


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Из всего количества солнечной энергии, поступающей в биосферу, примерно 70% преобразуется в тепло и затрачивается на испарение и другие процессы и только около 1% переходит непосредственно в пищу и другую биомассу. Не следует считать, что большая часть энергии теряется зря: она поддерживает определенные температурные условия на нашей планете, приводит в действие системы погоды и круговорот воды, необходимые для жизни. Энергия приливов и внутреннего тепла Земли в отдельных регионах может быть полезна, однако в тепловом балансе она невелика. Много тепловой энергии сосредоточено глубоко в недрах Земли, но она почти недоступна и потому практически не используется.
3.3. Пищевые цепи
Пищевая цепь - перенос энергии от ее источника (автотрофы или растения) через ряд организмов путем поедания одних другими. Пищевые цепи можно разделить на два основных вида.
Пастбищная пищевая цепь начинается с зеленого растения и тянется к пасущимся растительноядным животным и хищникам. В такой цепи при переносе большая часть (80-90%) потенциальной энергии теряется, переходя в тепло.
Детритная цепь – перенос энергии от мертвого органического вещества к микроорганизмам, а затем к детритофагам и к их хищникам.
В экосистемах пищевые цепи не изолированы одна от другой, а тесно переплетены, образуя пищевые сети. В сложных природных сообществах организмы, получающие энергию от Солнца через одинаковое число ступенек, относятся к одному и тому же трофическому уровню. Например, зеленые растения занимают первый трофический уровень (уровень продуцентов), травоядные – второй уровень (уровень первичных консументов), первичные хищники, поедающие травоядных – третий (уровень вторичных консументов), а вторичные хищники – четвертый (уровень третичных консументов).
Популяции одного и того же вида могут занимать один или несколько трофических уровней в зависимости от источников, которые они используют. Например, человек использует в пищу мясо животных, которые едят траву, улавливающую энергию Солнца. В этом случае – вторичный консумент на третьем трофическом уровне. Пищевая цепь может быть и более короткой, если человек питается исключительно растительной пищей. В случае смешанного питания (смесь растительной и животной пищи) он одновременно является первичным и вторичным консументом. При таком типе питания поток энергии разделяется между двумя или несколькими трофическими уровнями в пропорции, соответствующей долям растительной и животной пище в рационе.
При каждом переносе энергии в жизненной цепи значительная ее часть теряется, следовательно, число консументов, которые могут прожить при данном выходе первичной продукции, зависит от длины цепи.
3.4. Биологическая продуктивность экосистемы
Автотрофные экосистемы можно сравнить с промышленным предприятием, которое производит различные органические вещества. Используя солнечную энергию, диоксид углерода и элементы минерального питания, экосистемы производят биологическую продукцию – древесину, листовую массу растений, плоды, животную биомассу. Производительность экосистемы, измеряемая количеством органического вещества, которое создано за единицу времени на единицу площади, называется биологической продуктивностью. Единицы измерения продуктивности: г/кв. м. в день, кг/кв. м. в год, т/кв. км в год.
Различают первичную биологическую продукцию, которую создают растения в процессе фотосинтеза из диоксида углерода, воды и минеральных элементов, и вторичную биологическую продукцию, которую создают гетеротрофы (консументы и редуценты) в результате переработки растительной и животной биомассы. Первичную продукцию подразделяют на валовую – общее количество созданного органического вещества и чистую – то, что осталось после расходов на дыхание и корневые выделения. У большинства растений чистая продукция составляет примерно половину от валовой.
Поскольку консументы лишь используют ранее созданные органические вещества, вторичную продукцию на валовую и чистую не разделяют. Но ее количество также зависит от расходов на дыхание, которое тем больше, чем больше энергии затрачивает организм. При интенсивной физической нагрузке (например, у птиц во время миграции) продукция уменьшается. Привесы у сельскохозяйственных животных убывают, если им приходится затрачивать много энергии на переходы с отдаленных пастбищ или согревание тела в холодном помещении.
При переходе энергии с одного трофического уровня на другой (от растений к фитофагам, от фитофагов к хищникам первого порядка, от хищников первого порядка к хищникам второго порядка) с экскрементами и затратами на дыхание теряется примерно 90% энергии. Кроме того, фитофаги съедают только около 10% биомассы растений, остальная часть пополняет запас детрита и затем ее разрушают редуценты. Поэтому вторичная биологическая продукция в 20-50 раз меньше, чем первичная.
По продуктивности экосистемы разделяются на четыре класса.
1. Экосистемы очень высокой биологической продуктивности – свыше 2 кг/кв. м в год. К ним относятся заросли тростника в дельтах Волги, Дона и Урала. По продуктивности они близки к экосистемам тропических лесов и коралловых рифов.
2. Экосистемы высокой биологической продуктивности – 1-2 кг/кв. м в год. Это липово-дубовые леса, прибрежные заросли рогоза или тростника на озере, посевы кукурузы и многолетних трав при орошении и удобрении высокими дозами минеральных удобрений.
3. Экосистемы умеренной биологической продуктивности – 0,25-1 кг/кв. м в год. Такую продуктивность имеют многие посевы, сосновые и березовые леса, сенокосные луга и степи, заросшие водными растениями озера, “морские луга” из водорослей в Японском море.
4. Экосистемы низкой биологической продуктивности – менее 0,25 кг/кв. м в год. Это арктические пустыни островов Северного Ледовитого океана, тундры, полупустыни Прикаспия, вытоптанные скотом степные пастбища с низким и редким травостоем, горные степи, которые развиваются на почвах толщиной не более 5 см и состоят из растений-камнелюбов, покрывающих поверхность почвы на 20-40%. Такая же низкая продуктивность и у большей части морских экосистем.
Средняя продуктивность экосистем Земли не превышает 0,3 кг/кв. м в год, так как на планете преобладают низкопродуктивные экосистемы пустынь и океанов.
Биологическая продуктивность экосистемы отличается от запаса биомассы. Некоторые организмы в экосистеме живут много лет (деревья, крупные животные), и их биомасса переходит из года в год как некоторый капитал.
Биомасса леса велика за счет многолетних частей деревьев – стволов, веток, корней. Поэтому ежегодный прирост биологической продукции – новые листья, молодые веточки и корни, очередное годичное кольцо древесины и травяной покров – в 30-50 раз меньше, чем запас биомассы. На лугу запас биомассы значительно меньше, и он образован в основном корнями, живущими в почве по несколько лет, и корневищами растений. Он больше биологической продуктивности только в 3-5 раз. В полях биологическая продуктивность и запас биомассы практически равны, так как урожай наземных частей растений (и подземных, если это корнеплоды) убирают, а пожнивные остатки ржи или пшеницы запахивают в почву, где они к весне перегнивают. Как в луговой экосистеме, так и в экосистеме поля длительность жизни многочисленных почвенных беспозвоночных измеряется неделями и месяцами. Их биологическая продуктивность либо равна запасу биомассы, либо больше. Водоросли и мелкие беспозвоночные животные в водоемах живут по несколько дней или недель и потому за лето дают несколько поколений. В каждый конкретный момент биомасса организмов в озере или пруду меньше, чем их биологическая продукция за вегетационный сезон.
В некоторых водных экосистемах за счет того, что рыбы живут по несколько лет, а жизнь организмов фитопланктона непродолжительна, запас биомассы животных может быть выше запаса биомассы растений. Превышение биомассы животных над биомассой растений в морских экосистемах является правилом.
На суше 99,2% - биомасса растений, 0,8% - биомасса животных.
На море 6,3% - биомасса растений, 93,7% - биомасса животных.
Тема 4. БИОГЕОХИМИЧЕСКИЕ
ЦИКЛЫ
4.1 Структура и основные типы биогеохимических циклов
В экосистеме происходит постоянный круговорот элементов питания с участием биотического и абиотического компонентов. Движущей силов круговоротов служит солнечная энергия, которую используют непосредственно фотосинтезирующие организмы и затем передаю ее другим представителям биотического компонента. В итоге создается поток энергии и питательных веществ через экосистему, который носит название биогеохимического цикла.
Биогеохимические циклы – циркуляция в биосфере химических элементов и неорганических соединений по характерным путям из внешней среды в организмы, и из организмов во внешнюю среду. Такое перемещение элементов и неорганических соединений, необходимых для жизни, можно назвать круговоротом элементов питания.
При изучении круговорота удобно выделять две части, или два фактора:
· резервный фонд – большая масса медленно движущихся веществ, в основном небиологический компонент;
· подвижный, или обменный, фонд, для которого характерен быстрый обмен между органической и неорганической средой.
Для биосферы в целом можно выделить два основных типа биогеохимических циклов:
· круговорот газообразных веществ, с резервным фондом в атмосфере или гидросфере;
· осадочный цикл с резервным фондом в земной коре.
Такое разделение биогеохимических циклов основано на том, что некоторые круговороты, например те, в которых участвуют углерод, азот и кислород, из-за наличия крупных атмосферных или океанических фондов довольно быстро компенсируют нарушения. Так, накопленный в каком-либо месте избыток СО2 обычно быстро рассеивается воздушными потоками, а увеличение его концентрации в атмосфере способствует большему потреблению растениями и превращению в карбонаты в море. В целом круговороты газообразных веществ в глобальном масштабе можно считать хорошо забуференными, так как их способность к саморегуляции и поддержанию определенных концентраций различных веществ достаточно велика.
Следует отметить, что, хотя атмосфера и имеет большой резервный фонд и высокую способность к саморегуляции, все же они не беспредельны.
Осадочный цикл, в котором принимаю участие такие химические элементы, как фосфор и железо, в меньшей степени способен к саморегуляции и поэтому легче нарушается. Это связано с тем, что основная часть химических веществ сосредоточена в относительно малоподвижном и малоактивном резервном фонде земной коры. Если изъятие химических элементов в этих циклах происходит быстрее, чем возврат, какая-то их часть может на длительное время выбывать из круговорота. Механизмы возвращения химических элементов в круговорот основаны главным образом на биологических процессах.
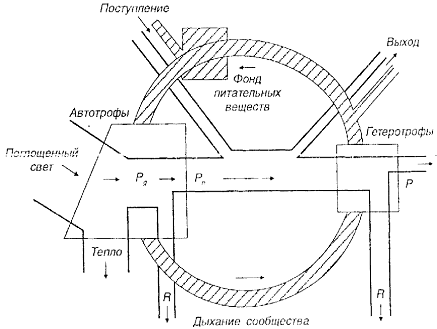
Рис. 2.3. Биогеохимический круговорот (заштрихованное кольцо) на фоне упрощенной схемы потока энергии (по Ю. Одуму, 1975): Pg – валовая продукция; Pn – чистая первичная продукция, которая может быть потреблена гетеротрофами в самой системе или же экспортирована; Р – вторичная продукция; R – дыхание
Схему биогеохимического цикла можно представить в сочетании с упрощенной схемой потока энергии, который приводит в движение круговорот веществ (рис. 2.3). В природе практически не наблюдается случаев, когда элементы равномерно распределены по всей экосистеме, к тому же они не всегда находятся в одной и той же форме. При изучении биогеохимических циклов изучают так называемый резервный фонд, то есть ту часть круговорота, которую условно можно считать отделенной физически или химически от организмов. Однако следует иметь в виду, что между доступными и недоступными фондами существует динамическое равновесие.
4.2 Биогенная миграция химических элементов и биогеохимические принципы
По Вернадскому, работа живого вещества в биосфере может проявляться в двух основных формах:
* химической (биохимической) – I род геологической деятельности;
* механической – II род такой деятельности.
Геологическая деятельность I рода – построение тела организмов и переваривание пищи, – конечно, является более значительной. Классическим стало функциональное определение жизни, данное Фридрихом Энгельсом: «жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь».
Сейчас появилась возможность вычислить скорость этого обмена. Так, по данным , в пшенице, например, полная смена атомов происходит для фосфора за 15 суток, а для кальция – в 10 раз быстрее: за 1,5 суток! Собственно говоря, постоянный обмен веществ между живым организмом и внешней средой и обуславливает проявление большинства функций живого вещества в биосфере. По подсчетам биолога -Кадошникова, в течение жизни человека через его тело проходит 75 т воды, 17 т углеродов, 2,5 т белков, 1,3 т жиров. Между тем по геохимическому эффекту своей физиологической деятельности человек отнюдь не самый важный вид разнородного живого вещества биосферы. Геохимический эффект физиологической деятельности организмов обратно пропорционален их размерам, и наиболее значимой оказывается деятельность прокариотов –бактерий и цианобактерий.
Большое значение имеет также количество пропускаемого через организм вещества. В этом отношении максимальный геохимический эффект на суше имеют грунтоеды, а в океане – илоеды и фильтраторы. Еще Чарлз Дарвин подсчитал, что слой экскрементов, выделяемых дождевыми червями на плодородных почвах Англии, составляет около 5 мм в год! Таким образом, почвенный пласт мощностью в 1 м дождевые черви полностью пропускают через свой кишечник за 200 лет. В океане с дождевыми червями по «пропускной способности» могут конкурировать их близкие родственники, представители того же типа кольчатых червей – полихеты, а также ракообразные. Достаточно 40 экземпляров полихет на 1 ![]() , чтобы поверхностный слой донных осадков мощностью в 20 – 30 см ежегодно проходил через их кишечник. Субстрат при этом существенно обогащается кальцием, железом, магнием, калием и фосфором по сравнению с исходными илами.
, чтобы поверхностный слой донных осадков мощностью в 20 – 30 см ежегодно проходил через их кишечник. Субстрат при этом существенно обогащается кальцием, железом, магнием, калием и фосфором по сравнению с исходными илами.
Копролиты (ископаемые остатки экспериментов) известны в геологических отложениях, начиная с ордовика, однако бесспорно, что большинство их при геологических описаниях не учитывается. Происходит это из – за слабой изученности вопроса и из – за отсутствия диагностических признаков для определения копролитов.
Между тем в донных отложениях современных водоемов фекальные комочки беспозвоночных распространены очень широко и нередко являются основной частью осадка. В южной Атлантике, например, илы почти нацело слагаются фекалиями планктонных ракообразных, а по берегам Северного моря донные осадки, образованные фекалиями мидий, имеют мощность до 8 м.
Биогенная миграция атомов II рода – механическая – отчетливо проявляется в наземных экосистемах с хорошо развитым почвенным покровом, позволяющим животным создавать глубокие укрытия (гнездовые камеры термитов, например, расположены на глубине 2 – 4 м от поверхности). Благодаря выбросам землероев, в верхние слои почвы попадают первичные невыветрившиеся минералы, которые, разлагаясь, вовлекаются в биологический круговорот. Недаром известный геолог (1889 – 1942) называл сурка-тарбагана «лучшим геологом Забайкалья» – его норы окружены «коллекциями» горных пород, добытых с глубины нескольких метров!
Понятие «нора» и «гнездо» обычно ассоциируются у нас с грызунами и птицами. Между тем биогенная миграция атомов II рода распространена не только в наземных, но и в морских экосистемах, и здесь ее роль, может быть, еще более значительна. И на дне моря организмы строят себе укрытия, причем не только в мягком, но и в скальном грунте. Олигохеты и полихеты углубляются в грунт на 40 см и более. Двустворчатые моллюски закрываются обычно неглубоко, но некоторые из них – солениды и миа – роют норы, которым позавидует и сурок: они достигают глубины нескольких метров. В зоне прибоя и на перемываемом волнами песке – вот беда! – норы не выроешь и гнездо не совьешь. Приходится сверлить скальные породы. И сверлят. Сверлят водоросли и губки, бактерии и моллюски, полихеты, морские ежи, рачки…
Сверлильщики появились в далеком геологическом прошлом. Источенные ими породы находят даже в докембрийских отложениях; и поныне они продолжают свою разрушительную работу. Сверлящая деятельность моллюсков фолад вызывает иногда катастрофические последствия. Когда в районе Сочи в результате непродуманного строительства берег обнажился от гальки, он начал отступать со скоростью до 4 м в год. Главным виновником разрушения были фолады, которые заселили каждый метр скального берега, сложенного глинистыми сланцами, и принялись дружно сверлить себе подводные норки. К счастью, был найден выход: берег стали укреплять поперечными стенками, а между ними засыпать гальку. В результате сверлильщики были уничтожены, движущаяся под ударами волн галька перемолола их. А в Западной Европе не менее опасную деятельность проводит случайно завезенный из Китая мохнаторукий краб – он проник во многие реки, и, строя свои норы, подрывает берега и разрушает плотины.
К биогенной миграции II рода можно отнести и перемещение самого живого вещества. Сюда относятся сезонные перелеты птиц, перемещения животных в поисках корма, массовые миграции животных. Естественно, что все эти разнообразные формы движения живого вызывают и транспортировку небиогенного вещества.
Вернадский, подразделял процессы, осуществляемые в биосфере живым веществом, по характеру самих процессов. Несколько иначе подошел к этому вопросу современник Владимира Ивановича . «Химическая деятельность организма вообще, имеющая геологическое значение, – писал Андрусов, – может быть сведена к двум категориям: во-первых, к образованию на нужной поверхности или внутри твердых выделений, способных сохраняться; во-вторых, к образованию жидких и газообразных выделений, способных вступать в различные химические реакции с окружающим неорганическим миром». По существу, эту же мысль развивала на современном материале микробиолог . Она указала, что миграция атомов химических элементов может быть как прямым, так и косвенным результатом жизнедеятельности организмов (в первую очередь бактерии). Для понимания той работы, которую совершает живое вещество в биосфере, очень важным являются три основных положения, которые Владимир Иванович называл «биогеохимическими принципами».
В формулировке они звучат следующим образом:
* I принцип: «Биогенная миграция атомов химических элементов в биосфере всегда стремится к максимальному своему проявлению».
* II принцип: «Эволюция видов в ходе геологического времени, приводящая к созданию форм жизни устойчивых в биосфере, идет в направлении, увеличивающем биогенную миграцию атомов биосфер» (или в другой формулировке: «При эволюции видов выживают те организмы, которые своею жизнью увеличивают биогенную геохимическую энергию»).
* III принцип: «В течение всего геологического времени, с криптозоя, заселение планеты должно было быть максимально возможное для всего живого вещества, которое тогда существовало».
Для Вернадского I биогеохимический принцип был тесно связан со способностью живого вещества неограниченно размножаться в оптимальных условиях. «Вихрь атомов», который представляет собой жизнь, по определению Жоржа Кювье, стремится к безграничной экспансии. Следствием этого и является максимальное проявление биогенной миграции атомов в биосфере.
II биогеохимический принцип, по существу, затрагивает кардинальную проблему современной биологической теории – вопрос о направленности эволюции организмов. По мысли Вернадского, преимущества в ходе эволюции получают те организмы, которые приобрели способность усваивать новые формы энергии или «научились» полнее использовать химическую энергию, запасенную в других организмах. В ходе биологической эволюции, таким образом, увеличивается «КПД» биосферы в целом.
В качестве примеров рассмотрим круговороты азота, фосфора и серы. Азот и фосфор часто является лимитирующими элементами и могут контролировать численность организмов, а сера – это химический элемент, который может служить примером связи между воздухом, водой и земной корой, то есть ее круговороту присущи особенности круговоротов азота и фосфора.
4.3 Круговороты азота, фосфора и серы
Круговорот азота. Это пример сложного и хорошо забуференного круговорота газообразных веществ. На рисунке 2.4 представлены два способа изображения сложного круговорота азота, каждый из которых иллюстрирует какую-то общую особенность или движущую силу. В круговороте азота ключевую роль играют микроорганизмы. Именно они осуществляют основные типы обмена между организмами и средой.
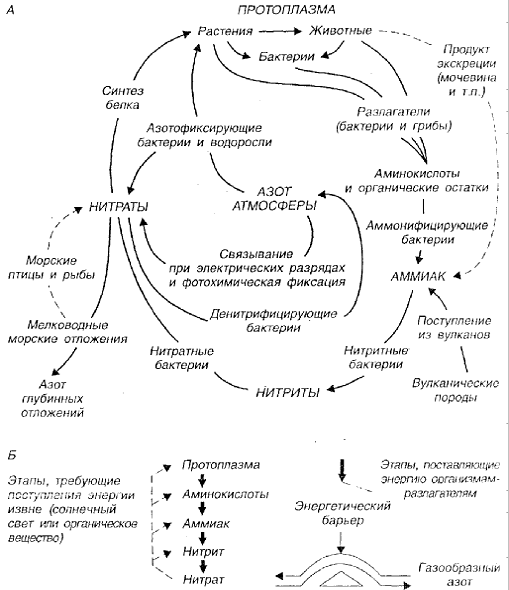
Рис 2.4. Два способа изображения биогеохимического круговорота азота (по Ю. Одуму, 1975): А – циркуляция азота между микроорганизмами и окружающей средой при участии микроорганизмов, отвечающих за ряд ключевых этапов; Б – те же основные этапы, но расположенные таким образом, что соединения, богатые энергией, находятся вверху; это позволяет отличить этапы, требующие затрат энергии, от процессов, протекающих с высвобождением энергии
На схеме А показано, что азот протоплазмы переводится из органической в неорганическую форму в результате деятельности бактерий-редуцентов, каждый вид которых выполняет определенную работу. Часть азота в конечном счете переводится в аммиачную и нитратную формы, доступные для питания растений. Как известно, воздух почти на 79 % состоит из азота и представляет собой одновременно крупнейший резервуар и буфер системы. Благодаря деятельности денитрифицирующих бактерий азот постоянно поступает атмосферу, а под действием азотофиксирующих бактерий возвращается в круговорот.
Схема Б иллюстрирует процессы, из которых складывается круговорот азота: фиксацию, ассимиляцию, нитрификацию, денитрификацию, разложение, выщелачивание, вынос, выпадение с осадками, и другие, а также оценки двух потоков, непосредственно связанных с деятельностью человека: выбросов в атмосферу и промышленной фиксации азота, соединения которого используются главным образом в качестве удобрений.
Так как содержание N2 в атмосфере резко не меняется, можно предположить, что приток т отток в целом уравновешивают друг друга.
На схеме Б представлены энергетические взаимоотношения в круговороте азота. Ступенчатый процесс разложения белков до нитратов служит источником энергии для организмов, принимающих участие в его разложении, а для обратного процесса требуются другие источники энергии – органическое вещество или солнечный свет. Например, хемосинтезирующие бактерии Nitrosomonas, превращающие аммиак в нитрит, получают энергию за счет разложения, а денитрифицирующие и азотофиксирующие – используют другие источники.
Однако ни животные, ни человек, ни растения потреблять молекулярный азот не могут. Огромное количество молекулярного азота в атмосфере в чрезвычайно малой степени затрагиваются биологическим круговоротом: общее отношение связанного азота к его количеству в природе составляет 1: 1000000 (Акимова, Хаскин, 1998). Несмотря на громадное количество молекулярного азота в атмосфере, он является одним из наиболее лимитирующих биогенных элементов. Столб воздуха над одним гектаром земной поверхности составляет 80000 т молекулярного азота. Если бы растения могли усваивать молекулярный азот, то такого его количества хватило бы для получения урожая 30 ц/га на полмиллиона лет. Однако растения могут использовать только азот минеральных соединений. Поэтому, буквально «купаясь» в молекулярном азоте, они испытывают его нехватку.
Из растений фиксировать азот могут только представители семейства бобовых, на корнях которых образуются клубеньки, состоящие из азотофиксирующих бактерий. Однако и среди бобовых далеко не все виды могут фиксировать атмосферный азот. Всего семейство бобовых насчитывает 13000 видов, а наличие клубеньковых бактерий обнаружено у 1300. Считается, что бактерии переводят в связанную форму приблизительно 1 млрд. т азота в год, промышленная его фиксация составляет около 90 млн. т.
Фиксировать азот могут следующие роды организмов:
· свободно живущие бактерии – Azotobakter и Clostridium (анаэроб);
· сибиотические клубеньковые бактерии бобовых растений – Rhizobium;
· цианобактерии – Anabaena, Nostoc и др.
Из всего азота, который ежегодно усваивается глобальным биотическим сообществом, около 80 % возвращается в круговорот суши и воды и только 20 % поступает из атмосферы с дождем и в результате фиксации.
Благодаря механизмам обратной связи, обеспечивающим саморегуляцию, круговорот азота можно считать относительно замкнутым, если рассматривать его в масштабе крупных площадей или всей биосферы.
В современных условиях человек своей деятельностью оказывает значительное влияние на круговорот азота: увеличивает содержание азота в резервном фонде (сжигание ископаемого топлива, осушение заболоченных земель, обработка почвы и т. д.) и снижает его содержание (выращиванием бобовых культур на громадных территориях, техническое связывание азота) в атмосфере.
Круговорот фосфора. В отличие от азота резервным фондом этого элемента служат горные породы и другие отложения, образовавиеся в прошлые геологические эпохи. По структуре круговорот фосфора проще, чем круговорот азота. Он циркулирует, постепенно переходя из органических соединений в фосфаты, которые снова могут использоваться растениями. Горные породы подвергаются воздействию выветривания, в результате чего фосфор высвобождается и становится доступным для растений. Под действием эрозионных процессов он попадает в море и на значительный промежуток времени высвобождается из круговорота. По всей вероятности, механизмы возврата фосфора в круговорот недостаточно эффективны и не возмещают его потерь. Перенос фосфора с морской воды на сушу не компенсирует его поток в море.
Деятельность человека ведет к усиленной потере фосфора, что делает круговорот недостаточно замкнутым. Важность фосфора как элемента, обеспечивающего продуктивность биосферы, со временем будет возрастать, так как уже сейчас он причисляется к редким макроэлементам. Поэтому возврат фосфора в круговорот имеет важное значение для человечества.
Круговорот серы. Круговорот серы имеет ряд характерных особенностей:
· обширный резервный фонд в почвах и меньший – в атмосфере;
· ключевая роль в быстро обменивающемся фонде микроорганизмов, выполняющих определенную работу в окислении или восстановлении;
· микробная регенерация из глубоководных отложений, в результате которой вверх движется газовая фаза (H2S);
· взаимодействие геохимических и метеорологических процессов с биологическими процессами;
· взаимодействие воздуха, воды и почвы в регуляции круговорота в глобальном масштабе.
Основная доступная форма серы – SO42– – восстанавливается автотрофами и включается в белки. Для растений серы требуется меньше, чем азота и фосфора, поэтому лимитирующим фактором она бывает реже. Тем не менее круговорот серы – ключевой в общем процессе продуцирования и разложения биомассы.
Круговороты различных элементов могут оказывать взаимное влияние друг на друга. Например, при образовании в осадках сульфидов железа фосфоров из нерастворимых соединений переходит в растворимые.
В последнее время на круговороты азота и серы все большее влияние оказывает промышленное загрязнение атмосферы. Особенно токсичны соединения азота в форме оксидов NO2 и N2O и серы – в форме SO2 которые являются промежуточными продуктами круговоротов этих элементов. В большинстве местообитаний их концентрация невелика, но в связи с неумеренным сжиганием топлива содержание в воздухе этих соединений, особенно в крупных промышленных центрах, увеличилось до такой степени, что они представляют опасность для важных биотических компонентов экосистемы.
Основным источником соединений азота являются выхлопные газы и другие промышленные выбросы, сернистого газа – продукты сжигания угля.
Особенно большой вред наносит SO2 растениям. Реагируя с водяным паром, он образует слабую серную кислоту, которая выпадает с осадками, известными как «кислотные дожди». Попав на листовую поверхность, H2SO4 вызывает химические ожоги, что снижает фотосинтезирующую поверхность растений.
Оксиды азота раздражают дыхательные пути высших животных и человека. Также следует иметь в виду, что, реагируя с другими соединениями, они могут образовывать соединения с синергическим эффектом, когда взаимодействие продуктов реакции больше суммарного воздействия каждого из реагирующих веществ в отдельности. Например, под действием ультрафиолетового излучения солнца NO2 вступает в реакцию с продуктами неполного сгорания углеводородов. В результате возникает фотохимический смог.
В конечном счете, оксиды азота и серы, попадающие в атмосферу, ухудшают качество жизни.
4.4 Глобальные круговороты углерода и воды
В глобальном масштабе биохимические круговороты воды и углекислого газа имеют, на наш взгляд, самое важное значение для человечества. Для биохимических круговоротов характерно наличие в атмосфере небольших, но подвижных фондов.
Атмосферный фонд СО2 в круговороте, по сравнению с запасами углерода в океанах, ископаемом топливе и других резервуарах земной коры, относительно невелик.
С наступлением научно-технического прогресса сбалансированные прежде потоки углерода между атмосферой, материками и океанами начинают поступать в атмосферу в количестве, которое не полностью может связаться растениями.
Существуют разные оценки влияния деятельности человека на обогащение атмосферы СО2 однако все авторы сходятся во мнении, что основными накопителями углерода являются леса, так как в биомассе лесов содержится в 1,5 раза, а в гумусе, содержащемся в почве – в 4 раза больше СО2, чем в атмосфере.
Растения – хороший регулятор содержания СО2 в атмосфере. Для большинства растений характерно увеличение интенсивности фотосинтеза при повышенном содержании диоксида углерода в воздухе.
Фотосинтезирующий «зеленый пояс» Земли и карбонатная система моря поддерживают постоянный уровень СО2 в атмосфере. Однако стремительное увеличение потребления горючих ископаемых, а также уменьшение поглотительной способности «зеленого пояса» приводят к тому, что содержание СО2 в атмосфере постепенно растет. Предполагают, что если уровень СО2 в атмосфере будет превышен вдвое (до начала активного влияния человека на окружающую среду он составлял 0,29 %), то не исключено повышение глобальной температуры на 1,5 – 4,5оС. Это может привести к таянию ледников и как следствие – к повышению уровня Мирового океана, а также к неблагоприятным последствиям в сельском хозяйстве. В настоящее время в США существует национальная научно-исследовательская программа по ведению сельского хозяйства на случай потепления или похолодания климата.
Помимо СО2 в атмосфере в небольших количествах присутствуют оксид углерода СО – 0,1 части на миллион и метан СН4 – 1,6 части на миллион. Эти углеродные соединения активно включены в круговорот и поэтому имеют небольшое время пребывание в атмосфере: СО – около 0,1 года, СН4 – 3,6 года, а СО2 – 4 года. Оксид углерода и метан образуются при неполном или аэробном разложении органического вещества и в атмосфере окисляются до СО2.
Накопление СО в глобальном масштабе не представляется реальным, но в городах, где воздух застаивается, имеет место повышение концентрации этого соединения, что негативно влияет на здоровье людей.
Метан образуется при разложении органического вещества в болотистых местностях и мелководных морях. По мнению некоторых ученых, метан выполняет полезную функцию – он поддерживает стабильность озонового слоя, который предохраняет все живое на Земле от гибельного воздействия ультрафиолетового излучения.
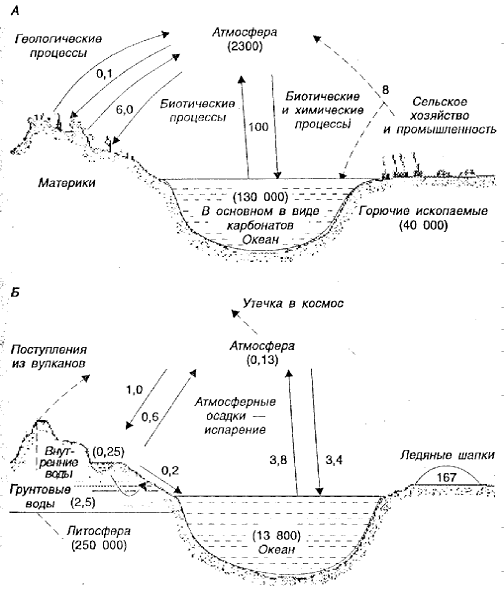
Рис. 2.5. Глобальные круговороты: А – круговорот диоксида углерода. Числа обозначают содержание СО2 в миллиардах тонн; Б – круговорот воды. Содержание Н2О указано в геограммах (1020 г) (по Ю. Одуму, 1975)
Фонд воды в атмосфере, как показано на рисунке 2.5 невелик, и скорость ее оборота выше, а время пребывания меньше, чем СО2. Как и на круговорот СО2, деятельность человека оказывает влияние на круговорот воды.
С энергетической точки зрения можно выделить две части круговорота СО2: «верхнюю», которая приводится в движение Солнцем, и «нижнюю», в которой выделяется энергия. Как уже отмечалось, около 30 % всей энергии Солнца, поступающей на поверхность Земли, затрачивается на приведение в движение круговорота воды.
В экологическом плане особое внимание следует обратить на два аспекта круговорота воды. Во-первых, море за счет испарения теряет больше воды, чем получает с осадками, то есть значительная часть осадков, поддерживающих экосистемы суши, в том числе и агроэкосистемы, состоит из воды, которая испарилась с поверхности моря. Во-вторых, в результате деятельности человека возрастает поверхностный сток и сокращается пополнение фонда грунтовых вод. Уже сейчас имеются территории, на которых используются грунтовые воды, накопившиеся в предыдущем столетии. Следовательно, в этом случае вода – невозобновимый ресурс. После истощения грунтовых вод ее будут доставлять с других территорий, что потребует вложения дополнительного количества энергии.
4.5 Осадочный цикл
Большинство элементов и соединений входит в общий осадочный цикл, циркуляция в котором осуществляется за счет эрозионных процессов, осадкообразования, горообразования, вулканической деятельности и биологического переноса.
Твердые частицы, переносимые по воздуху в виде пыли, могут выпадать на землю в виде сухих осадков или вместе с дождем. Они могут состоять из природных веществ (образующихся при вулканических извержениях, ветровой эрозии, лесных пожарах), а также из соединений антропогенного происхождения (например, ядовитые вещества, радиоактивные осадки), которые даже в небольших количествах могут оказывать мощное негативное влияние на живые организмы.
В осадочном цикле большое значение имеет перенос почвенных частиц и химических элементов под воздействием эрозионных процессов. В периоды с малой геологической активностью происходит перенос химических элементов с возвышенностей в понижения, моря и океаны (рис. 2.6).
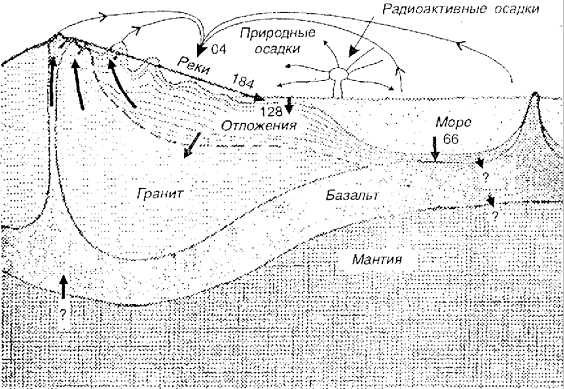
Рис. 2.6. Схема осадочного цикла (по Ю. Одуму, 1975)
Интенсивность осадочного цикла в разных регионах мира неодинакова и зависит от природно-климатических условий, освоенности территории, хозяйственной деятельности человека (табл. 2.3).
Таблица 2.3
Годовой вынос осадочного материала в океаны
(по Ю. Одуму, 1986)
Территория | Площадь водосбора, 1´106 км2 | Вынос общий | |
т/км2 | 1´109 т | ||
Северная Америка Африка Австралия Европа Азия | 20,7 19,4 19,9 5,2 9,3 26,9 | 634,0 414,3 181,3 297,5 233,0 346,2 | 1,96 1,20 0,54 0,23 0,32 15,91 |
Всего | 101,4 | å | 20,16 |
Из данных таблицы видно, что особенно велики потери плодородной почвы в Азии. Известно немало случаев, когда из-за разрушения пахотных земель вследствие развития эрозионных процессов люди покидали насиженные места или даже погибали от голода.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
