

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Если мы говорим, что конкретный человек умеет решать дифференциальные уравнения, это вовсе не значит, что его мозг способен выполнять дифференцирование. Это значит, что человек помнит последовательность действий над формулами, включая многократное обращение к собственной памяти и к справочникам, в результате которых будет получена формула-ответ. А дальше – трудоёмкий путь вычислений по формуле на основе заученных в школе и институте правил.
При всём разнообразии интеллектуальных задач, автору и его друзьям не удалось найти ни одной, которую нельзя было бы свести к задачам распознавания или к определению функций корреляции!
Нужно оговориться, что пока не вполне понятны и требуют дальнейшего изучения приёмы сверхсложных устных расчётов, быстро выполняемых отдельными людьми. Например, описаны такие факты:
– в 1980 г. Шакунтала Деви из Индии в присутствии комиссии за 28 секунд устно перемножила два 13-значных числа:
7
и 2 ,
получив в результате:
18
– в 1989 г. колумбиец Хайме Гарсиа Серрано за 15 секунд извлёк корень 13-й степени из 100-значного числа;
– в 1991 г. Александр Васильевич Некрасов из Липецка за 61 секунду в присутствии комиссии устно извлёк корень тысячной степени из 547-значного числа! (Все данные – из одесской газеты „Пресс-курьер” за 24–30 сентября 1998 г., стр. 6, со ссылкой на газету „Киевские ведомости”.)
Такие вычисления, конечно, опираются на отличную память. С учётом времени вычислений и чисел волн нервного возбуждения, способных пройти за это время в разных нейрокорреляторах мозга, видно, что расчёты дробятся на малые доли, не превышающие допустимую вычислительную сложность. Как раз отыскание алгоритма разделения вычислений на части, а также способность чётко, без отдыха и сбоев пройти все операции, составляют главную тайну подобных расчётов.
Мозг как вычислительное устройство, оказался ориентированным на решение задач высшей интеллектуальной сложности, словно Природа заранее отбросила более простые (например – арифметические) задачи „на плечи” вычислительных машин. Вероятно, окружающий мир выдвигает чаще всего именно задачи высшей сложности.
3.4.4. Мысль – организованная группа волн
Представление о прохождении волны-образа через нейрокоррелятор близко к известному выражению „мелькнула мысль”. Но элементарная осознаваемая мысль не эквивалентна прохождению единственной волны-образа. Элементарный акт мышления включает в себя обособленную, но внутренне функционально связанную группу волн нервного возбуждения, в которой каждая волна выполняет особую, индивидуальную функцию.
Данные исследований позволяют заключить, что мысль, как правило, включает в себя от трёх до десяти волн нервного возбуждения, причём запоминается и потому осознаётся только исходная информация и общий результат этого составного процесса.
Например, в одной из серий экспериментов [Sheatz, Chaрman, 1969] регистрировались волны электроэнцефалограмм при исследовании отношения человека к различным стимулам. Вызванные потенциалы состояли как минимум из трёх последовательных волн. Если испытуемый наперёд знал, что сигнал не имеет значения и не требует ответа, то амплитуды второй и третьей волн снижались. Если он должен был сначала правильно определить сигнал, а затем мог не отвечать и больше не обращать на него внимания – тогда уменьшалась только третья волна. И все три волны достигали максимума, когда испытуемому приходилось сперва на мгновенье задуматься над стимулом, а затем ответить на него.
В подобных экспериментах особенно проявилась независимость управления каждой солитонной волной, что принципиально отличает их от синусоидальных колебаний.
Различие в функциях разных волн цикла, вероятно, вытекает из различия задач, решаемых каждой из них. Во-первых, пришедшая информация должна быть зафиксирована в голографической памяти, что требует прихода одновременно с волной-образом и немодулированной опорной волны. Во-вторых, новый образ должен быть классифицирован, должен пройти сравнение с несколькими из хранящихся в памяти эталонов, для чего может понадобиться несколько волн нервного возбуждения. В-третьих, после отыскания наилучшей корреляции результат должен быть подан на выход, и зафиксирован в последующих отделах памяти, где регистрируются уже не образы, получаемые извне, а итог их корреляционного сравнения с ранее записанными, известными образами.
В промежутке между прохождением двух солитонов, в нервной ткани, не относящейся к области памяти, происходит „стирание” предыдущей информации, чтобы она не повлияла на независимое протекание дальнейших вычислений. Обычно такое „стирание” имеет вид перехода ткани к хаотической импульсной активности. По-видимому, характер этих процессов сходен с характером процессов в зрительном анализаторе. Вот как описывается цикличность работы нашего зрения в связи с саккадическими „скачками” глаза.
„В первый момент после скачка... система имеет характеристику фильтра низких пространственных частот,.. способна различать только крупные детали изображения... В следующие моменты разрешающая способность увеличивается... Процесс идёт непрерывно 50-70 мс и к концу этого времени... имеет... максимальную разрешающую способность. В этом оптимальном состоянии система находится 30-50 мс... /Затем/ ... происходит... функциональная дезорганизация полей. Благодаря этому стирается „предыстория” ... и они становятся готовыми к новому циклу восприятия...” [Подвигин, 1979]
* * *
С позиций концепции нейроголографии, мозг (имеются в виду, совместно, головной и спинной мозг) можно условно поделить на две части – на подсистему принятия решений и исполнительную подсистему.
Подсистема принятия решений – это та часть нервной системы человека или обладающего мозгом животного, которая обеспечивает выработку стратегии поведения в сложных, быстро изменяющихся условиях внешней среды. Исполнительная подсистема обеспечивает реализацию найденной стратегии с учётом выработанных ранее навыков. Принципы их построения сходны, но не одинаковы. В рамках данной работы, ради ограничения объёма, будем говорить только о первой из них.
Подсистема принятия решений представляет собой сложную иерархическую структуру, построенную из нейрокорреляторов, и сходную с пирамидой. Разнообразная сенсорная информация приходит в эту подсистему со стороны основания пирамиды, и по мере обработки перемещается вверх, пока не выльется у вершины в единственное принятое решение.
Корреляторы первого уровня принимают информацию непосредственно от разных сенсоров. Следующие уровни получают от предыдущих всё более сжатую и абстрагированную информацию. Если первый уровень обрабатывает информацию разнотипных сенсоров раздельно и определяет корреляционные сигналы первого порядка, то на высоких уровнях сигналы разнотипных сенсоров всё более объединяются, смешиваются, теряют второстепенные детали, уменьшаются в объёме. Происходит переход к вычислениям корреляционных функций всё более высоких порядков. Соответственно, на верхних уровнях уменьшается количество корреляторов.
От этажа к этажу пирамиды несколько меняется и содержание обрабатываемой информации. Сигналы на выходах корреляторов первого уровня говорят о степени и характере отличий вновь полученной информации от ранее зафиксированной. Корреляторы более высокого уровня определяют степень и характер новизны обнаруженных отличий по сравнению с хранящимися в памяти случаями. И так далее – по мере продвижения вычислений на верхние уровни пирамиды корреляторов происходит переход от обработки живых образов внешнего мира к обработке всё более абстрактной информации.
Принятие решения – это отыскание в памяти сходной ситуации, а затем для неё – удачной стратегии поведения. Чем привычнее задача, чем больший автоматизм её решения выработан предшествующим опытом, тем быстрее – на более низком уровне иерархической структуры – принимается решение (замыкается петля условного рефлекса). При более трудной ситуации процесс решения задачи докатывается до вершины пирамиды, максимально используя весь хранящийся в памяти опыт.
Сходное строение – в виде пирамиды нейрокорреляторов, но перевёрнутой вершиной вниз и опирающейся на вершину пирамиды принятия решений – имеет и исполнительная подсистема мозга. Здесь принятое решение, по мере поднятия на верхние этажи нейрокорреляторов, используя информацию, хранящуюся в памяти в виде привычных поз, движений, рабочих приёмов, всё более расширяется, превращается в массив сигналов, управляющих всеми элементами исполнительных органов.
Наиболее частые операции в работе мозга – это операции с данными из памяти: сравнение образов, поиск корреляционных связей и различий между ними, классификация образов по тем или иным признакам, выявление далёких ассоциаций и пр. Даже во время восприятия очень актуальной новой информации от органов чувств, не меньший объём данных вызывается из памяти.
Чем выше уровень развития животного, тем больше оно думает, т. е. тем большая доля вычислительной мощности мозга занята обработкой информации, извлекаемой из памяти, а не получаемой в этот момент от сенсоров.
3.4.5. Проблема инвариантности зрения
Поскольку зрительная система приносит подавляющую часть сенсорной информации, а также в связи с запросами робототехники, новые нейрокибернетические разработки обычно примериваются, в первую очередь, к зрительному анализатору.
Прямое приложение принципов КНГ к зрительному анализатору человека обнаруживает два резких несоответствия между свойствами голографических корреляторов и свойствами нашего зрения. Во-первых, корреляторы инвариантны (безразличны) к плоскопараллельному смещению анализируемого объекта в поле зрения, а человек к этому вовсе не безразличен, он упорно сосредотачивает взгляд на рассматриваемом предмете. Во-вторых, человек без малейшего напряжения узнаёт приблизившийся объект, если узнал его издали, тогда как для коррелятора, изображённого на рис. 3.4, это невозможно. Коррелятор работает только при том масштабе изображения, какой заложен в эталоне.
Анализ показал, что объяснение этих противоречий заключено в особой анатомии глаза и зрительного нерва. Система нейронов зрительного анализатора человека построена так, что наблюдаемое изображение деформируется путём сжатия его периферийных областей. Сжатие, насколько можно судить, соответствует логарифму расстояния от центра поля зрения. Деформирование изображения происходит под совместным действием трёх факторов:
– от центра к периферии сетчатки глаза уменьшается плотность расположения фоторецепторов;
– от центра к периферии сетчатки растёт число рецепторов, посылающих сигналы в одно и то же волокно зрительного нерва;
– на входе зрительного нерва в ядро, называемое наружным коленчатым телом таламуса, наблюдается редкое явление – плотность расположения нервных окончаний оказывается наиболее высокой на периферии пучка волокон и плавно уменьшается к центру.
Реализуемая таким способом логарифмическая деформация входного изображения превращает первый голографический коррелятор зрительной системы в одну из его разновидностей, в так называемый коррелятор Меллина [Василенко, Цыбулькин, 1985]. Отличительной особенностью коррелятора Меллина является инвариантность к размерам анализируемого образа. Иначе говоря, коррелятор Меллина, как и наше зрение, способен распознавать знакомые объекты при различной угловой величине, т. е. на разном расстоянии, лишь бы они были хорошо различимы. Это достигается нелинейностью (логарифмическим масштабом) системы относительно центра поля зрения, что влечёт за собой необходимость центрирования распознаваемого образа.
Следовательно, свойства нашей зрительной системы, как распознающего устройства, точно соответствуют специфике того типа голографического коррелятора, к которому она должна быть отнесена на основании её анатомических характеристик. Кстати, это лишний раз подтверждает голографическую природу вычислений.
3.4.6. О мышлении в целом
В главе 3.4.1. приведены данные, говорящие о существовании в мозге, по крайней мере, одного нейрокоррелятора. Но что говорит о существовании пирамидальной иерархической структуры из многих корреляторов? Подтверждение этому дало интересное общее свойство мозга, обнаруженное при исследовании условных рефлексов. Эксперименты выявили некую универсальную закономерность, понять природу которой исследователям не удалось.
Закономерность, о которой идёт речь, обнаружилась при изучении вариаций биопотенциалов мозга подопытного животного в ответ на изменения воздействий внешней среды. После того, как рефлекс выработан, кривая активации биопотенциалов в ответ на любое изменение внешних условий приобретала сходство с кривой частотного резонанса, как показано на рис. 3.7.
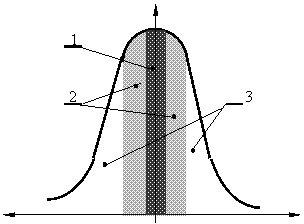 |
Рис. 3.7. Зависимость активации биопотенциалов мозга от изменения параметров сигнала при выработанном условном рефлексе [Кратин и др., 1982].
1 – область специфических условнорефлекторных реакций; 2 – область неспецифических вегетативных эффекторных ответов; 3 – область электроэнцефалографического проявления активации биопотенциалов мозга без эффекторных ответов.
Однако, вопреки стараниям исследователей, самое настойчивое экспериментальное изучение всех возможных „кандидатов” не выявило ни в мозге, ни в организме в целом, колебательного процесса, на который можно было бы возложить ответственность за такой „резонанс” [Кратин и др., 1982; Кратин, 1986].
Другая удивительная особенность явления состояла в том, что достижение пика электрической активности мозга зависело не от одной какой-то характеристики, одного раздражителя, а от целостного комплекса параметров, от всей суммы условий, существовавших во время исследований. Достаточно изменить любой параметр эксперимента или обстановки, окружавшей животное в ходе выработки условного рефлекса, чтобы изменился уровень активации биопотенциала.
Другими словами, мозг животного реагирует на мир в целом, биопотенциалы изменяются так, будто в мозгу „резонирует” единый полисенсорный образ внешнего мира!
То, что осталось загадкой для исследователей, получило чёткое объяснение с позиций КНГ. Перед учёными оказалась не резонансная кривая, а очень похожая на неё кривая функции корреляции информационных массивов. Тот факт, что кривая во всех случаях отображает зависимость активации биопотенциалов мозга от целостного полисенсорного образа внешнего мира, говорит как раз об объединении сигналов разных сенсорных систем по мере подъёма по этажам пирамиды принятия решений. Говорит о таком объединении сигналов в пирамиде корреляторов, которое, в конце концов, приводит к формированию единого полисенсорного образа на входе нейрокоррелятора высшего уровня.
* * *
Вспомним одно из свойств мозга, противоречащее постулатам теории “ассоциативной сети” (см. гл. 3.1.4.). По отношению к мозгу неверно считать, будто единичные изменения схемы соединений или сбои отдельных нейронов резко изменяют результат на выходе. В работе нейрокоррелятора зачастую участвуют миллиарды нейронов, а принцип вычисления функции корреляции и принцип действия голографической памяти малочувствительны к искажениям единичных элементов сравниваемых образов.
Отдельные нарушения схемы соединений или сбои отдельных нейронов эквивалентны незначительному повышению уровня шумов, не способному изменить результат корреляционного сравнения образов. Существенное ухудшение работы мозга наблюдается только при серьёзном локальном повреждении или в случаях, когда рассредоточенные нарушения в нейронах становятся массовыми.
Такой характер реагирования голографической системы мозга на сбои и отмирание отдельных нейронов вызван тем, что хаотически расположенные дефектные информационные элементы всегда оказываются не коррелированными между собой. Они не складываются в целостный образ, способный наложиться на обрабатываемый информационный массив и привести к ошибке распознавания. Так же нечувствительны к не коррелированным помехам (царапинам, точкам, пятнышкам на фотоэмульсии) и технические голографические устройства.
Иное дело – помехи работе мозга, приходящие извне. Такие помехи, по самой своей природе, коррелированны между собой и оттого более существенно влияют на результат работы нейроголографической системы. Пример – закономерное изменение уровня биопотенциалов мозга после выработки у животного условного рефлекса, происходящее, согласно рис. 3.7, при любых отклонениях параметров окружающей обстановки от привычной нормы. Биопотенциал мозга изменяется при перестановке лабораторного оборудования, переодевании лаборанта, замене посуды, в которой лежит пища, изменении освещения или звукового фона и т. д., и т. п. Изменения сигналов элементарных сенсоров в каждом таком случае коррелированны между собой, что принципиально важно.
Таким образом, устойчивость мозга к шумам, к гибели отдельных нейронов и местным повреждениям, с одной стороны, принципиально противоречит теории „ассоциативной сети”, а с другой – естественно вытекает из свойств нейрокорреляторов.
* * *
В научной литературе давно ведётся спор о степени принципиальности отличий мозга человека от мозга остальных животных. В предыдущем изложении автор рассматривал мозг вообще, как мозг произвольного животного, и при этом говорил о физиологии мышления. Выходит, автор считает мыслящими всех животных, наделённых мозгом? От человека до золотой рыбки? А как же воспринимать такие явные отличия человека, как наличие второй сигнальной системы, абстрактное мышление? Где автор проводит границу между мышлением и другими процессами в нервной системе?
Прежде всего, вспомним, что термин „мышление” имеет много ипостасей. Смысл, вкладываемый в этот термин психологом, физиологом, инженером, поэтом, наконец, домохозяйкой – далеко не одинаков. Из многих возможных подходов, в данной книге используется лишь один – мышление рассматривается с сугубо физиологической точки зрения. Можно добавить, что оно рассматривается как высший тип физиологических информационных процессов в нервной системе животного.
Известно, что лишь человеку присуще общение с помощью понятий, фиксируемых словами, математическими или графическими символами, образами художественных произведений, что связывают с физиологической системой, названной второй сигнальной. Можно ли считать, что данное различие между человеком и остальными животными, как и абстрактное мышление, с физиологической точки зрения, не принципиально, не имеет физиологической и анатомической основы?
Концепция нейроголографии даёт следующее определение.
С физиологической точки зрения, мышлением являются процессы прохождения волн массовой импульсации нейронов (волн-образов и немодулированных волн) в тканях мозга и обработка этих волн системой нейрокорреляторов.
Конечно, обнаруживается различие между привычным, довольно расплывчатым смыслом термина „мышление”, используемым разными людьми, и принятой здесь трактовкой. К мышлению отнесен определённый тип нервных процессов, главное отличие которых в массовости, в согласованном волнообразном распространении – по законам оптики – импульсации миллиардов нейронов, и в специфичном характере обработки информации – с помощью нейрокорреляторов. Все организмы, у которых существует такой тип нервных процессов (вплоть до золотой рыбки), с физиологической точки зрения, являются мыслящими.
* * *
Нервное возбуждение распространяется в мозге не по отдельным цепочкам нейронов, но с другой стороны, и не в однородном изотропном пространстве. Массовая импульсация нейронов распространяется в среде, имеющей сложное анатомическое строение и сформированной из разных типов нервных тканей с сильно отличающимися (и динамически изменяющимися!) свойствами. Именно законы распространения нервного возбуждения в этой сложной среде и определяют процессы, которые выше названы мышлением.
Каждый мозг содержит слоистую запоминающую ткань (например, в виде коры полушарий), содержит ядра (например – таламуса) как инструменты преломления и фокусировки волн нервного возбуждения, и протяжённые пучки нейронов, топологически точно переносящие волны из одних зон в другие. (Точнее было бы сказать, что пучки переносят волны от одних органов мозга к другим, но это плохо вяжется с современной терминологией, в которой сам мозг считается органом.) Процессы, использующие волны массовой импульсации нейронов в анатомических структурах мозга, обеспечивают подавляющую часть обработки сенсорной информации в организме. Такие процессы свойственны любому мозгу, и как раз в этом смысле (а не при ином понимании термина „мышление”) следует считать мыслящими всех животных, наделённых мозгом.
Нас не должно пугать различие между мышлением как физиологическим процессом и мышлением в высочайшем понимании этого термина, как наисложнейшего интеллектуального действа, каким оно фигурирует в высоких диспутах. Воздвигая стену между мышлением гениального шахматиста и, скажем, нервными процессами в мозге тритона, мы уподобились бы биологу, не признающему общности физиологии мышц Родена, высекающего из мрамора очередной шедевр, и мышц хвоста крокодила, плывущего среди тины.
Существование у человека второй сигнальной системы, использование им обобщённых понятий, членораздельной речи, математических и прочих символов не связано с появлением в мозге специальных физиологических или вычислительных процессов. И анатомически оно основано лишь на большем развитии системы ядер мозга, на резком увеличении объёма памяти, т. е. на количественных, а не на качественных отличиях.
Переход от примитивного мозга к мозгу человека происходил постепенно, через усложнение анатомии, и на этом пути трудно отыскать резкие границы. Неверно представление, будто животные вовсе не обладают обобщёнными понятиями – даже широко используемые ими сигналы тревоги выражают обобщённое понятие опасности. Не менее распространены обобщённые сигналы о наличии пищи, о занятости территории, о вызове соперника на поединок и др. Обезьяна, голосовой аппарат которой не приспособлен к членораздельной речи, в эксперименте усвоила и осмысленно использовала до 500 понятий из языка жестов глухонемых.
Точно так же, высокий уровень абстрактного мышления достигнут человеком не потому, что принципиально изменились ткани или физиология мозга, а на основе расширения возможностей присущей каждому мозгу многоуровневой системы нейрокорреляторов, где степень абстракции сигналов естественно возрастает по мере их обработки, передачи на всё более высокие ступени пирамиды корреляторов.
Не расходясь с законами диалектики, оба качественных скачка – появление второй сигнальной системы и освоение абстрактного мышления – оказались следствием количественных изменений в мозге человека по сравнению с мозгом животного.
Возможно, с точки зрения физиологической развитости, есть смысл различать, как минимум, две градации мышления: низшую – примитивное мышление и высшую – интеллект. Но и это не намного упростило бы ситуацию. Например, автор не считает, что интеллект присущ только человеку, и сегодня не готов провести чёткую грань между названными категориями. Вероятнее всего, две названные градации могут характеризовать крайние выражения такого сложного явления, как мышление, и между ними можно выделить длинный ряд переходных градаций, но нельзя найти действительно резкую границу.
3.4.7. Суть физиологии мышления
Объединяя изложенное выше, заметим, что нейробиология должна чётко делить системы промежуточных нейронов на две принципиально разные категории – типа ассоциативной сети и типа мозга (при чём, малые по числу нейронов системы типа ассоциативной сети существуют и рядом с мозгом). Ассоциативные сети используют чёткие, дискретные пути прохождения сигналов, информация выражена в них частотой нервных импульсов. В отличие от них, для мозга и мышления характерны более размытые, хаотичные пути распространения сигналов. Для них характерны также следующие особенности.
1. В основе мышления лежит использование уединённых волн (солитонов) нервного возбуждения, которые прокатываются по нервной ткани в виде волн плотности импульсации множества нейронов. Передний фронт солитона бывает промодулирован двумерным массивом сенсорной информации (образом), и в таком случае его называют волной-образом.
2. Базовой математической операцией мышления является определение корреляции между информационным массивом, переносимым волной (это может быть образ, полученный от сенсоров или от предыдущих этапов обработки, или образ, извлечённый из памяти), и массивом, находящимся в памяти. Таким вычислениям свойственна одновременная обработка больших информационных массивов за одну элементарную операцию мышления.
3. Отдельное вычисление корреляции между образами производится нейроголографическим коррелятором, типовая схема которого включает два процессорных ядра мозга, выполняющих прямое и обратное двумерные преобразования Фурье, одну из областей памяти, включённую между преобразователями, и соединяющие их пучки нервных волокон.
4. Запись и считывание образа осуществляется на основе принципов голографии. Запись в запоминающую ткань (в кору мозга) происходит при взаимодействии (в ткани) предметной волны с опорной немодулированной солитонной волной нервного возбуждения, генерируемой ядрами таламуса, а считывание – при прохождении по ткани такой же считывающей немодулированной волны.
5. Адресация образа в запоминающей ткани производится путём отклонения вектора опорной (или считывающей) волны нейронной структурой, действующей по принципу фазированной антенной решётки.
6. Элементарный осознаваемый акт мышления включает в себя прохождение через нейрокорреляторы группы из 3–10-ти волн нервного возбуждения, каждая из которых выполняет собственную функцию. Этот пакет волн обслуживает процессы записи информации и отыскания наилучшей корреляции с ранее запомненными образами. В каждом акте мышления запоминается и потому осознаётся только итог процесса.
7. В структуре мозга можно выделить подсистему принятия решений, вырабатывающую стратегию поведения животного. Многочисленные нейрокорреляторы этой подсистемы образуют пирамидальную структуру, в которой узлы нижнего уровня напрямую получают сигналы от сенсорных систем, а при переходе на верхние уровни, происходит всё большее освобождение информации от второстепенных деталей (абстрагирование) и объединение сигналов разнотипных сенсоров. Сходное строение (уже в виде перевёрнутой пирамиды нейрокорреляторов) имеет исполнительная подсистема мозга, получающая на вход сигнал о принятом решении, и вырабатывающая на этой основе, с помощью набора хранящихся в памяти типовых действий, подробные команды всем элементам исполнительных органов.
8. Неделимым информационным массивом, которым оперируют высшие отделы мозга в процессе мышления, является понятие, как совокупность данных о некотором объекте, явлении, качестве, абстрактном понятии и т. д., имеющем словесное или иное символическое обозначение. Разнообразная (в том числе – по сенсорным источникам) информация, относящаяся к понятию, благодаря большой ёмкости двумерных информационных массивов, объединяется в целостный образ, так что при обращении к символу, например, при использовании слова в разговоре, она вызывается из памяти только как неразрывное целое. Это и придаёт символу увязку с контекстом, смысловую наполненность, обеспечивает мозгу, в отличие от современных ЭВМ, осмысленное восприятие информации.
9. Однако не вся информация, используемая при формировании понятия, хранится в единственном абстрактном образе, в памяти верхних уровней „пирамиды” принятия решений. Ни один абстрактный образ верхнего уровня не мог бы существовать без информационной поддержки многими образами более низких уровней.
* * *
Опыт ЭВМ показал, что система, основанная на сложении двоичных чисел, может худо-бедно решать практически любую интеллектуальную задачу. Но если это возможно на основе сложения двоичных чисел, то, вероятно, можно найти и другие способы вычислений. Правда, выяснилось, что по отношению к мозгу не оправдала себя идея „ассоциативной сети”. Однако, может быть, есть иные варианты? Почему автор отстаивает и развивает именно концепцию нейроголографии?
Этот принципиальный вопрос требует чёткого ответа. По мнению автора, адекватность описания работы мозга концепцией нейроголографии подтверждают следующие факты.
а) Многократная нехватка памяти ДНК для описания мозга как ассоциативной сети показала, что геном описывает, в основном, не клеточный, а более высокий уровень организации мозга – уровень больших ансамблей нейронов, слоёв коры, пучков волокон, скоплений нейронов в виде ядер и т. п. Но из таких элементов, как слои, пучки и ядра можно создать лишь один тип вычислительных и, вообще, информационных систем – только оптические информационные системы.
б) На организацию мозга по типу оптической информационной системы указало и то, что вместе с мозгом Природа создала в ходе эволюции два типа нервных тканей, специально приспособленных для выполнения оптических закономерностей. Это, во-первых, серое вещество мозга, отличающееся малыми размерами нейронов, большим количеством синапсов и диффузным распространением волн нервного возбуждения, в соответствии с принципом Гюйгенса. Во-вторых, это пучки нервных волокон, перенос нервного возбуждения по которым, в информационном смысле, аналогичен переносу оптического изображения по стекловолоконным жгутам.
в) Пучки нервных волокон, топологически точно переносящие информационные массивы со своих входов на выходы, получили широкое распространение вопреки общей тенденции к повышению хаотичности нервной системы при появлении мозга. Это доказывает особую роль в работе мозга именно топологически организованных двумерных информационных массивов, т. е. изображений или образов.
г) Низкий уровень фоновой активности нервных тканей мозга (5–7%) подтвердил, что регистрируемые энцефалографами волны интегральной импульсации нейронов имеют форму солитонов. А это объяснило их способность эффективно переносить двумерные информационные массивы в виде образов, модулирующих передний фронт пространственной волны. Синусоидальные волны не удалось бы модулировать таким способом.
д) Внимание исследователей давно привлекло совпадение необычных свойств памяти мозга и голографической памяти – независимая запись разных информационных массивов в общем пространстве памяти; распределённая запись, мало чувствительная к шумам и локальным повреждениям; быстрый вызов любой информации, независимо от адреса; огромная скорость записи и считывания при медленных элементарных процессах, что говорит о записи за одну операцию целостных образов, а не последовательности данных (см. гл. 3.3.2.).
е) На эквивалентность мозга голографическому коррелятору указало полное соответствие человека свойствам оптимального приёмника, т. е. свойствам теоретически наилучшей распознающей системы, какой является голографический коррелятор при восприятии двумерных образов в шумах (см. гл. 3.4.1.).
ж) Организация мозга в виде системы нейрокорреляторов подтверждается анатомическими данными, а также точным совпадением свойств голографических корреляторов не только с сильными, но и со слабыми сторонами вычислительной системы мозга. Известно, что мозг предельно легко распознаёт образы – даже ребёнок узнаёт обстановку и людей, что относится к очень сложным задачам. Мозг быстро оценивает корреляцию между массивами данных, хорошо решает задачи в условиях неопределённости и шумов, что характерно для голографического коррелятора. Но даже искушённый в расчётах инженер с трудом решает в уме более простую задачу перемножения многозначных чисел, а это уже совпадает с трудностями выполнения арифметических операций голографическими корреляторами.
з) Расположение фоторецепторов глаза млекопитающего и особенность их коммутации, а также анатомия нервного пучка на входе наружного коленчатого тела таламуса, выявили деформацию зрительного образа, близкую к логарифмической, что характерно именно для одной из разновидностей голографического коррелятора – для коррелятора Меллина. Специфика данного коррелятора (инвариантность к масштабу и отсутствие инвариантности к сдвигу) демонстрируется и зрительной системой человека (см. гл. 3.4.5.).
и) Характер зависимости биопотенциалов мозга от изменений стимула при выработанном условном рефлексе (см. гл. 3.4.6.) подтвердил организацию системы принятия решений в виде пирамиды нейрокорреляторов, а также поэтапное объединение сигналов различных сенсоров по мере перехода на верхние уровни пирамиды, что точно соответствует концепции нейроголографии в целом.
* * *
В таком виде основы физиологии мышления мало понятны физиологам, и даже мало похожи на физиологию. Часть концепции выражена через отрицания (например, тезис о том, что работу мозга бесполезно рассматривать с позиций „ассоциативной сети”), но немногим лучше и позитивные сведения, описывающие мышление языком кибернетики и оптических вычислительных систем. Существующая же „физиология высшей нервной деятельности” оказалась далёкой от природы мозга, что отразилось и на терминологии. Новые термины в данной области ещё только будут возникать в практике исследований физиологии мышления. Как же сегодня описать мышление на языке физиологии? Приводимая ниже попытка, конечно, далека от чёткости и убедительности.
Мышление – это оформление возбуждений наших сенсоров в образы, переносимые через ткани мозга в виде волн нервного возбуждения. Это оптические преобразования таких волн-образов в ходе их преломления на границах тканей с разными скоростями распространения возбуждений. Это интерференция между новыми образами и вереницами образов, уже записанных в памяти, а также интерференция с опорными волнами, приходящими от ядер таламуса, с записью пришедших образов в память. Мышление – это рождение, в ходе преобразований и интерференции, всё новых образов по мере поднятия на верхние уровни „пирамиды принятия решений”. Рождение образов, обобщённых в ходе соединения потоков волн-образов от разных сенсоров, и упрощённых благодаря абстрагированию, исключению ненужных подробностей. Мышление – это мгновенное отыскание каждой мыслью некоего идеала, наилучшего совпадения высшего, итогового образа с одним из предъявленных ему эталонов. И как результат – выработка пирамидой нейрокорреляторов РЕШЕНИЯ, концентрирующего всю информацию и энергию данной мысли.
А затем – вторая часть пути по участку петли обратной связи, проходящему в тканях мозга. Прохождение волной-образом перевёрнутой „исполнительной пирамиды”, преобразование информации и энергии РЕШЕНИЯ в расходящиеся, дробящиеся и усиливающиеся волны-образы команд эффекторным органам. И снова – похожие преобразования образов в ходе преломления волн на границах тканей, снова интерференция новых, заданных волн-образов с вереницами образов, ранее записанных в памяти, с корректирующими сигналами афферентных нейронов и с опорными волнами от ядер таламуса, пока множество команд ни уйдёт из мозга в периферическую нервную систему и через неё – к эффекторам. Вот, что такое (сугубо упрощённо и приблизительно) мышление, с точки зрения физиологии.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
