
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

 БИЛЕТ 19
БИЛЕТ 19
•Циклические нуклеотиды. Строение, свойства, механизмы синтеза и распада.
 Основания – пуриновые (аденин (А) гуанин (G). Важные интермедиаты - Гипоксантин (ГКс) и ксантин (Кс) Пиримидиновые цитозин (C), Тимин (Т) урацил (U). [Nu1] Есть также множество минорных. Свойства кето-енольная таутомерия – лактимная и лактамная формы существования. При pH=7 растворим G далее Кс. Неплохо – ураты моч. кислоты. Плохо – при ↓pH.
Основания – пуриновые (аденин (А) гуанин (G). Важные интермедиаты - Гипоксантин (ГКс) и ксантин (Кс) Пиримидиновые цитозин (C), Тимин (Т) урацил (U). [Nu1] Есть также множество минорных. Свойства кето-енольная таутомерия – лактимная и лактамная формы существования. При pH=7 растворим G далее Кс. Неплохо – ураты моч. кислоты. Плохо – при ↓pH.
Нуклеозиды – основание + рибоза или дезоксирибоза. [Nu2] Тимин – только дезоксирибоза! Нуклеотид – нуклеозид с фосф. гр. на ОН рибозы.
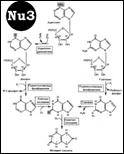
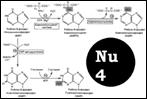 Синтез пуриновых. [Nu3] 1.образование ФРПФ (5-фосфо-рибозил-1-пирофосфата). ФРПФсинтетазой. 2. ФРПФ-амидотрансфераза превр-ет ФРПФ и Gln в фосфорибозиламин. 3.Вешается Gly. Глицинамид-киносинтетаза. 4. Формилирование N ферм-т – формилтрансфераза. 5. Амидирование как и вконденсация. 7 – карбоксилирование, СО2 в СООН превр-ся. 8. присоед аспартат.9 уходит Фумарат – остается новай NН2. Аденилосукциназа превр-ет 10. Формилтрансфераза – формилир-е. конденсация – и образование IMP. Важная роль фолатов! дают C8 и C2. Образование АМР, GMP схема [Nu4] Ингибиторы синтеза. Азасерин – 5.
Синтез пуриновых. [Nu3] 1.образование ФРПФ (5-фосфо-рибозил-1-пирофосфата). ФРПФсинтетазой. 2. ФРПФ-амидотрансфераза превр-ет ФРПФ и Gln в фосфорибозиламин. 3.Вешается Gly. Глицинамид-киносинтетаза. 4. Формилирование N ферм-т – формилтрансфераза. 5. Амидирование как и вконденсация. 7 – карбоксилирование, СО2 в СООН превр-ся. 8. присоед аспартат.9 уходит Фумарат – остается новай NН2. Аденилосукциназа превр-ет 10. Формилтрансфераза – формилир-е. конденсация – и образование IMP. Важная роль фолатов! дают C8 и C2. Образование АМР, GMP схема [Nu4] Ингибиторы синтеза. Азасерин – 5.
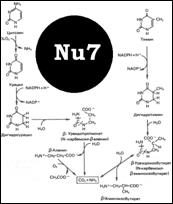
Диазонорлейцин – 2. 6-меркаптопурин – 13 и 14 синтеза AMP и GMP. Распад пуринов [Nu5] Синтез пиримидинов отличия от пуринового: рибозофосфат только на посл стадиях присоединяется. Сходство: необходимы Тетрагидрофолаты, аспартат, ФРПФ, глутамин и СО2. Распад пиримидинов схема [Nu7]
•Цикл мочевины. Связь орнитинового цикла с превращениями фумаровой и аспарагиновой
кислот. Нарушения синтеза и выведения мочевины. Гипераммониемии.
 Обновление белков – 1-2% в сутки. 80% высвободившихся АК Þсинтез новых белков. Сут. деградация белков 30-40г (т. е.5-7г азота) Þнужно 30-60 г белка (+!незаменимые АК). !Избыток АК не запасается (подверг. катаболизму). [Азотистый баланс].
Обновление белков – 1-2% в сутки. 80% высвободившихся АК Þсинтез новых белков. Сут. деградация белков 30-40г (т. е.5-7г азота) Þнужно 30-60 г белка (+!незаменимые АК). !Избыток АК не запасается (подверг. катаболизму). [Азотистый баланс].
Метаболизм АК: угл. скелет Þамфибол. интермедиаты; амино. группа Þпереаминирование/ окисл. дезамин. Þ1.5NH3 (рыбы; аммонотелические), 2.5Моч. к-та (птицы; урикотелические), 3.5Мочевина (млеки; уреотелические).
Катаболизм АК: I.Переаминирование. трансаминазы (аминотрансферазы). ВзаимоQ α-АК и α-кетокислоты. В акт. центре фермента – пиридоксальфосфат. 2 вида ферм.: 1.аланин-пируват - (обр-ся аланин) и 2.глутамат-α-кетоглутарат-трансаминазы (обр-ся глутамат). Аланин – субстрат еще и 2ого фермента Þазот всех АК может в кон. итоге переходить в L-глутамат (единст. АК, которая оч. быстро ~>окисл. дезаминирование). Субстратами для трансаминаз не явл-ся: Lys, Tre, Pro и гидрокси-Pro.
II.Окислительное дезаминирование. L-глутаматдегидрогеназы и оксидаза L- и D-аминокислот (роль???). Ферменты =окисляемые флавопротеины (FMN & FAD окисл-ся непоср. О2 ÞН2О2 Þрасщепление каталазой. Оксидазы АК Þα-иминок-та Þ+Н2О Þсоотв. α-кетокислота (потеря α-NH2-группы в виде иона аммония).
III.L-глутаматдегидрогеназа. Ингибиторы: ATP, GTP, NADH; стимуляторы: ADP. Гормон. регуляция. Косубстрат: NAD или NADP +р-ция обратима Þused и для синтеза, и для биосинтеза АК.
IV.Образование аммиака. В тканях +много бактериями в киш-ке из белка и мочевины Þv. porta Þhepar (!NH3 ~>в кровоток при циррозе Þколлатерали с v. cava inf. Þотравление ЦНС).
Отдельный трёп: 5NH3 в поч. канальцах (поддерж. КОС) из амидного-азота глутамина (а не из мочевины; не α-аминогруппа used!; глутманаза).
 V.Транспорт аммиака. Глутаматдегидрогеназа, обр-ие глутамина: глутаминсинтетаза (мх. фермент, тратим АТР). Обр. процесс – за счет глутаминазы. Аналогичный механизм – L-аспарагиназа.
V.Транспорт аммиака. Глутаматдегидрогеназа, обр-ие глутамина: глутаминсинтетаза (мх. фермент, тратим АТР). Обр. процесс – за счет глутаминазы. Аналогичный механизм – L-аспарагиназа.
Обмен аминокислот – гл. роль – у мышц (тут освобожд-ся 50% аминок-т, половина образующегося в мышцах азота – Ala и Gln) и печени (осн. поглощ. Ala) и кишечника (осн. поглощ. Gln). Почки – гл. источник серина, поглощ. Gln, Pro & Gly), Мозг – поглощ. Val (в мозге проще окисл-ся АК с разветвл. бок. цепью).
!Аланин – глюкогенная АК (Þглюконеогенез в печени).
БИОСИНТЕЗ МОЧЕВИНЫ
За сутки выделяем 16,5 г азота (85% ~>мочевина Þэкскреция).
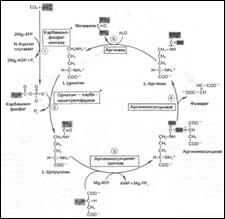
Цикл мочевины: 1.Синтез карбамоилфосфата (карбамоилфосфатсинтаза, в мх., тратим 2 АТР, нужен Mg2+ и дикарб. к-та); 2.Синтез цитруллина (L-орнитин-карбамоилтрансфераза, мх. печени, высокоспециф.); 3.Синтез аргининосукцината (аргининосукцинат-синтаза, тратим АТР); 4.Расщепление аргининосукцината на Arg и фумарат (аргининосукциназа; фумарат далее Þ*оксалоацетат, или аспартат при переаминировании); 5.Расщепление Arg на орнитин и мочевину (аргиназа).
НАРУШЕНИЯ ЦИКЛА МОЧЕВИНЫ
 •Гипераммониемия I типа (недостаток карбамоилфосфатсинтазы; наслед.); •Гипераммониемия II типа (недостаток орнитин-карбамоилтрансферазы, сцепл. с Х-хромосомой, ↑[Gln] в крови Ü↑синтеза глутамина); •Цитруллинемия (много цитрул. в моче, плазме, ликворе; дефект аргининосукцинат-синтазы); •Аргининосукцинатная ацидурия (наслед. рецесс. тип; ↑арг.-сукцината в крови, Qроста волос; недостаток аргининосукциназы).
•Гипераммониемия I типа (недостаток карбамоилфосфатсинтазы; наслед.); •Гипераммониемия II типа (недостаток орнитин-карбамоилтрансферазы, сцепл. с Х-хромосомой, ↑[Gln] в крови Ü↑синтеза глутамина); •Цитруллинемия (много цитрул. в моче, плазме, ликворе; дефект аргининосукцинат-синтазы); •Аргининосукцинатная ацидурия (наслед. рецесс. тип; ↑арг.-сукцината в крови, Qроста волос; недостаток аргининосукциназы).
•Рибосомный гиперцикл. Цикл элонгации трансляции.
Элонгацию на 3 стадии. В 1-ой пептидильный участок(P)рибосомы заним тРНК, несущая на 3'-конце растущую пептидную цепь. Затем вторая тРНК, соединенная с соответствующ ам. к-той Val-тPHKVal),взаимодействует своим антикодоном с кодоном мРНК, фиксированным на акцепторном участке(А, в данном случае GUG).тРНК связыв-ся в виде комплекса с ГТФ-содержащим белком, фактором элонгацииTu (EF-Tu). Диссоц-я комплекса происх только после того, как связан ГТФ гидролиз-ся до ГДФ и фосфата. До гидролиза ГТФ взаимод-е тРНК с мРНК относит-но слабое. Т обр, гидролиз ГТФ с участием комплекса служит лимитир-м фактором, дающим вр для проверки, правильно ли связана тРНК. Затем след белок, фактор элонгацииTs (EF-Ts),катализирует обмен ГДФ на ГТФ и т. обр регенерирует комплекс EF-Tu · GTP. Синтез пептидной св происх на след-ей стадии. Рибосомная «пептидилтрансфераза» катал-ет(без потребления АТФ)перенос растущ пептидной цепи от тРНК, наход-ся в Р-участке, на амгр валинового остатка, присоединенного к тРНКVаl, связанной на А-участке. Пептидилтрансферазная акт-ть рибосом завис не от какого-либо рибосомного белка, а,скорее всего, связана с 28S-PHK. Каталитически активныеРНК получ назв рибозимов. Предполагают, что существующие рибозимы мож рассм-ть как реликты«мира РНК»,раннего периода биохим эвол, когда белки еще не получ распростр-я и не приобрели такого знач. После переноса растущей цепи в А-участ, свобод аминоацил-тРНК диссоциирует от Р-участка и с рибосомой связыв-ся другойГТФ-содержащий фактор элонгации(EF-G · GTP).ГидролизГТФ этим фактором дает Е для транслокации рибосомы. Во вр этого проц рибосома сдвигает мРНК на 3 основ-я в направл 3'-конца. тРНК, несущая полипептид цепь, не мен полож-я относ-но мРНК, она попад в Р-участок рибосомы, в то вр как след кодон мРНК(в данном случае GUG),попадает в А-участок. Теперь рибосома готова для вступл-я в следующ цикл элонгации. Когда 1из стоп-кодонов(UAG, UAAилиUGA)попад в Α-участок, наступ терминация трансляции.
