различают понятие домен - небольшие участки в 100-150 остатков аминокислот с характерной структурой. Их называют структурными доменами.
Между доменами и отдельными структурными элементами внутри домена имеются так называемые шарнирные участки. Часто в одном белке обнаруживается несколько похожих однотипных доменов.
Существует еще понятие функциональный домен. В последнем случае один или несколько структурных доменов вместе образуют функционально обособленный участок в молекуле белка: субстратную площадку, окружение активного центра фермента или ингибитора, ионный канал в мембране и др.
Третичная структура - расположение полипептидной цепи (спирализованной, малоспирализованной или неспирализованной) в трехмерном пространстве.
Несмотря на кажущуюся беспорядочность глобулярного клубка, его строение строго определенное и имеет некоторые закономерности.
1. Полипептидные цепи в глобуле упакованы очень плотно.
2. Обычно полярные группы белка находятся на поверхности глобулы, а гидрофобные радикалы спрятаны внутри нее.
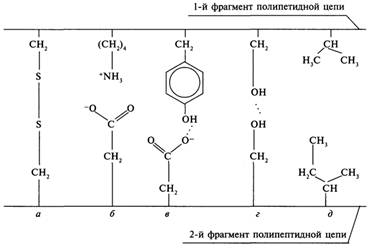 Третичная структура белков удерживается следующими силами (см. рисунок):
Третичная структура белков удерживается следующими силами (см. рисунок):
а) дисульфидными мостиками, образующимися между остатками цистеина, расположенных на удаленных участках полипептидной цепи;
б) электростатическим притяжением между противоположно заряженными ионизированными группами (ионными связями);
в, г) водородными связями между пептидными связями, между гидроксильными и карбоксильными группами и др.;
д) ван-дер-ваальсовыми силами (например, гидрофобным взаимодействием между радикалами гидрофобных (неполярных) аминокислот);
Все перечисленные силы слабые, но в комплексе способны удержать полипептидную цепь в уникальной конформации. Вместе с тем, благодаря непрочности этих связей, становятся возможными конформационные изменения в молекуле белка, необходимые для его функционирования. Непрочность позволяет белку изменять структуру в зависимости от условий окружающей среды.
Четвертичная структура определяется количеством полипептидных цепей в молекуле белка. Не все белки имеют этот уровень структуры, а только олигомерные, в которых 2 и более полипептидных цепи. Отдельные цепи со своей вторичной и третичной структурой называются протомерами олигомерного белка. Так, молекула гемоглобина состоит из 4 полипептидных цепей (протомеров). Четвертичная структура олигомерных белков удерживается теми же силами, что и третичная структура, с той лишь разницей, что они объединяют разные полипептидные цепи.
Свойства белков
Растворимость белков
Белки обладают различной растворимостью в воде (смотри классификацию белков). Будучи коллоидно-дисперсными системами, растворы белков менее устойчивы по сравнению с истинными растворами солей и других веществ. Молекулы белка в растворах склонны к агрегации и седиментации. Гидратная оболочка повышает устойчивость белков в растворах. Эта оболочка образуется вследствие гидратации отдельных гидрофильных групп белка: карбоксильной, аминной, гидроксильной, тиоловой и пептидной связи. У глобулярных белков эти группы находятся на поверхности глобулы. Гидратная оболочка препятствует объединению (агрегации) молекул белка и, следовательно, разъединяет их.
Другой фактор устойчивости белка - заряд. В растворе при определенных условиях все молекулы данного белка имеют одинаковый по знаку и количеству заряд. Благодаря этому молекулы электростатически отталкиваются друг от друга, не объединяются и не выпадают в осадок.
Лишение белка одного из факторов устойчивости часто приводит к выпадению белка из раствора в осадок. Так, при добавлении к раствору белка достаточного количества солей щелочных и щелочноземельных металлов, спирта или ацетона белок выпадает в осадок. Это осаждение называется высаливанием. Механизм высаливания заключается в том, что ионы солей и молекулы спирта и ацетона, имея мощную собственную гидратную оболочку, отнимают воду у молекулы белка. Разные белки высаливаются при разной концентрации солей. Глобулины высаливаются в полунасыщенном растворе сульфата аммония, а альбумины только в насыщенном растворе этой соли. Фракционное высаливание используется для разделения и очистки белков.
Некоторые белки выпадают в осадок при рН, соответствующем изоэлектрической точке. Так, казеин осаждается при рН 4,7, поскольку при этом значении рН молекулы не имеют заряда и быстро агрегируются в крупные, неустойчивые в растворе частицы. Другие белки более устойчивы, и для их осаждения необходимо воздействие на оба фактора устойчивости белков.
Диализ белков
Диализ. Черные кружки - молекулы белка, белые кружки - молекулы хлористого натрия |
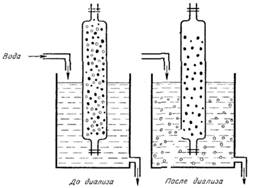
Благодаря крупным размерам, молекулы белка не проникают через некоторые пленки; целлофан, рыбий пузырь и др. Это свойство используется для очистки белков от низкомолекулярных примесей, т. е. для диализа.
В целлофановый мешок наливается раствор белка с примесью солей, мешок помещается в сосуд, через который протекает дистиллированная вода. Мелкие ионы солей и др. вещества проникают через целлофан в воду и удаляются, а раствор белка остается в мешке.
Заряд белка
В составе белка, как правило, сумма кислых, отрицательно заряженных аминокислот (глутаминовой, аспарагиновой) не равняется сумме основных, положительно заряженных аминокислот (лизина, аргинина, гистидина). В силу этого белки в воде имеют заряд либо положительный, либо отрицательный. При подкислении раствора такого белка, (в избытке Н+), будет подавляться ионизация карбоксильных групп и наступает такой момент, когда сумма положительно заряженных групп будет равна сумме отрицательно заряженных. В этом случае молекула белка в целом не имеет заряда. Такое состояние белка называется изоэлектрическим, а рН, при котором наступает изоэлектрическое состояние, называется изоэлектрической точкой (ИЭТ). ИЭТ - одна из важнейших характеристик белка.
При дальнейшем подкислении раствора белок становится положительно заряженным. Происходит перезарядка молекул белка. Если же взять положительно заряженный белок, то при подщелачивании он приобретает вначале изоэлектрическое состояние, а затем становится заряженным отрицательно.
Общее правило такое: белок при рН ниже ИЭТ заряжен положительно и является катионом, а при рН выше ИЭТ заряжен отрицательно и является анионом.
Различие в заряде белков позволяет разделить их в постоянном электрическом поле. Этот метод разделения называется электрофорезом.
В основе ионообменной хроматографии также лежит различие в заряде разделяемых веществ смеси.
Денатурация белков
Денатурацией называется любое негидролитическое изменение структуры белков, сопровождающееся изменением их биологической активности и функции. Денатурацию могут вызвать многие факторы: кипячение, высокая температура, ультрафиолетовое и ионизирующие излучение, избыточное давление, соли тяжелых металлов, экстремальные значения рН (крепкие кислоты и щелочи), некоторые органические соединения.
Нагревание и различного вида излучения разрушают в белке водородные и ионные связи. Сильные кислоты, щелочи и концентрированные растворы солей разрывают ионные связи. Тяжелые металлы дают прочные связи с карбоксианионами и разрывают ионые связи. Органические растворители и детергенты нарушают гидрофобные взаимодействия и разрывают водородные связи в белках.
При денатурации изменяются или разрушаются все слабые связи в белке: водородные, электростатические, гидрофобные и т. д., но остаются нетронутыми пептидные связи.
Признаками денатурации являются:
1) изменение растворимости. Растворенный в воде белок выпадает в осадок или, наоборот, нерастворимый белок переходит в раствор;
2) изменение оптической активности, например, угла вращения плоскости поляризованного луча;
3) появление новых реакционноспособных групп, спрятанных до денатурации внутри белковой глобулы;
4) главный и первый признак денатурации - потеря функции. Структурный белок становится рыхлым, ферменты теряют каталитическую активность и т. д.
После освобождения от денатурирующего агента белок постепенно приобретает свои первоначальные свойства. Этот процесс называется ренатурацией.
Оптические свойства белков
За исключением хромопротеинов белки не имеют окраски. Белки поглощают ультрафиолетовый свет с максимумом при λ=280 нм за счет ароматических аминокислот. Второй максимум поглощения при λ=216 нм принадлежит пептидной связи.
Растворы белков прозрачны, но обладают опалесценцией - при боковом освещении просматривается мутность. Перечисленные свойства используются для количественного определения белка.
МОНОНУКЛЕОТИДЫ
 Мононуклеотиды - соединения, построенные из пуринового или пиримидинового основания, рибозы или дезоксирибозы и остатка фосфорной кислоты. Фосфат в мононуклеотиде чаще всего присприсоединяется к пятому углеродному атому рибозы.
Мононуклеотиды - соединения, построенные из пуринового или пиримидинового основания, рибозы или дезоксирибозы и остатка фосфорной кислоты. Фосфат в мононуклеотиде чаще всего присприсоединяется к пятому углеродному атому рибозы.
Функции мононуклеотидов:
1) являются мономерами нуклеиновых кислот,
2) их производные принимают участие в энергетическом обмене, например АТФ,
3) некоторые моно - и динуклеотиды являются коферментами (ФАД, НАД, ФМН и др.)
Наиболее часто в нуклеотидах встречаются пять азотистых оснований: с пуриновым ядром: аденин, гуанин; с пиримидиновым ядром: урацил, цитозин, тимин.
| |
Пурин Гуанин Аденин | Пиримидин Цитозин Тимин Урацил |
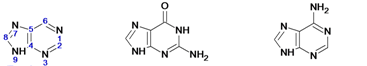
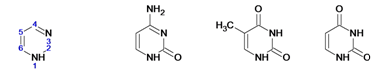
Кроме перечисленных оснований, встречаются метилированные, серосодержащие и др. производные азотистых оснований. Их называют минорными основаниями. Например, у прокариот встречаются: риботимидин, инозин, ксантин, гипоксантин и др. Всего известно около 60 азотистых оснований.
Азотистые основания и соединения, построенные из них, интенсивно поглощают свет в ультрафиолетовой области (260-280 нм). Это свойство используется для количественного определения веществ, содержащих в своем составе азотистые основания.
 Соединения, построенные из азотистого основания и рибозы или дезоксирибозы, называются нуклеозидами. В зависимости от строения пентозы различают рибонуклеозиды и дезоксирибонуклеозиды. Нуклеозиды легко расщепляются кислотами, но устойчивы к щелочам Нуклеозиды, к которым присоединены два или три остатка фосфорной кислоты называются нуклеозидди- и нуклеозидтрифосфатами.
Соединения, построенные из азотистого основания и рибозы или дезоксирибозы, называются нуклеозидами. В зависимости от строения пентозы различают рибонуклеозиды и дезоксирибонуклеозиды. Нуклеозиды легко расщепляются кислотами, но устойчивы к щелочам Нуклеозиды, к которым присоединены два или три остатка фосфорной кислоты называются нуклеозидди- и нуклеозидтрифосфатами.
Нуклеозидди - и нуклеозидтрифосфаты (АДФ, АТФ, ГТФ, УТФ и т. д.) содержат богатые энергией связи - макроэргические связи. Они обозначаются знаком ~ тильда. В отличие от обычной фосфатной связи, при гидролизе макроэргической связи высвобождается большое количество энергии, используемое клеткой.
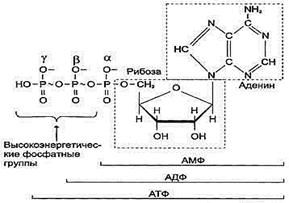 Важным производным нуклеозидов является цАМФ. Она образуется из АТФ при участии фермента аденилатциклазы. цАМФ участвует в регуляции метаболических процессов в клетке. В частности, она выполняет роль второго посредника в действии некоторых гормонов на клетку.
Важным производным нуклеозидов является цАМФ. Она образуется из АТФ при участии фермента аденилатциклазы. цАМФ участвует в регуляции метаболических процессов в клетке. В частности, она выполняет роль второго посредника в действии некоторых гормонов на клетку.
Соединения, построенные по типу нуклеотидов, входят в состав некоторых сложных ферментов, выполняя роль коферментов. Часто в составе таких коферментов обнаруживаются азотистые вещества, отличающиеся по строению от пуриновых и пиримидиновых оснований. Они не синтезируются в организме животных, а поступают с пищей (витамины).
Флавинмононуклеотид (ФМН) - фосфорилированный рибофлавин (витамин В2).
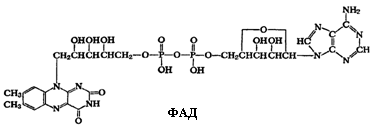
Флавинадениндинуклеотид (ФАД) состоит из двух нуклеотидов АМФ и ФМН.
|
|




 Никотинамидадениндинуклеотид (НАД) состоит из АМФ и второго нуклеотида, включающего в качестве азотистого основания никотинамид - витамин РР.
Никотинамидадениндинуклеотид (НАД) состоит из АМФ и второго нуклеотида, включающего в качестве азотистого основания никотинамид - витамин РР.
|
|
|

Никотинамидадениндинуклеотидфосфат (НАДФ) отличается от НАД тем, что АМФ этого динуклеотида фосфорилирована по гидроксилу третьего углерода рибозы.
Перечисленные выше соединения входят в состав дегидрогеназ - ферментов, осуществляющих транспорт электронов и протонов в окислительно - восстановительных реакциях.
 Кофермент А (коэнзим А, коА) состоит из фосфорилированной АМФ и фосфопантотеина (фосфорилированный витамин B3).
Кофермент А (коэнзим А, коА) состоит из фосфорилированной АМФ и фосфопантотеина (фосфорилированный витамин B3).
|

|
|

Коэнзим А осуществляет активацию и перенос ацильных радикалов, которые с помощью тиоэфирной связи присоединяются к SH-гpyппe.
В зависимости от переносимой кислоты соединения называют: ацетил-коэнзим А, малонил-коэнзим А, сукцинил-коэнзим А.
НУКЛЕИНОВЫЕ КИСЛОТЫ
Нуклеиновые кислоты - дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК) являются полимерами из дезоксирибонуклеотидов и рибонуклеотидов соответсвенно. Мононуклеотиды в нуклеиновых кислотах соединяются через остаток фосфорной кислоты между пятым углеродом рибозы и третьим углеродом рибозы соседнего нуклеотида.
Строение ДНК
В 1950 году Чаргафф обнаружил ряд закономерностей в нуклеотидном составе ДНК, которые впоследствии были названы правилами Чаргаффа. Вот эти правила: 1) Пур=Пир, 2) А=Т, 3) Г=Ц, 4) А+Ц=Г+Т. Правила"Чаргаффа помогли сформулировать комплементарную модель строения ДНК.
Первичная структура ДНК представлена полинуклеотидными цепями.
|
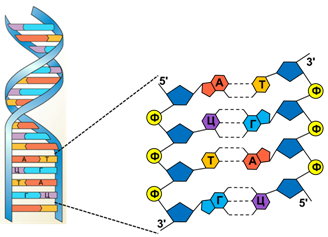 Структура ДНК
Структура ДНКЕсть ряд закономерностей в цепочках ДНК:
1) У вирусов и прокариот почти вся последовательность ДНК уникальна, у эукариот 30-40% ДНК составляют повторяющиеся последовательности, особенно много повторяющихся участков в ДНК в области центромеров.
2) Цепи ДНК не имеют разветвлений.
3) В ДНК много (тысячи) обратно бегущих последовательностей - палиндромов, "перевертышей". Примеры перевертышей в русском языке: "нажал кабан на баклажан". Палиндромы образуют крестообразные структуры - шпильки, играющие существенную роль в регуляции экспрессии (работе) генов.
Вторичная структура ДНК
В 1953 году Дж. Уотсон и Ф. Крик установили, что ДНК - это двойная спираль антипараллельных полинуклеотидных цепей. Удерживаются цепи друг возле друга водородными связями, образующимися между азотистыми основаниями, причем между аденином и тимином связь двойная, а между цитозином и гуанином - тройная. Снаружи двойной спирали ДНК расположен сахарофосфатный остов.
cпираль А cпираль В cпираль Z |
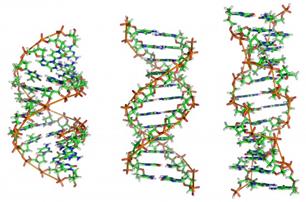
Комплементарно связанные азотистые основания обращены вовнутрь. В стопке азотистые основания смещены относительно друг друга. Существует несколько разновидностей спирали ДНК:
1) спираль типа В, обнаруживается при репликации ДНК;
2) спираль типа А, наблюдается при транскрипции;
3) спираль типа Z, закрученная влево, а не вправо как спираль А или В, бывает при кроссинговере.
4) Описаны еще спирали типа С и SBS. Последняя не закручена.
У вирусов может встречаться одноцепочечная ДНК.
Третичная структура ДНК
У прокариот молекулы ДНК кольцевые. У эукариот концы ДНК свободны - это линейная форма ДНК. У вирусов бывают линейные и кольцевые ДНК.
У прокариот нет ядра. У них ДНК вместе с белками прикрепляется к цитоплазматической мембране, образуя нуклеоид.
У эукариот ДНК отделена от остальной части клетки ядерной мембраной. В интерфазе ДНК эукариот сосредоточена в хроматиновых нитях. Хроматин, кроме ДНК содержит белки. 50% белка хроматина - гистоны. Гистоны содержат большое число остатков диаминокарбоновых кислот: аргинина и лизина. Это очень консервативные глобулярные белки практически одинаковые у всех эукариот. Вторая половина белков хроматина - негистоновые белки, характеризующиеся большим разнообразием.
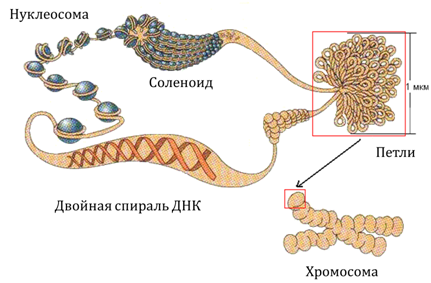 В хроматине рассматривается несколько уровней организации:
В хроматине рассматривается несколько уровней организации:
1) Нуклеосомы. На ядре из четырех пар молекул гистонов намотано почти два витка ДНК. Это - кор. Между корами расположен линкер - 40 пар нуклеотидов, частично покрытых гистоном и (или) негистоновыми белками или вообще не покрытых белками. Гистоны участвуют в активации и репрессии генов на уровне транскрипции.
2) Соленоиды: 6-10 нуклеосом дают один виток соленоида.
3) Петли. На скелете из негистоновых белков обнаруживаются петли в 30-90 тыс. пар нуклеотидов, у которых начало и конец располагаются рядом.
4) Высшим уровнем организации ДНК у эукариот является хромосома. Основу хромосомы сотавляет белковый матрикс, к которому крепится ДНК. На концах хромосомы находятся участки ДНК, называемые теломерами. С теломер может начинаться репликация; теломеры защищают концы хромосом от деградации.
При каждой репликации теломеры укорачиваются. Достигнув определенной критически малой величины теломер, клетка гибнет. Теломераза - фермент, восстанавливающий длину теломеров, делает клетку бессмертной. Теломераза есть в половых, стволовых и раковых клетках, в других клетках ее нет. В центре хромосомы находится центромера - тоже некодирующая ДНК, обеспечивающая правильное расхождение хромосом при делении клеток.
Большая часть ДНК находится в петлях. Здесь расположены гены. В каждой петле локализованы один или несколько генов. С матриксом хромосомы петли взаимодействуют некодирующими участками ДНК.
Физико-химические свойства ДНК
Хромосома - это одна молекула ДНК. У прокариот всего одна хромосома. Размеры ДНК различны от 5000 нуклеотидов у вирусов до 5 млрд (ее длина - 8см) у человека. Длина ДНК всех хромосом одной клетки человека около 2 м.
ДНК - белая волокнистая масса. Растворы очень вязкие. Вязкость увеличивается с увеличением молекулярной массы. Растворы ДНК поглощают ультрафиолетовый свет с максимумом при 260 нм. ДНК в воде заряжена отрицательно.
В кислой, щелочной средах при температуре° С в присутствии формамида, мочевины и ряда др. факторов происходит расхождение полинуклеотидных цепей ДНК - денатурация. При денатурации разрушаются водородные связи - ДНК "плавится". Температурой плавления считается такая, при которой ДНК денатурирует наполовину (разрушена половина водородных связей). При плавлении наблюдается повышение оптической плотности растворов при 260 нм - гиперхромный эффект.
Чем больше в ДНК Г-Ц пар, тем выше температура плавления, потому что Г-Ц пары прочнее, чем А-Т, поскольку удерживаются тремя водородными связями.
После снижения температуры денатурированная теплом ДНК восстанавливает свою вторичную структуру, происходит ренатурация, или отжиг, кислоты.
Если ДНК разных источников в смеси подвергнуть денатурации и отжигу, то произойдет гибридизация чужеродных цепей ДНК по законам комплементарности. Возможна гибридизация цепей ДНК и РНК. В этом случае образуется гибридная нуклеиновая кислота, в которой одна цепь - РНК, другая - ДНК.
