
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Обушко Олена Миколаївна
Черкаський національний університет ім. Б. Хмельницького
Теоретичне дослідження будови та ІЧ спектру комплексу Fe(II) порфіну з киснем методом теорії функціоналу густини
Науковий напрям: 9.1 Хімія (квантова хімія).
Ключові слова: оксигемоглобін, комплекс Fe(ІІ)-порфіну з киснем, квантово-хімічний метод функціоналу густини, ІЧ спектр поглинання
Вступ
Ферум порфірини відіграють важливу роль в багатьох біологічних системах, де вони утворюють активні центри, що сприяють зв’язуванню кисню і проходженню каталітичних реакцій [1]. Добре відомо, що гемоглобін переносить кисень кровоносними судинними системами до тканин живих істот. Гем, який розглядається як активний центр гемоглобіну і міоглобіну, є комплексом порфірину з йонами Fe2+. Транспорт і акумулювання кисню можливе за рахунок оберненої реакції взаємодії кисню з гемоглобіном і міоглобіном без окиснення гемового Fe2+-йону в Fe3+-йон.
У структурі гему Fe2+-йон координаційно зв’язаний із чотирма атомами Нітрогену пірольних кілець, а два координаційні зв’язки Fe2+-йона орієнтовані перпендикулярно до площини гема. Продукт взаємодії йону металу з диоксигеном може мати різноманітну геометричну структуру. У перших дослідженнях структури гем–О2 передбачалося, що молекула О2 розташована перпендикулярно до макроциклу порфірину (модель 1). У структурі Гриффіта (модель 2) молекула диоксигену розташована паралельно до макроциклу порфірину (side-on), а в структурі Полінга – під кутом (end-on), рівним приблизно 120° (модель 3) [1].
Зв’язування кисню з Fe2+-йоном гему інтенсивно вивчається як експериментально, так і теоретично [1–10]. Але на сьогоднішній день не існує єдиної точки зору на орієнтацію кисню відносно площини порфінового макроциклу і на ступінь перенесення електронів від йону Fe2+ до молекули кисню.
Можливість існування різних форм активних центрів оксигемоглобіну може вплинути на їхні електронні властивості, а, відповідно, і на характер зв’язування гему з киснем. Тому необхідні комплексні дослідження електронних і спінових властивостей різних геометричних структур різних моделей оксигемоглобіну в різних спінових станах. Коливальні властивості активних груп можна досліджувати шляхом моделювання інфрачервоних спектрів. Зміщення частот характеристичних коливань безпосередньо пов’язано зі зміною електронного стану зв’язків і фрагментів у межах деяких локалізованих коливань, що розглядаються.
Для розрахунку рівноважної геометрії та електронної будови досліджуваних комплексів нами використано квантово-хімічний метод теорії функціоналу густини [11, 12] на рівні теорії B3LYP [13] з розширеним розщепленим базисом атомних орбіталей 6-311G.
Мета роботи полягає у проведенні теоретичного дослідження будови і ІЧ спектру комплексу Fe(II) порфіну з киснем методом теорії функціоналу густини.
Аналіз одержаних результатів важливий для з’ясування орієнтації кисню відносно площини порфінового макроциклу у міоглобіні і гемоглобіні.
Основна частина
В якості геометричної моделі оксигемоглобіну для теоретичних розрахунків нами вибрані комплекси ферум(ІІ) порфіну з киснем (Fe(II)P×××O2), де периферійні замісники в макроциклі порфірину замінені на атоми Гідрогену, а молекула кисню розташована перпендикулярно до макроциклу Fe(II)P (модель 1), паралельно (модель 2) і під кутом (модель 3). Досліджувані комплекси були нейтральними (сумарний заряд дорівнює нулю) і мали максимальну мультиплетність високоспінового стану, яка дорівнює семи (септетний стан).
При розгляді геометрії та електронної будови молекули ми користувались традиційною системою позначень [14].
Повну оптимізацію геометрії проведено для всіх досліджуваних сполук, тобто для кожної моделі знайдено повний мінімум на гіперповерхні потенціальної кривої. Досліджувані системи обговорюються в термінах електронних параметрів (заряди і спінові густини на атомах) і геометричних параметрів (довжини зв’язків, валентні і двогранні кути). Крім того, кожна система характеризується повною енергією. Для сполук 1-3 розраховані ІЧ спектри.
Можливість утворення, а також стійкість структур комплексу Fe(II)P×××O2 можна оцінювати виходячи із повної енергії комплексу. Розраховані нами повні енергії для пятикоординованих комплексів дорівнюють: -65373,768 – для комплексу 1, -65374,547 – для комплексу 2 і -65374,466 еВ – для комплексу 3. Як видно із отриманих даних, комплекс 2, де молекула О2 орієнтована паралельно до макроциклу порфіну, більш стійкий (в септетному стані). Повна енергія комплексу, в якому молекула кисню розміщена під кутом 121,0° до “площини” порфіну, лише на 0,081 еВ більша, ніж комплексу 2, а комплексу 1 на 0,779 еВ більша, ніж комплексу 2.
В комплексі Fe(II)P з киснем (сполука 1) спостерігається невелика делокалізація спінової густини на атомах Карбону і зменшення спінової густини на атомах Нітрогену. Два неспарених електрони Оксигену зберігаються майже без змін, тобто при орієнтації молекули О2 перпендикулярно до макроциклу спінова поляризація в порфіновому макроциклі значно не проявляється.
В інших моделях спінова густина перерозподіляється значно сильніше. У сполуках 2 і 3 спостерігається перенесення спіну з атомів Оксигену в макроцикл, головним чином на атом Феруму і атоми Нітрогену. Довжина зв’язку Fe – О при перенесенні спінової густини зменшується з 2,678 Å в комплексі 1 до 2,136 – в комплексі 2 і 2,084 – в комплексі 3 (рис. 1). Тобто в слабко зв’язаному комплексі 1 молекула кисню знаходиться на такій відстані від гему, що можна говорити про слабку міжмолекулярну взаємодію, а в комплексах 2 і 3 – про координаційний зв'язок між Fe і О.
Довжина зв’язку О – О змінюється від 1,254 в комплексі 1 до 1,369 і 1,329 Å в комплексах 2 і 3, відповідно; окремий розрахунок молекули О2 в такому ж наближенні ТФГ дає довжину зв’язку 1,256 Å, що знову говорить про те, що в першому випадку властивості кисню майже не змінюються. Нагадаємо, що даний метод ТФГ завищує значення довжини зв’язку в молекулі О2, оскільки за даними експериментальних значень вона становить 1,207Å [15]. В комплексах 2 і 3 довжини зв’язку О – О ближчі до їхнього значення в О2– (1,35Å [15]). Таким чином спостерігаємо, що в цих сполуках відбувається перенесення електрону з гему на кисень.
Введення хлорид-йону в комплекс 3 зменшує перенесення спінової густини з О37 на атом Fe, що призводить до послаблення зв’язку Fe – О, тобто до збільшення довжини зв’язку з 2,084 до 2,185 Å (рис. 1).
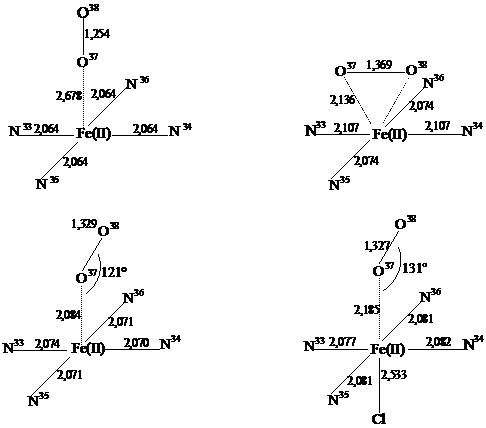
Рис. 1. Довжини зв’язків (Å) у досліджуваних моделях оксигемоглобіну
Зв’язування молекули О2 призводить до перерозподілу електронної густини у вивчених комплексах, що кількісно може бути оцінено за зміною зарядів на атомах. Як видно з рис. 2, зв’язування молекули кисню в сполуці 1 викликає невелике перенесення електронної густини на атом О37 з атомів Нітрогену (33, 34) порфінового макроциклу і з сусіднього атома Оксигену, що викликає незначну поляризацію зв’язку О – О. Така електрична поляризація викликає збільшення інтенсивності ІЧ поглинання валентного коливання зв’язку О – О в сполуці 1 порівняно з вільною молекулою О2. При цьому частота коливання зв’язку О – О практично не змінюється (1455 і 1453 см-1 в молекулі О2 і в комплексі 1, відповідно).
При утворенні комплексів 2 і 3 спостерігається інша картина, а саме значне перенесення електронної густини з Fe2+-йону порфінового макроциклу на атоми Оксигену, в результаті чого заряд Fe2+-йону в моделях 2 і 3 скомпенсований у меншій мірі порівняно з комплексом Fe(ІІ)Р, у якому заряд на йоні Феруму дорівнює 1,491 е. Молекула кисню, зв’язана з Fe(ІІ)Р, набуває значного негативного заряду (стає нуклеофільною), який в моделі 2 рівномірно розпреділений на атомах Оксигену, а в моделі 3 локалізований в більшій мірі на атомі Оксигену (О37). Таким чином порфіновий макроцикл і атом Феруму (в меншій мірі), в модельних сполуках 2 і 3 являються джерелами електронної густини. 
Рис. 2. Заряди на атомах у досліджуваних моделях оксигемоглобіну
Валентне коливання зв’язку О – О в комплексі Fe(II)P×××О2 (2) включає не тільки рух атомів вздовж осі зв’язку О – О, але також зміщення молекули О2 відносно макроциклу (валентне коливання зв’язку Fe – О). При такому русі виникає і змінюється дипольний момент. Похідна від дипольного моменту по зміщенню координати не дорівнює нулю, що і дає не нульову інтенсивність ІЧ поглинання. В комплексі 3 ефект значно посилений за рахунок поляризації зв’язку О – О і за рахунок того, що коливання цього зв’язку пов’язано з коливанням Fe – О безпосередньо, що і визначає збільшення інтенсивності із 112,5 в моделі 2 до 335,9 км/моль в моделі 3, і зміну частот коливання з 1078 – в моделі 2 до 1103 см-1 в моделі 3. При цьому в даній коливальній моді комплексу 3 з’являються деформаційні коливання СbН. Амплітуда валентного коливання зв’язку О – О у комплексах 2 і 3 дуже значна (приблизно 0,37 Å). Дуже значне посилення інтенсивності коливання О – О при зв’язуванні кисню гемом та його залежність від зв’язування показано на рис. 3, де наведені ІЧ спектри поглинання, розраховані методом ТФГ для всіх трьох моделей. Як і слідувало спостерігати, спектр комплексу 1 дуже схожий на ІЧ спектр Fe(ІІ)Р у квінтетному стані [16]. Смуга поглинання кисню, яка є суто забороненою для вільної молекули О2, дуже слабко проявляється в ІЧ спектрі даного комплексу. В комплексах 2 і 3 смуга поглинання кисню посилюється в тисячу разів і здвигається з 1453 см-1 в сторону низьких частот (1078 см-1 в моделі 2 і 1103 см-1 в моделі 3). В комплексі 3 валентні коливання О – О спостерігаються також при розрахованих частотах 1001, 1006 і 1110 см-1 (моди 68, 70 і 71), в ІЧ спектрі (рис. 3с) ці моди перекриваються дуже інтенсивною модою при 1103 см-1. За даними робіт [2, 3] валентні коливання О2 в ІЧ спектрах гемоглобіну і міоглобіну спостерігаються при 1155, 1130 і 1105 см-1. Автори роботи [4] в ІЧ спектрі комплексу ферум тетрафенілпорфіну з киснем спостерігали смугу ν(О – О) при 1106 см-1.
У високочастотній області ІЧ спектрів досліджуваних сполук, де проявляються валентні коливання С – Н зв’язків пірольних кілець і метинових містків, і в проміжній області спектрів, де проявляються скелетні коливання метинових містків та пірольних кілець, не спостерігається значних відмінностей частот та інтенсивностей коливальних мод при порівнянні ІЧ спектрів сполук Fe(II)P [17] і Fe(II)P×××О2, за винятком смуг, пов’язаних з валентними коливаннями зв’язку О – О. Однак при аналізі низькочастотної області ІЧ спектрів можна спостерігати певні відмінності. Низькочастотна мода 3 в ІЧ спектрі Fe(II)P (n = 78 см -1, І = 1,6 км/моль, у квінтетному стані), в якій під час руху відбувається утворення куполу або прогинання молекули, в ІЧ спектрах сполук 2 і 3 комплексів Fe(II)P×××О2 (рис. 3, мода 5) набуває інтенсивності (6,0 і 5,2 км/моль, відповідно), достатньої для експериментальних досліджень.
Потрібно відмітити появу в ІЧ спектрі комплексу 3 при частоті 262 см-1 малоінтенсивної смуги (мода 16, І = 4,7 км/моль), обумовленої лише валентними коливаннями зв’язку Fe – O і деформаційним коливанням FeОО (рис. 3с). Така смуга в комплексі 2 не спостерігається. Таким чином наш розрахунок дозволив передбачити основну відмінність між комплексами 2 і 3 при аналізі низькочастотної частини спектру. Валентні коливання Fe – O в комплексі 3 спостерігаються також при частотах 377 см-1 (мода 23) і 403 см-1 (мода 25), а в комплексі 2 при частотах 380 (мода 23) і 397 см-1 (мода 24). В цих модах коливання Fe – O змішуються з nas(Fe – N) зі зміщенням пірольних кілець і значним зміщенням атома Феруму. В комплексі 1 в аналогічних модах n(Fe – O) не спостерігається. В комплексі 2 валентне коливання О2 (мода 19) спостерігається також при 346 см-1 (І=7,19 км/моль), яке змішується із неплощинним коливанням CmH-груп. У ІЧ спектрі на цю моду накладаються моди 23-25 і утворюють один пік.
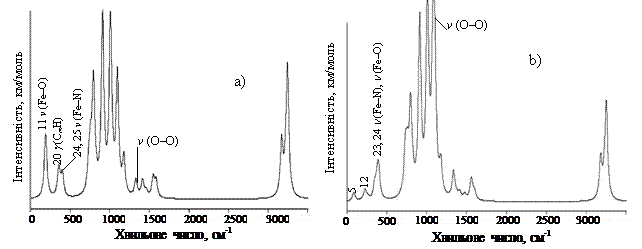 |
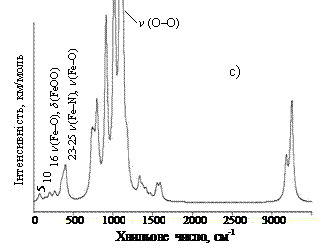
Рис. 3. ІЧ спектри поглинання комплексу Fe(II)P з О2: a) молекула О2 розміщена перпендикулярно (максимальна інтенсивність 163,8 км/моль); b) молекула О2 розміщена паралельно (максимальна інтенсивність 112,5 км/моль); c) молекула О2 розміщена під кутом (максимальна інтенсивність 335,8 км/моль). Напівширина смуг 20 см-1.
Висновки
1. Розрахунки методом ТФГ трьох відомих із літератури моделей комплексу Fe(ІІ)Р з О2 у високоспіновому стані показали, що модель 1 не відповідає експериментальним даним за ІЧ спектрами і характеризується дуже слабким міжмолекулярним зв’язуванням.
2. Моделі 2 і 3 відповідають міцним координаційним зв’язкам кисню з Ферумом і дають сильний перерозподіл як спінової, так і зарядової густини. При цьому тільки модель 3 відповідає всім експериментальним даним як за ІЧ спектрами так і за даними ЕПР і рентгеноструктурного аналізу.
3. Величезне посилення ІЧ смуги валентних коливань зв’язку О – О в комплексах Fe(ІІ)Р з О2 (моделі 2 і 3) у наших розрахунках пояснюється тим, що дана нормальна коливальна мода змішана з коливаннями порфінового макроциклу і відносним рухом кисню (ν (Fe – О)), що викликає значну зміну дипольного моменту всього комплексу.
4. Передбачена основна відмінність між моделями 2 і 3 комплексу Fe(ІІ)Р з О2, яка обумовлена появою в ІЧ спектрі комплексу 3 при частоті 262 см -1 малоінтенсивної смуги (мода 16), пов’язаної лише з валентними коливаннями зв’язку Fe – O і деформаційним коливанням FeOO. Така смуга в комплексі 2 не спостерігається.
Література
1. Неорганическая биохимия / Под ред. и .– М.: Мир, 1978.–Т. 2.–736 с.
2. Wagner W.-D., Paeng I. R., Nakamoto K. Simultaneous Observation of O-O and Fe-O2 Stretching Vibration of Fe(TPP)O2 in Dioxygen Matrices by Resonanse Raman Spectroscopy // J. Am. Chem. Soc.–1988.–110, N 16.–P. 5565–5567.
3. Potter W. T. , Tucker M. P. , Houtchens R. A, Caughey W. S. Oxygen infrared spectra of oxyhemoglobins and oxymyoglobins. Evidence of two major liganded oxygen structures // Biochemistry–1987.–26, N 15.–P. 4699–4707.
4. Watanabe T., Ama T. , Nakamoto K. Matrix-isolation infrared spectra of dioxygen adducts of iron(II) porphyrins and related compounds // J. Phys. Chem.–1984, 88, N 3.–P.440–445.
5. Jensen K. P., Roos O. B., Ryde U. O2-binding to heme: electronic structure and spectrum of oxyheme, studied by multiconfigurational methods // J. of Inorg. Biochem.–2005.–99, N 1.–P. 45–54.
6. Blomberg L. M., Blomberg M. R. A., Siegbahn P. E. M. A teoretical study on the binding of O2, NO and CO to heme proteins // J. of Inorg. Biochem.–2005.–99, N 4.–P. 949–958.
7. Franzen S. Spin-dependent mechanism for diatomic ligand binding to heme // Proc. Natl. Acad. Sci. USA.–2002.–99, N 26.–P. 16754–16759.
8. Jensen K. P., Ryde U. How O2 binds to heme // J. of Biolog. Chem.–2004.–279, N 15.–P. 14561–14569.
9. Tsuda M., Dino W. A., Kasai H. Spin Polarization Effect on O2 Dissociation from Heme-O2 Adduct // Jap. J. of Appl. Phys.–2005.–44, N 2.–P. L57–L59.
10. Kozlowski M. P., Kuta J., Ohta T., Kitagava T. Resonance Raman enhancement of FeIV=O stretch in high-valent iron porphyrins: An insight from TD-DFT calculations // J. of Inorg. Biochem.–2006.–100, N 4.–P. 744–750.
11. Momentean M., Reed C. A. ESR study of hemoglobin and myoglobin // Chem. Rev.– 1994.– 94.– P. 659–685.
12. Минав механизмы активации молекулярного кислорода // Успехи химии.–2007.–76, N 11.–С. 1039–1063.
13. Strickland N., Harvey J. N. Spin-Forbidden Ligand Binding to the Ferrous–Heme Group: Ab Inito and DFT Studies // J. Phys. Chem.–2007.–111, N 4.–P. 841–852.
14. , , Шкирман порфиринов: колебательные состояния –Мн.: Наука и техника, 1985.– 415 с.
15. -П., Константы двухатомных молекул. М.: Мир, 1984.–Ч. 2.–368 с.
16. Мінаєв Б. П., Мінаєва В. О., Говорун ідження інфрачервоного спектру молекули Fe(II)-порфіну в різних спінових станах квантово-хімічним методом функціоналу густини // Біополімери і клітина.–2007.–23, N 6. – С. 527–536.
17. Мінаєв Б. П., Мінаєва В. О., Спінові стани молекули Fe(II)-порфіну: квантово-хімічні дослідження методом функціоналу густини // Ukrainica Bioorganica Acta.–2007, N 1. –С.24–31.
