
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Глава 2. ПРИЧИНЫ, ПРИВОДЯЩИЕ К ТРАВМИРОВАНИЮ И ГИБЕЛИ ГИДРОБИОНТОВ В ПРОТОЧНЫХ КАНАЛАХ ГИДРОМАШИН
Анализ воздействия потока на рыб в камерах рабочих колес гидротурбин подробно излагается в работе , , (1999), где обобщены результаты исследований ИПЭЭ РАН и многих зарубежных ученых, изучавших на ГЭС и на лабораторных стендах судьбу рыб различных видов и возрастных групп, мигрирующих и скатывающихся через турбины ГЭС. В гл. 1 было показано, что этот этап покатной миграции ихтиофауны приводит к гибели абсолютного большинства покатников.
, и др. (1999.) проанализировали, как влияет поток на молодь рыб и планктон в проточных каналах гидромашин (турбины ГЭС, гребные винты, насосы) на основе натурных и теоретических методов исследований, рассматривая с позиций возможности решения задачи минимизации гибели гидробионтов при скате через турбины ГЭС. На наш взгляд полученные результаты в основном согласуются между собой, дополняют друг друга и могут стать основой для практического решения задач безгибельного ската ихтиофауны и планктона через проточные каналы гидромашин. Учитывая большое экологическое, экономическое и научное значение решения этой задачи, считаем целесообразным еще раз проанализировать основные факторы воздействия, приводящие к гибели рыб и планктона в турбинах ГЭС в нашем представлении, сопоставляя его с данными, приведенными в других работах (в частности и др.).
2.1. Энергетическая диаграмма рабочего процесса в проточном
канале гидротурбины и представление воздействия потока
на гидробионты
В проточной части гидромашины происходят рабочие процессы, под которыми понимают совокупность явлений, происходящих в проточной части при взаимодействии органов с потоком воды (, 1974; и др. 1984). Проточную часть можно разделить на три зоны: подвод к рабочему колесу, область рабочего колеса, включающая лопастную систему, и отвод потока от колеса. Лопастная система рабочего колеса, взаимодействуя с жидкостью, обеспечивает преобразование энергии потока в механическую энергию колеса в турбине. Момент Мк (Нм) силового взаимодействия потока с лопастной системой рабочего колеса турбины можно представить в виде:
![]()
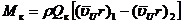 , (2.1)
, (2.1)
где ρ – плотность жидкости, (кг/м3); Qк – объемный расход жидкости, проходящий через рабочее колесо (м3/с); ![]()
![]() – средние (осредненные по расходу) значения моментов окружной составляющей абсолютной скорости, (м/с). Моменты этих скоростей с индексом 1 относятся к напорной области проточной части рабочего колеса, а с индексом 2 – к области с меньшим давлением на входе в отсасывающую трубу.
– средние (осредненные по расходу) значения моментов окружной составляющей абсолютной скорости, (м/с). Моменты этих скоростей с индексом 1 относятся к напорной области проточной части рабочего колеса, а с индексом 2 – к области с меньшим давлением на входе в отсасывающую трубу.
При взаимодействии лопастной системы колеса с жидкостью момент скорости потока уменьшается от входа к выходу, в результате чего рабочее колесо турбины отнимает энергию от потока. Величина момента Мк определяется кинематикой потока в соответствующих сечениях рабочего колеса.
Мощность N, развиваемая рабочим колесом, с одной стороны, равна произведению момента Мк на угловую скорость ротора, а с другой – произведению весового расхода ρgQ (g – ускорение свободного падения) на теоретический полезный напор Нт.
Умножив на угловую скорость выражение (2.1) и приравняв их правые части величине ρgQНт, определим теоретический напор (м):
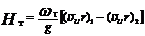
Это выражение называется основным уравнением турбины. Из него следует, что для преобразования энергии потока в турбине необходимо изменение момента скорости жидкости в области рабочего колеса. При этом в зависимости от режима работы гидромашины в проточной части рабочего колеса и сопряженных с ним элементах формируется поток определенной кинематики, а на лопастях рабочего колеса возникает соответствующее ей распределение скоростей и давлений. Последнее определяет по существу уровень гидравлических характеристик машины, в том числе таких нежелательных явлений, как кавитация с ее различными проявлениями и вихреобразование, сопровождающееся пульсацией давлений в потоке и на обтекаемых поверхностях проточной части. Вследствие этого на некоторых режимах работы наблюдаются колебания мощности гидроагрегата, гидродинамических нагрузок на элементах проточной части, вибрация гидроагрегата и кавитационное разрушение обтекаемых поверхностей.
Теоретический напор Нт гидротурбины – часть полезно используемой удельной механической энергии жидкости, т. е. преобразованной рабочим колесом в механическую энергию ротора. Он меньше напора Н1 турбины (иногда его называют располагаемым или рабочим) на величину hг гидравлических потерь в ней, т. е.
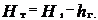 ,
,
где hг = hп + hк + hо, hп, hк, hо – гидравлические потери соответственно в турбинной камере, статоре и направляющем аппарате, рабочем колесе и отсасывающей трубе (рис. 2.1). Другими видами потерь энергии из-за их малости в данном случае можно пренебречь, и потому здесь они не учитываются.
Согласно правилам Международного кода под напором гидромашины Н(м) понимают разность удельных механических энергий жидкости в характерных сечениях ее проточной части. Для турбины это сечение на входе в турбинную (спиральную) камеру и на выходе из отсасывающей трубы,
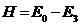
Индексами 0 и 3 обозначены соответственно сечения спиральной камеры и отсасывающей трубы. Удельная механическая энергия потока в рассматриваемом сечении проточной части выражается равенством
![]() , (2.2)
, (2.2)
Здесь ![]() – удельная энергия избыточного давления (пьезометрическая высота или пьезометрический напор), z – удельная потенциальная энергия положения (геометрическая высота или геометрический напор, иногда его называют геодезическим),
– удельная энергия избыточного давления (пьезометрическая высота или пьезометрический напор), z – удельная потенциальная энергия положения (геометрическая высота или геометрический напор, иногда его называют геодезическим), ![]() – удельная скоростная энергия (скоростная высота или скоростной напор), a – коэффициент скоростной энергии, учитывающий неравномерность распределения скоростей в рассматриваемом сечении. Каждый из этих членов измеряется в Дж/(кг м/с-2) = Дж/Н и представляет собой удельную энергию на 1 Н силы веса протекающей жидкости, но Дж/Н = Нм/Н = м – единица измерения напора. Последнее удобно для представления закономерностей движения жидкости и работы лопастных гидравлических машин. На рис.2.1 и 2.2 показано изменение удельной энергии жидкости по длине проточной части турбины.
– удельная скоростная энергия (скоростная высота или скоростной напор), a – коэффициент скоростной энергии, учитывающий неравномерность распределения скоростей в рассматриваемом сечении. Каждый из этих членов измеряется в Дж/(кг м/с-2) = Дж/Н и представляет собой удельную энергию на 1 Н силы веса протекающей жидкости, но Дж/Н = Нм/Н = м – единица измерения напора. Последнее удобно для представления закономерностей движения жидкости и работы лопастных гидравлических машин. На рис.2.1 и 2.2 показано изменение удельной энергии жидкости по длине проточной части турбины.
Разность удельных энергий воды в верхнем и нижнем бьефах при неработающих гидроагрегатах, т. е. разность отметок бьефов, называется статическим напором ГЭС. По заданной величине статического напора и потерь энергии в элементах блока, расположенных вне характерных сечений, принимаемых для определения напора, последний можно вычислить с достаточной для практических расчетов точностью:
![]() ,
,
![]() ,
,
где: hб – потери энергии в указанной выше области элементов блока турбины: hб = h1б + h2б
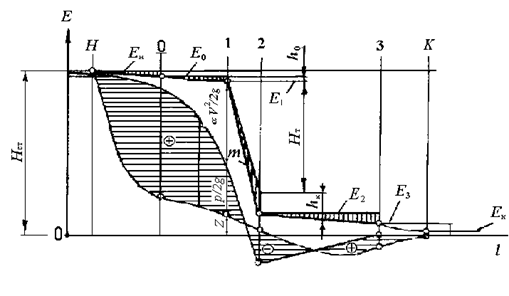
Энергетическая диаграмма рабочего процесса в проточной части гидромашины и данные экспериментальных исследований, приведенных в этой работе, дают возможность представлять и судить о воздействии потока на гидробионтов. Более того, мы попытались раскрыть механизм данного воздействия и дать ему научное обоснование, хотя следует указать, что не все детали этого воздействия раскрыты, ибо рабочий процесс в проточной части еще не изучен в полной мере особенно в зонах рабочего колеса и отсасывающей трубы. Обратим прежде всего внимание на то, что в уравнении удельной механической энергии (2.2), которая представлена диаграммой на рис. 2.2, введены средняя скорость потока по сечениям vср и коэффициент a, учитывающий неравномерность распределения скоростей.
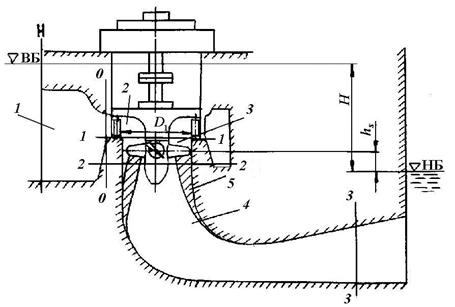
Рис. 2.1. Схема гидроагрегата и зона воздействия.
1 – подводящая камера, 2 – турбинная камера, 3 – зона рабочего колеса,
4 – отсасывающая труба, 5 – зона разрежения, К – переход к нижнему
бьефу (НБ), ВБ – верхний бьеф.
Эти параметры вводят некоторую неопределенность при оценке воздействия рабочего процесса на гидробионтов, так как величины скоростей и перепадов, а значит, и воздействий по радиусу сечения существенно не равномерны, поэтому правильнее представлять изменение удельной энергии по сечению, используя струйную теорию потока жидкости. Исходя из этой теории, удельная скоростная энергия будет изменяться по радиусу сечению в зоне рабочего колеса на входе в отсасывающую трубу, ибо здесь абсолютная скорость зависит от окружной составляющей скорости потока. Из рис.2.2 следует, что максимальные перепады давлений и скоростей потока возникают в зоне рабочего колеса на выходе в отсасывающую трубу, поэтому на этом участке будет травмироваться и погибать основная масса гидробионтов. Совершенно очевидно, что в струйках на периферии потока, где относительные составляющие окруженных скоростей максимальные, воздействие будет наибольшим, а в струйках потока у втулки – наименьшим, и, как показывают расчеты, на периферии рабочего колеса могут возникать отрицательные давления (растягивающие напряжения в потоке струйки), а в струйке ближе к оси давление при определенной высоте отсасывания hs (см. рис. 2.1), может оказаться положительным.
 |
Кроме перепада давлений, гидробионты будут подвергаться воздействию кавитации, которая, как правило, всегда проявляется в потоках рабочих колес гидромашин ( 1974; , 1974).
Рис. 2.2 Изменение механической удельной энергии жидкости по длине проточной части турбины.
Н–0 – область водоводов ГЭС; 0–1 – область турбинной камеры; 1–2 – зона рабочего колеса; 2–3 – область отсасывающей трубы; m – линия изменения удельной энергии.
2.2. Воздействие кавитации на гидробионты
Кавитационный процесс – это образование полостей в жидкости и их замыкание, который возникает в кавитационных течениях с зонами разрежения и резкими перепадами давлений (Кнэпп, 1974). Внутри среды этот процесс появляется на ядрах кавитации, которыми могут быть газовые пузырьки (газовая кавитация) либо несмачиваемые взвешенные частицы (паровая кавитация). На ядрах образуются полости в зонах разрежения, затем замыкающиеся в зонах положительного давления жидкости. На поверхности обтекаемых элементов при определенных условиях могут возникать обширные каверны (суперкавитация), при которых изменяется геометрия обтекания и которые периодически срываются и захлопываются в зонах положительных давлений. Такая кавиитация сопровождается вихреобразованиями, пульсациями давлений в потоке и на обтекаемых поверхностях проточной части. Суперкавитация в проточных каналах гидромашин не допускается. Поэтому эксплуатационные параметры гидротурбин назначаются с учетом ограничений по кавитации, для чего проводятся обязательные кавитационные испытания на специальных стендах моделей рабочих колес, при которых определяются их кавитационные свойства.
Кавитационные свойства гидротурбин характеризуются кавитационным коэффициентом σТ, который пропорционален отношению динамического разрежения на лопастях рабочего колеса к напору. Определение σТ сводится к нахождению минимально предельных значений высот отсасывания hS (см. рис. 2.1) во всем поле энергетической характеристики гидротурбины, при которой еще не меняются основные параметры модели турбины: момент на валу рабочего колеса, напор, расход, частота вращения.
Условия возникновения кавитации в гидротурбинах устанавливаются через барометрическое давление В, высоту отсасывания hS, напор Н и НВп – давление насыщенных паров воды при данной температуре. Рассчитывается величина, которая называется коэффициентом установки:
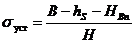
Кавитация в гидротурбинах не наступает до тех пор, пока σуст ≥ σт.
Это условие не следует понимать в прямом смысле. Здесь имеется в виду, что отсутствие кавитации при выполнении указанного условия означает следующее: при высоте отсасывания hS в проточной части турбины не возникают такие сплошные каверны с насыщенными парами воды, которые могли бы исказить геометрию обтекания проточных элементов турбины и привести к изменению ее эксплуатационных параметров. Существует начальный этап кавитации, так называемый пузырьковый, который не оказывает влияния на параметры турбин, но разрушает материалы деталей проточной части турбины (кавитационная эрозия). Такая кавитация проявляется почти на всех гидромашинах, в том числе и в турбинах низконапорных ГЭС, и, как правило, за рабочим колесом на границе с камерой отсасывания трубы. Примером может служить турбина Саратовской ГЭС (напор 10 м), где разрушается от кавитации звено стальной обшивки, переходящее в камеру отсасывающей трубы. Рассмотрим более подробно кавитацию, которая постоянно сопровождает рабочий процесс при эксплуатации гидромашин.
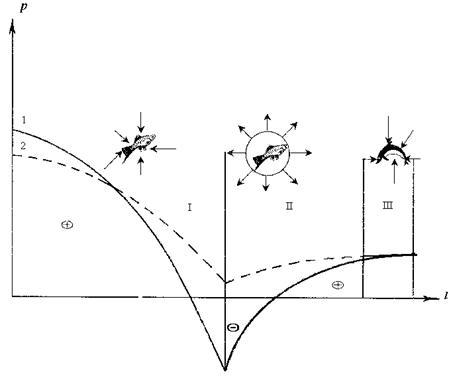 |
Рис. 2.3. Механизм воздействия на гидробионты перепадов давлений кавитационного (1) и безкавитационного (2) течений в проточном канале гидротурбины.
I – зона сжатой жидкости до рабочего колеса и воздействия положительного давления на гидробионтов; II – зона образования каверны и воздействия разрежения за рабочим колесом; III – зона захлопывания каверны, сжатой жидкости в отсасывающей трубе.
Согласно диаграмме удельной энергии, поток воды имеет три основные зоны (рис. 2.2): I ‑ зона сжатой жидкости до рабочего колеса, II – зона разрежения за рабочим колесом, III – зона сжатой жидкости в отсасывающей трубе. Ядрами кавитации могут быть, как принято считать, пузырьки воздуха либо взвешенные частицы с несмачиваемой поверхностью. В научной литературе обсуждаются различные гипотезы об источниках зародышей кавитации. Это связано с тем, что в естественных условиях зародыши кавитации в виде пузырьков воздуха и частиц очень малы по размерам (доля микрометра) и на таких ядрах в работающих гидромашинах не может развиваться кавитация, тогда как элементы проточной части машины разрушаются от кавитации. Большие пузырьки (если по какой-либо причине они появятся), на которых может развиться кавитация, обязательно всплывут (например, в верхнем бьефе ГЭС), а крупные частицы осядут на дно.
Ядрами кавитации могут быть планктонные организмы и рыбы. Диапазон размеров их от 1 мкм и более до больших, их поверхность мало - и несмачиваемая. Это вполне подходящие ядра, на которых может развиваться кавитация в проточных каналах турбин ГЭС, винтах судов и в насосах.
Поясним, как развивается кавитация на рыбах или планктонных организмах в проточном канале турбины. На рис. 2.3 изображены две кривые абсолютных давлений в сечении 1–1 (см. рис. 2.1): 1 – кривая изменения давления в струе кавитационного течения (периферия лопасти), 2 – кривая изменения давления в струе безкавитационного течения (область корневого сечения лопасти на втулке). В струйке 1, имеющей отрицательные давления возможна кавитация, а в струйке 2, без отрицательного давления кавитации не будет. На рис. 2.3 схематично изображено прохождение рыб в кавитационном и безкавитационном потоках.
Рыба (либо планктон) в зоне положительного давления сжата жидкостью и, если рыба адаптирована к этому давлению, с ней ничего не произойдет. Затем живой организм мгновенно попадает в зону разрежения (кривая 1), где произойдет отрыв жидкости от поверхности гидробионта с образованием вокруг него каверны с давлением насыщенных паров воды. Через доли секунды данная каверна, войдя в зону сжатия, мгновенно захлопнется. При этом произойдет такой же мгновенный процесс конденсации паров, который усилит удар. Микроструя жидкости при захлопывании разрушит либо травмирует организм. Таким образом, организмы в этой струе 1 будут подвергаться воздействию перепада давления и кавитационного удара и их травмирование и гибель неизбежны. В струе 2 кавитация не возникает, так как при положительном давлении отрыв жидкости (образование каверны) от поверхности организма не происходит. Здесь организм будет подвергаться воздействию только перепада давления, что создает шанс пройти гидробионту этот опасный путь без повреждений.
Так как за рабочим колесом турбины разрежение возникает в большей части объема потока, в кавитационный процесс будет включаться большая часть проходящих через турбину гидробионтов.
Если известна величина разрежения жидкости, можно определить, на каких размерах планктона при этом давлении будет образовываться кавитационная полость, т. е. какие обитатели воды погибнут. Соотношение между давлением р и характерным размером планктона d, на котором образуется сферическая полость диаметром d, можно представить в виде.
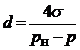 (2.3)
(2.3)
где рн = 0,002 МПа – давление водяных паров; s = 7,5∙10 -4 Н/см –поверхностное натяжение воды. Это соотношение справедливо при р £ 0.
Зависимость между d и p согласно (2.3) представлена на рис 2.4, где наглядно иллюстрируется воздействие кавитации на группы планктона, классифицированных по размеру. В высоконапорных ГЭС (Н = 100–300 м), где разрежение может достигать 0,3 МПа, кавитация будет развиваться не только на нанопланктоне, но и частично на ультрапланктоне. Это значит, что в таких турбинах полностью погибнет зоо - и фитопланктон. Возможна также трансформация бактериопланктона. При величине разряжения, близкой к нулю, что характерно для низконапорных ГЭС, кавитация может развиваться на планктонных организмах с размером 150 мкм.
 |
Это теоретическая оценка воздействия кавитации на гидробионты в кавитационных течениях гидромашин. Возможно она недостаточна в силу того, что представленная аналитическая зависимость не учитывает ряд других факторов в сложном физическом процессе возникновения и развития кавитации. Здесь важно было раскрыть механизм воздействия кавитации на ихтиофауну и планктон, Именно это позволило сформировать основной принцип защиты гидробионтов от гибели в кавитационном течении гидромашин.
Рис.2.4. Образование кавитационной каверны на планктоне и рыбах:
а- ультрапланктон; б- нанопланктон; в- микропланктон; г - мезопланктон и рыба
2.3. Результаты лабораторных исследований воздействия на рыб перепадов давлений и кавитации
В монографии , и (1999) обобщены и приведены весьма ценные результаты экспериментальных исследований воздействия перепадов давлений, кавитации и турбулентности на рыб различных видов, возрастов и размеров, а также анализа травмирования рыб при их контакте с конструктивными элементами турбин. Дана сравнительная оценка действия различных факторов на травмирование и гибель рыб при их миграции через турбины ГЭС. Обобщение материалов российских и зарубежных авторов охватывают почти 70-летний период, когда с 1930-х годов ХХ в. стали изучать это явление.
Рассмотрим основные результаты, изложенные в ней. Рыбы по-разному воспринимают воздействие на них потока в проточном канале турбины, и это зависит от их вида, возраста и в значительной мере от того, к какой глубине они адаптированы перед проходом через турбину. Исследования проводились для четырех характерных режимов изменения давления – компрессии, компрессии с последующей декомпрессией, декомпрессии рыб, адаптированных к поверхности и глубине.
Компрессия. Большое количество экспериментов было выполнено по влиянию повышенного давления на рыб. Harvey (1963) подвергал мальков и смолтов нерки очень высокому давлению (2064 кПа) и не получил значимой величины смертности. Nishiyama (1965) указывает на то, что при давлении, близком к 2500 кПа, у рыб наблюдались конвульсивные движения и нарушения дыхания, а Quasim с сотр. (1963) сообщают о нарушениях в ориентационной способности рыб. Их данные пополнили результаты многих других исследователей (Calderwood, 1945; Lucas, 1962; Muir, 1959, и др.), отмечающих толерантность рыб к высокому давлению. Все они сходятся в том, что даже значительное повышение давления изменяет поведение рыб, но не приводит к их мгновенной гибели.
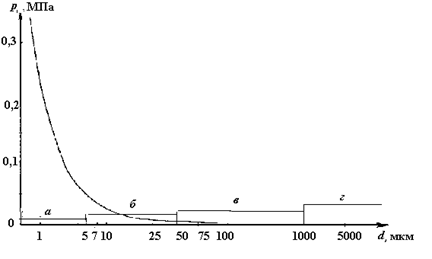
Компрессия с последующей декомпрессией. Некоторые исследователи в своих опытах сначала увеличивали давление, а затем снижали его до прежней величины (рис. 2.5). Такой режим изменения давления можно наблюдать на начальном участке турбинного тракта. Он характерен для рыб, обитающих в поверхностных слоях воды, когда их переносят в заглубленные горизонты с последующим подъемом к поверхности.
В ряде работ (Calderwood, 1945; Lucas, 1962; Gordon, 1970) отмечается, что медленная компрессия и декомпрессия не оказывают существенного влияния как на тихоокеанских, так и на атлантического лососей.
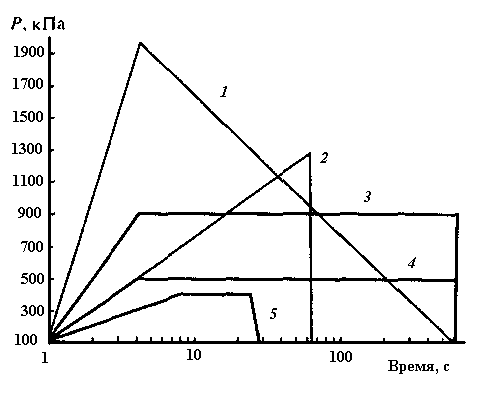 |
Рис. 2.5. Режимы изменения давления в опытах
(по данным литературных источников):
1 – Foye, Scott, 1965; 2 – Rowley, 1955; 3 – Федяй, 1981; 4 – Lampet, 1976;
5 ‑ Quasim et al., 1992.
Rowley (1955), использовал мальков микижи, повышал давление от атмосферного[1] до 1276 кПа менее чем за 1 мин, а затем мгновенно сбрасывал его до прежнего уровня. При повышении давления рыбы замирали и оставались неподвижными. Однако сразу после его снижения до первоначального двигательная активность рыб восстанавливалась, и они не гибли. Foye и Scott (1965) также не наблюдали гибели у шести видов пресноводных: полосатого окуня, желтого окуня, семотилуса, обыкновенного шайнера, озерной форели и атлантического лосося, подвергая их действию мгновенного увеличения давления до 1964 кПа с последующей декомпрессией до атмосферного давления за 10-минутный период.
Lampert (1976) использовал в экспериментах мальков сига и обыкновенного карпа. Он повышал давление до 500 кПа, с 10-минутным периодом адаптации рыб, а затем понижал до атмосферного. Такой режим изменения давления также не вызывал гибели рыб. Очевидно, это связано с малым временем их адаптации к повышенному давлению. Аналогичные данные получены (1981) при 10-минутной адаптации годовиков синца и взрослых особей колюшки к повышенному давлению в 300–900 кПа и сбросом его до атмосферного. Turnpenny с сотр. (1992) в течение 5 с увеличивали давление до 405 кПа и выдерживали сельдь, шеда, лосося, кумжу, микижу, окуня и угря в этих условиях в течение 15 с, после чего снижали давление до атмосферного. Это не вызывало у рыб травмирующих эффектов.
Толерантность рыб к указанным режимам изменения давления, по-видимому, связана с тем, что при компрессии они не успевали адаптироваться к высокому давлению, и по этому в процессе декомпрессии никаких физиологических изменений в тканях и органах рыб не происходило. На основании этих данных можно сделать вывод о том, что резкая компрессия с последующей быстрой декомпрессией до уровня атмосферного давления не приводит к гибели рыб.
Декомпрессия рыб, адаптированных к поверхностным горизонтам воды. В опытах с резким изменением давления до величин, ниже атмосферного, и быстрым его подъемом (рис. 2.6) многие исследователи наблюдали гибель рыб. Смертность в этих условиях во многом зависела от давления адаптации рыб (горизонта их обитания).
Одним из первых, кто подвергал пресноводных рыб различным режимам воздействия вакуума, был Hogan (1941 г.). Он адаптировал их к атмосферному давлению, а затем снижал его до 17 кПа в течение 15 с. Открытопузырные рыбы – гольян, обыкновенный карп и длиннорылый панцирник – переносили такое изменение давления лучше, чем закрытопузырные (синежаберный солнечник, карпии и большеротый окунь), большая часть которых погибла. Причем чем длительней была экспозиция в условиях вакуума, тем выше была их смертность. Ни одна из открытопузырных рыб не погибла. При декомпрессии эти рыбы выделяли газ из плавательного пузыря через пневматический проток. Его эксперименты показали, что резистентность к понижению давления у открытопузырных рыб выражена гораздо сильнее, чем у закрытопузырных.
Turnpenny et al. (1992) в течение 10 с увеличивали давление от атмосферного до 405 кПа, а затем резко (за 0,1 с) снижали до вакуума (15 кПа). Они выдерживали рыб при этом давлении 30 с, а затем увеличивали его до атмосферного. В этих условиях повреждений у рыб не наблюдалось. Но когда они такую процедуру проделали очень быстро, в течение 5 с, у 10% рыб (лосось, кумжа и микижа) плавательный пузырь был поврежден. У сельди травм не было. Авторы отнесли это к способности открытопузырных рыб при декомпрессии быстро удалять из плавательного пузыря излишки газа. Тем не менее они пришли к выводу, что даже для этих рыб при достаточно высокой и быстрой декомпрессии возможен разрыв плавательного пузыря, когда скорость увеличения его объема будет превышать возможности вентиляционной системы.
 |
Рис. 2.6. Режимы изменения давления в опытах
(по данным литературных источников).
1 – Harvey (1963); 2 ‑ Turnpenny et al. (1992); 3 ‑ Turnpenny et al. (1992); 4 – Hogan (1941).
Harvey (1963) подвергал смолтов нерки очень быстрой декомпрессии (52кПа/с), которая приводила в некоторых случаях к их гибели. При давлении менее 67 кПа уже наблюдалась гибель рыб, которая составила 2%. Он установил, что скорость декомпрессии существенно влияла на смертность рыб. Постепенное снижение давления до 16,5 кПа не приводило к травмированию рыб. Кроме того, он подвергал рыб, адаптированных к поверхностному слою воды, резкому повышению давления с последующей быстрой декомпрессией до вакуума и фиксировал небольшую величину смертности рыб.
Все рассмотренные опыты показали, что резкая декомпрессия может вызвать гибель рыб, обитающих в поверхностных горизонтах. Ее причина связана с разрывом плавательного пузыря. Причем смертность рыб напрямую зависит от степени декомпрессии и скорости изменения давления. С увеличением этих показателей число погибших рыб возрастает, при этом больше страдают закрытопузырные рыбы, которые не могут быстро привести в равновесие давление газа в плавательном пузыре с давлением окружающей среды.
Декомпрессия рыб, адаптированных к глубине. Harvey (1963) для рыб, адаптированных к давлению 2000 кПа, очень быстро (52 кПа/с) снижал его до 16,5 кПа. В этих условиях резкой декомпрессии смертность смолтов нерки составила 35%. Основной причиной гибели рыб являлась газовая болезнь. В большинстве случаев она поражала сердце или брюшную аорту.
В опытах (1981) резкое (90 кПа/с) снижение давления от 300, 700 и 900 кПа до 20 кПа вызвало значительную (до 90%) гибель личинок щуки, плотвы и годовиков синца, которая связана в основном с развитием газовой болезни.
Таким образом, для рыб, адаптированных к нижним горизонтам воды, резкое уменьшение давления до вакуума в районе рабочего колеса турбины может приводить к их гибели, обусловленной развитием газовой болезни.
Рыбы, адаптированные к глубине, испытывают также изменение гидростатического давления, при котором они в нижнем бьефе гидроузла оказываются на значительно меньших глубинах, чем в верхнем бьефе. Исследования такого действия перепада давления на рыб можно найти в ряде отечественных работ ( и др., 1972; , , 1974; Павлов и др., 1981), которые показали, что после длительной адаптации рыб к повышенному давлению (200–700 кПа) и резкого (10–600 кПа/с) его снижения до атмосферного изменяется поведение рыб. Они, как правило, находились у поверхности воды с нарушенным дорзовентральным положением тела, и у большинства из них была понижена реакция на внешние раздражители. Кроме нарушения поведения, отмечались и характерные травмы, вызванные действием давления ( и др., 1981), например выпучивание глаз (особенно это свойственно окуневым рыбам) и разрыв стенок плавательного пузыря (у карповых чаще на его вентральной стороне, а у окуневых – в зоне газовой железы). В пищеварительным тракте можно было встретить, независимо от состояния плавательного пузыря, скопление пузырьков – это, видимо, часть сбрасываемого из пузыря газа или заглоченного воздуха при компрессии. Кровеносные сосуды в основаниях плавников расширены и в большинстве случаев разорваны. Обнаружены и внутриполостные кровоизлияния. В глазах, жабрах и слизистой оболочке жаберной полости имелись пузырьки газа, а в некоторых случаях – целые скопления пузырей, образующих комок пены.
Смертность в экспериментах была различной у рыб разных видов и размеров ( и др., 1981). Так, при мгновенном изменении давления в 300 кПа для рыб длиной 50–100 мм она составила (%): у балхашского окуня – 9, у чебачка – 14, у сазана – 70, а у белого амура – 80. Для молоди судака длиной 10–19 мм при таком же изменении давления летальный исход отмечен только у 6% исследованных рыб, а гибели молоди амурского бычка не отмечалось. В то же время у молоди чебачка длиной 35–49 мм при перепаде в 300 кПа погибало 42%, длиной 50–75 мм – всего 14%. Эти опыты показали, что характер травм и смертность рыб зависит не только от их видовой принадлежности и возраста. Во многом они определяются величиной перепада давления и интенсивностью его изменения. Например, смертность у молоди амурского чебачка длиной 35–49 мм составляла при перепаде в 200кПа 14%, в 350 кПа – 71% и в 400 кПа – 100%. Гибель рыб во многом зависела и от скорости изменения давления. Так, для молоди сазана длиной 75–90 мм при интенсивности 250 кПа/с она составила 50%, а при 20 кПа/с рыба не гибла.
Аналогичные исследования провели Feather, Knable (1983), которые адаптировали большеротого окуня к давлению 191, 280 и 368 кПа, а затем быстро снижали его до атмосферного. Смертность рыб в опытах была напрямую связана с величиной декомпрессии и при давлении адаптации 191 кПа составила 25%, а при 369 кПа – 46%. Причем, если при высоком исходном давлении (адаптации) большинство рыб гибло в течение первого часа, то при меньшей величине декомпрессии летальные исходы наблюдались первые пять дней. Причины смерти у рыб, адаптированных к разной величине давления, были неодинаковыми. При минимальной декомпрессии (191 кПа) гибель рыб происходила в результате повреждения дыхательных органов и из-за стресса, вызванного их сильной положительной плавучестью. Декомпрессия от больших величин давления (369 кПа) вызывала газовую болезнь, которая выражалась в разрыве кровеносных сосудов и образовании крупных газовых пузырей в области сердца, жабрах и мозге. Результаты этих опытов показали, что даже сравнительно небольшое, но резкое снижение давления может стать причиной гибели рыб.
Имеющиеся в распоряжении авторов литературные данные, отражающие влияние снижения давления на рыб, обобщены и представлены на рис. 2.7, на котором показано изменение показателя смертности рыб в зависимости от степени декомпрессии
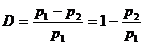 .
.
Она показывает величину снижения давления относительно адаптационного для рыб. При анализе были использованы результаты, полученные для рыб старше одного года из трех семейств (окуневые, карповые и лососевые) и только тех опытов, в которых декомпрессия от давления адаптации (р1) до минимального (р2) происходила очень быстро (менее чем за 5 с). Такой режим изменения давления характерен для турбин, где наблюдается резкое по интенсивности и большое по абсолютной величине понижение давления. Из представленных материалов следует, что с увеличением степени декомпрессии смертность рыб возрастает. Она больше у закрытопузырных рыб и рыб, адаптированных к заглубленным горизонтам воды. В меньшей степени от действия декомпрессии страдают лососевые и сельдевые, у которых имеются возможности быстрой газорегуляции плавательного пузыря. Так, D = 0,85 гибель сельдевых не наблюдается, а при D = 0,91 их гибнет только 4% (Turnpenny et. al., 1992). У карповых при одной и той же степени декомпрессии смертность больше, чем у лососевых, но несколько меньше, чем у таких ярких представителей закрытопузрных рыб, как окуневые. У последних, даже у рыб, адаптированных к поверхностным горизонтам воды, при D > 0,8 отмечена гибель до 100%.
Можно считать, что смертность рыб становится значительной при D >0,6. По-видимому, эту величину степени декомпрессии можно считать критической и использовать ее в качестве критерия для оценки гибели рыб в результате перепада давления на рабочем колесе турбины.
К такому же выводу пришли в своей работе Cada и др., (1997), а некоторые исследователи (см. Bell, 1990; Development ARL 1996) считают, что для лососевых этот показатель может быть увеличен до 0,7.
Травмирование и гибель рыб во многом зависят от скорости декомпрессии.
Разброс данных, приведенных на рис. 2.7, можно объяснить не только видовой принадлежностью испытуемых рыб, но и различной интенсивностью действия давления. Она в представленных опытах изменялась от мгновенного сброса давления до 54 кПа/с. Следует отметить, что исследований по влиянию скорости декомпрессии на рыб очень мало. Их результаты в основном сводятся к тому, что с ее увеличением число погибших рыб возрастает. Однако, учитывая то, что большинство опытов проведено в различных условиях и разнообразными методами, обобщить их не представляется возможным. Поэтому для оценки влияния скорости декомпрессии на смертность рыб ограничились небольшим набором данных (рис. 2.8), где это влияние изучалось, что называется, в явном виде. Так, для плотвы и верховки, адаптированных к давлению 400 кПа, снижение давления с разной скоростью до 100 кПа (атмосферное) приводило к неодинаковым результатам ( и др., 1972). При скорости декомпрессии 300 кПа/с погибало 100% мальков плотвы длиной 20–25 мм, при 30 кПа/с – 56%, а при 10 кПа/с – только 10%. У верховки длиной 15–25 мм в исходных условиях смертность была значительно ниже. Для ювенильных особей лососевых рыб доля погибших еще ниже. Она превышает 10%-й уровень только при скорости декомпрессии более 70 кПа/с. Если учесть, например, что рыбы проходят турбинный тракт менее чем за 15 с, можно оценить степень воздействия на них перепада гидростатического давления. Так, для рыб, адаптированных к глубине 30 м, при скорости декомпрессии в 30 кПа/с может погибнуть 15% верховки и 52% плотвы, а лососевые рыбы в этом случае не гибнут.
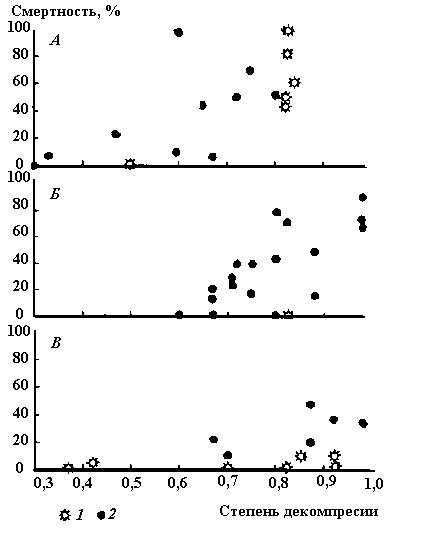 |
Рис. 2.7. Зависимость смертности рыб от степени декомпрессии
(по данным разных авторов):
А – окуневые; Б – карповые; В – лососевые. Адаптация рыб: 1 – к поверхностным горизонтам, 2 – к придонным горизонтам
 |
Травмирование и гибель рыб на ГЭС во многом зависят от периода онтогонеза. Личинки, обитающие у поверхности воды, по-видимому, в меньшей степени будут страдать от воздействия давления. Beck с сотр. (1975) подвергали икру и личинок полосатого окуня изменению давления в разных комбинациях. Их выдерживание в условиях разряженного давления (44 кПа) приводило к некоторой дополнительной гибели по сравнению с контролем. Повышение давления от 01.01.01 кПа не вызвало гибель личинок. В опытах с быстрым понижением давления на 86% относительно адаптационного было зарегистрировано незначительное количество погибших рыб. Cada с сотр. (1980) получили аналогичный результат для личинок большеротого окуня и канального сомика, которых подвергали резкому воздействию давления в диапазоне от 56 до 146 кПа. Kedl, Coutant (1976) экспериментировали с личинками ушастого окуня, обыкновенного карпа, белого окуня и полосатого окуня при резком изменении давления от 2000 до 5 кПа, а Ginn с сотр. (1978) подвергали личинок обыкновенного карпа быстрому действию декомпрессии от 100 до 53 кПа. Во всех этих опытах гибель рыб не отмечалась.
Рис. 2.8. Смертность рыб в зависимости от интенсивности изменения давления
(по литературным данным).
1 – плотва (длина 20–25 мм); 2 – верховка (длина 15–25 мм);
3 – лососевые (длина 60–80 мм).
Тем не менее для молоди, обитающей в толще воды и у дна, смертность может быть достаточно высокой. Так, для личинок плотвы, на этапе С2, адаптированных к повышенному давлению, перепад в 100 кПа приводит к гибели 67% рыб ( и др., 1972). Более 90% личинок щуки и плотвы погибало в опытах (1981) при быстром снижении давления от 300–900 кПа до атмосферного. По данным Jones (1951), поздние личинки и ювенильные особи с развитым плавательным пузырем, забираемые с глубин более 10 м, могут погибнуть под воздействием быстрой декомпрессии.
Таким образом, фактор изменения давления может оказывать губительное воздействие на рыб при прохождении ими турбинных трактов. Они травмируются и гибнут в основном при резком снижении давления. Обитатели поверхностных горизонтов в меньшей степени подвержены действию декомпрессии при прохождении турбины. Такой режим изменения давления наиболее опасен для обитателей глубин. Как правило, у них травмируется плавательный пузырь. Его стенки могут разрываться, так как при снижении давления происходит значительное увеличение объема газа. Величина критического давления декомпрессии, при котором происходит этот разрыв, во многом определяется системой газорегуляции и жесткостью оболочки пузыря. По этому смертность рыб в данном случае зависит от их вида и возраста. Она различна для открыто - и закрытопузырных рыб. Последние страдают в большей степени, так как не могут быстро уравнять давление газа в пузыре и давление окружающей жидкости. Количество газа в плавательном пузыре тем больше, чем глубже место обитания (адаптации) рыб; отсюда их смертность, вызванная нарушением целостности этого органа, возрастает с увеличением исходного (адаптационного) давления и степени декомпрессии. Другой причиной гибели мигрантов является газовая болезнь, которая наблюдается при быстрой декомпрессии. Отрицать ее наличие у рыб нельзя, так как большинство травм в условиях действующих ГЭС связано с кровоизлияниями, вызванными этой болезнью. По мнению В. Lynwood (1974), от нее на ГЭС гибнет от 20 до 40% покатников лососевых рыб.
Для того чтобы снизить губительное действие давления на рыб, необходимо идти по пути уменьшения степени декомпрессии и не превышать ее критическое значение.
Воздействие кавитации. Многие исследователи отмечают губительное воздействие кавитации на рыб, но экспериментальных работ в этом направлении очень мало, что по видимому, связано с трудностями моделирования в лабораторных условиях эффектов кавитации. Наиболее полно такие работы проводил Muir (1959). Он выдвинул гипотезу о том, что гибель рыб в турбине в основном вызвана действием кавитации и смог смоделировать ее действие на лабораторном стенде. Кратковременная выдержка молоди кижуча длиной 66 мм при таком давлении, когда вода превращается в пар, не вызвала их гибели. Однако, когда их подвергли мгновенной декомпрессии в течение 0,4 с до величины давления парообразования, а затем вернули к атмосферному (это привело к созданию кавитационных каверн), наблюдался летальный исход более чем у 60% тестируемых рыб. Визуальный осмотр показал наличие у погибших рыб кровоизлияний в глазах и на жаберных пластинах. В результате своих экспериментов Muir пришел к заключению о том, что гибель рыб от кавитации является результатом удара, вызванного взрывным действием срыва кавитационных каверн.
Cramer, Oligher (1964) сообщили, что при проведении натурных экспериментов наличие кавитации приводило к дополнительной гибели рыб в турбинах Френсис на 52–93% и турбинах Каплан – на 66–85%.
Травмирование рыб при их контакте
с конструктивными элементами турбин
Авторы, изучавшие этот фактор, пришли к выводу, что столкновение рыб с элементами конструкции турбин может приводить к гибели крупных особей, но она маловероятна и менее губительна для рыб с малой длиной тела, например ранней молоди. Помимо параметров тела рыб и характера их распределения в турбинном тракте, вероятность контакта мигрантов с лопастями рабочего колеса во многом определяется кинематикой потока и турбины, а также ее конструктивными элементами (числом лопастей, расстоянием между лопастями рабочего колеса, зазором между кромками лопаток направляющего аппарата и лопастями турбины).
При оценке этого фактора также надо иметь в виду, что площади сечений проточных элементов, где могут происходить столкновения с рыбами, на порядок и более меньше площадей сечений потока.
Большой процент гибели следует ожидать среди крупных особей рыб. При скате молоди рыб, как показывают наши исследования, травмы от столкновений получают не более 5% рыб.
Многие авторы изучали воздействие на рыб турбулентности потоков в жидкости, для чего пропускали их с различными скоростями по трубам, помещали в бассейн и воздействовали на них высокоскоростной струей, использовали в эксперименте вращающиеся в разные стороны цилиндры и другие установки, в которых создавались турбулентные потоки с образованием сдвигающих напряжений. Было установлено, что турбулентные потоки, возникающие в некоторых зонах проточного тракта турбины, могут влиять на травмирование и гибель рыб. Однако при учете этого фактора следует иметь в виду, что объем воды с зонами турбулентности на порядок, а может быть и более, меньше расходного объема проходящей воды в этих зонах, так как проточные элементы рассчитываются и проектируются из условия ламинарности потока при их обтекании. Поэтому, по нашему мнению, турбулентностью как фактором травмирования и гибели рыб при оценке воздействия турбин ГЭС на ихтиофауну можно пренебречь.
Главные факторы воздействия потока
Результаты лабораторных и натурных исследований, их анализ, сопоставление с данными проведенных нами исследований дают основание сделать вывод, что главными факторами воздействия потока в турбине на живые организмы являются перепады давлений и кавитация, при этом фактор столкновения с проточными элементами турбины следует учитывать при скатывании крупных особей рыб.
Главные факторы воздействия можно обосновать в кратком изложении следующими результатами научных исследований:
1. Все турбины ГЭС эксплуатируются в режимах с начальным этапом кавитации, которая неизбежно возникает на периферии выходных кромок лопастей. Каверны, которые образуются на живых организмах как на ядрах кавитации, за лопастями захлопываются, разрушая малые по размерам организмы: планктон и ихтиофауну. В пробах на планктон всегда обнаруживаются мелкие фрагменты разрушенных организмов. По данным испытаний на Усть-Илимской ГЭС (Н = 85 м) и Волжской ГЭС (Н = 20 м) в кавитационных течениях потока, где создаются резкие перепады давлений с отрицательными напряжениями в жидкости (см. рис. 2.3, кривая 1), около 30% биомассы зоопланктона разрушается в мелкую структуру.
2. Muir (1959) на стенде впервые смоделировал действие кавитации на молодь кижуча, которых подвергли мгновенной (в течение 0,4 с) декомпрессии до парообразования, а затем вернули к атмосферному давлению. Летальный исход был у 60% тестируемых рыб. Он на основании опытов пришел к заключению, что гибель рыб в турбине в основном вызвана действием кавитации (взрывное действие при захлопывании каверны). Видимо, каверна в его опытах образовывалась на всей или большей части поверхности подопытных рыб. Если размер рыб относительно большой, энергии хлопка может оказаться недостаточно для разрушения рыбы на мелкие части. В планктонных пробах на Волжской ГЭС в нижнем бьефе попадались тела молоди рыб в сильно «помятом» состоянии.
3. Испытания в опытах на стендах с резким изменением давления до величины ниже атмосферного (до нуля абсолютного давления) и быстрым его подъемом (см. рис 2.6) показали, что такое воздействие приводит к гибели рыб и в значительной мере тех, которые были адаптированы к глубине. В проточном канале турбины такое воздействие возникает в безкавитационных течениях (см. рис. 2.3, кривая 2), когда, без образования каверны мгновенно происходят декомпрессия в камере рабочего колеса и затем быстрый подъем давления в камере отсасывающей трубы. Гибель рыб в проточных каналах турбин с характерными травмами воздействия кавитационных и безкавитационных течений подтверждается натурными испытаниями. Рыбы могут пройти турбину без летального исхода у втулки рабочего колеса, где интенсивность перепада давления минимальна.
[1] Здесь и далее указано атмосферное давление на поверхности воды в экспериментальной емкости.
