

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
ВЛАДИМИРЦЕВА Мария Всеволодовна
ЭКОЛОГИЯ СТЕРХА GRUS LEUCOGERANUS (Pallas, 1773)
И КАНАДСКОГО ЖУРАВЛЯ
GRUS CANADENSIS CANADENSIS (Linnaeus, 1758)
В РЕПРОДУКТИВНЫЙ ПЕРИОД ЖИЗНИ
НА СЕВЕРО-ВОСТОКЕ СИБИРИ
03.02.08 – экология
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Якутск – 2011
Работа выполнена в лаборатории зоологических исследований
Института биологических проблем криолитозоны СО РАН
Научный руководитель: доктор биологических наук
Официальные оппоненты:
доктор биологических наук
,
Якутский государственный
инженерно-технический институт
доктор биологических наук, профессор
Институт систематики и экологии животных
СО РАН
Ведущая организация: Бурятский государственный университет |
Защита состоится 27 октября 2011 года в 10 час. на заседании диссертационного совета Д 212.306.03 при ФГАОУ ВПО «Северо-Восточный федеральный университет им. », ГСП, Якутск, .
С диссертацией можно ознакомиться в научной библиотеке ФГАОУ ВПО «Северо-Восточный федеральный университет им. » по адресу: Якутск, .
E-mail: *****@***ru
Факс: (74112)335812
Автореферат разослан: «___» сентября 2011 г.
Ученый секретарь
диссертационного совета
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Стерх (белый журавль) является редким исчезающим видом, включенным в Красный список Международного Союза Охраны Птиц (МСОП); Красные книги птиц Азии и РФ; Красные книги ряда субъектов РФ, в том числе Республики Саха (Якутии). В Международной красной книге (IUCN) классифицируется как критически угрожаемый вид (Bird Life International 2000); является третьим по редкости в мире видом журавлей (Meine, Archibald, 1996). С целью сохранения восточной популяции стерха, Республиканский ресурсный резерват (РРР) «Кыталык» в 1997 г. включен в Международную сеть особо охраняемых местообитаний редких журавлей Северо-Востока Азии, созданной по инициативе Международного Фонда Охраны Журавлей (МФОЖ / International Crane Foundation) и Международного бюро по сохранению водно-болотных угодий (Wetlands International), а также Общества диких птиц Японии, при содействии Агенства окружающей среды Японии. Канадский журавль в Якутии представлен периферийной популяцией многочисленного подвида (малый канадский журавль), занесенного в региональную Красную книгу; в Красном списке МСОП малый канадский журавль относится к группе видов пониженного риска (Лабутин, Дегтярев, Ларионов, 1982; Лабутин, Дегтярев, 1988). Гнездовые ареалы стерха и канадского журавля, заселяющих северо-восточные субарктические тундры Якутии, перекрываются.
Несмотря на многолетние исследования, экология, питание и поведение стерха и канадского журавля в Якутии изучены далеко недостаточно. Взаимоотношение стерха и канадского журавля в гнездовой период на территории совместного обитания ранее практически не рассматривались. Ввиду особой редкости белого журавля, неустойчивости и ранимости его популяции, особое место для его сохранения приобретает изучение экологии вида в период гнездования, роста и развития птенцов.
Цель и задачи исследования.
- определить пространственное распределение стерха и канадского журавля на гнездовье в прииндигирской тундре;
- изучить основные характеристики и особенности их воспроизводства;
- исследовать кормодобывание, питание, их особенности в разные сезоны репродуктивного периода журавлей;
- оценить бюджет времени и выявить особенности поведения журавлей в репродуктивный период.
Научная новизна и практическая значимость. В процессе исследования пространственного распределения и численности стерха и канадского журавля на гнездовье в северо-восточных районах России впервые получены свидетельства гнездования канадского журавля в прииндигирской тундре и успешности его дальнейшего расселения на запад. Впервые исследован бюджет времени различных аспектов жизнедеятельности сравниваемых видов журавлей, связанных с обеспечением их собственного существования и воспроизводства. Получены новые материалы по характеристике структуры гнездового участка журавлей и особенностей его использования. Изучена социальная и возрастная структура их населения на модельных участках гнездового ареала. Установлены некоторые ранее неизвестные аспекты гнездостроения журавлей, поведения птиц на гнезде, особенностей их питания. Установлены особенности перемещения выводков в условиях субарктической тундры.
Результаты исследований имеют прикладное и фундаментальное значение при анализе адаптаций тундровых видов журавлей к условиям субарктической зоны. Изучение различных аспектов жизнедеятельности журавлей необходимо для разработки дальнейших мер их охраны, учитывающих новые потенциальные угрозы и специфические требования рассматриваемых видов к среде обитания. Полученные материалы могут быть полезны для научного обоснования расширения сети охраняемых водно-болотных угодий, имеющих ключевое значение для стерха и других мигрирующих околоводных птиц в Азии. Полученные материалы использованы при выполнении Проекта программы ООН по Охране Окружающей Среды и Глобального Экологического Фонда GF/7 «Cовершенствование сети охраняемых водно-болотных угодий, имеющих ключевое значение для стерха и других мигрирующих околоводных птиц в Азии» Программы ООН по Охране Окружающей Среды и Глобального Экологического Фонда и при составлении программы эколого-просветительского проекта МФОЖ «Три белых журавля, два пролетных пути, один мир» в Якутии.
Защищаемые положения:
1. Оптимальные местообитания в бассейне Индигирки стерха – это увлажненные приозерные низменности и межозерные понижения в системе крупных озер; канадского журавля- главным образом слабоувлажненные и сухие кочкарниковые возвышения.
2. Вытеснение в бассейне Индигирки канадского журавля стерхом на более высокие и сухие ярусы тундры не оказывает заметного негативного воздействия на популяцию этого вида благодаря его пластичности и толерантности.
3. Распределение по различным экологическим нишам стерха и канадского журавля в прииндигирской тундре позволяет им реализовывать имеющийся потенциал к росту.
Апробация работы: Материалы исследований были представлены на региональных конференциях (конференция научной молодежи «Эрэл-2005», 2005 г.; «Эрэл-2007», 2007 г., Якутск; научно-практическая конференция «Экологическая культура населения как фактор формирования имиджа Республики Саха (Якутия)», 2010 г., Якутск; региональная выставка-ярмарка инновационных идей и проектов молодых ученых и предпринимателей "Молодежь. Наука. Бизнес", 2009, Якутск); рабочих совещаниях проекта GF/7 «Совершенствование сети охраняемых водно-болотных угодий, имеющих ключевое значение для стерха и других мигрирующих околоводных птиц в Азии» Программы ООН по Охране Окружающей Среды и Глобального Экологического Фонда; Международной конференции по мигрирующим птицам севера Тихоокеанского региона (Якутск, 2007 г.); Международной Конференции «Проблемы популяционной экологии животных» (Томск, 2006 г.); III Международной Орнитологической конференции «Современные; «Проблемы орнитологии Сибири и Центральной Азии», Улан – Уде, 2006; IV Международная орнитологическая конференция «Современные проблемы орнитологии Сибири и Центральной Азии» (Улан-Уде, 2009 г.); Всероссийской научно-практической конференции с международным участием (Рязань, 2009 г.); Международной конференции «Россия и Германия: исторический опыт научного сотрудничества в Сибири и на Дальнем Востоке (к 300-летию , 2009 г., Якутск); North-East V European crane Conference, 2003; XX International Congress of Zoology, University Pierre et Marie Curie (University Paris-Sud and the Museum National dיּHistoire Naturelle, 2008); 46th Animal Meeting of the Society for Cryobiology (Hokkaido, University Sapporo, Japan. 2009).
Публикации: По теме диссертации опубликовано 34 работы, из них три в журналах списка ВАК.
Структура и объем диссертации. Работа состоит из введения, трех глав, заключения и выводов; изложена на 168 стр., содержит 24 таблицы, 33 рисунка. Список использованной литературы включает 159 публикаций (иностранных – 62).
ГЛАВА 1. ЭКОЛОГИЯ И СРЕДА ОБИТАНИЯ СТЕРХА И КАНАДСКОГО ЖУРАВЛЯ В ЯКУТИИ (ОБЗОР ЛИТЕРАТУРЫ)
Глава состоит из двух частей: 1. Систематическое положение и история изучения; 2. Эколого-географическая и ландшафтная характеристика основных районов исследований. В первой части главы рассмотрены вопросы систематики видов, история изучения и современное состояние изученности стерха и канадского журавля по материалам отечественной и мировой литературы. Во второй части подробно рассмотрены природно-климатические условия обитания журавлей на северо-востоке Сибири: рельеф, освещенность и температура, осадки, снежный покров и прочие атмосферные явления, гидрологическая сеть и гидрологический режим, растительность и животный мир.
ГЛАВА 2. МАТЕРИАЛ И МЕТОДИКИ ИССЛЕДОВАНИЙ
Основные исследования гнездовой жизни журавлей проводились в индигирском очаге повышенной плотности гнездования стерха (Дегтярев, Лабутин, 1991), на левобережье р. Индигирка (в границах территории Республиканского ресурсного резервата (РРР) «Кыталык»), и в нижнеколымской тундре (РРР «Чайгургино»), в гг. В РРР «Кыталык» (участок «Елонь) и на сопредельных с ним территориях, протяженность наземно-водных маршрутов составила 1290 км. На модельной территории площадью 1314 км2 в общей сложности зарегистрировано 103 гнездовых участка журавлей: стерх - 47, канадский журавль - 56, обнаружено 63 гнезда (соответственно, 47 и 16).
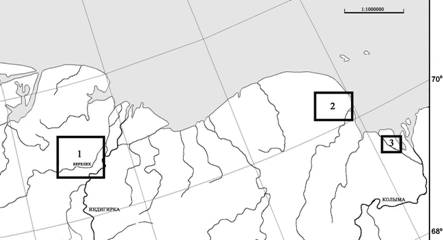
Рис. 1. Районы исследований в нижнеиндигирской (1) и нижнеколымской тундрах (2 – бассейн реки Большая Чукочья, 3 - местность «Походская Едома»)
На различных стадиях гнездования журавлей под хронометрируемыми наблюдениями различной продолжительности находилось 11 гнезд (стерх - 4, канадский журавль - 7), 13 пар (соответственно, 8 и 6) и 7 выводков (5 и 2). Общая продолжительность хронометрируемых наблюдений по установлению бюджета времени журавлей в период гнездования составила 143:30 час (стерх -118:09, канадский журавль – 25:21): во время гнездостроительства и насиживания кладки – 95:34 час (соответственно, 83:21 и 12:13), в выводковый период – 47:56 (34:48 и 13:08). В нижнеколымской тундре протяженность водно-пеших маршрутов в гг. составила 592 км. В бассейне нижнего течения р. Большая Чукочья на территории 400 км2 выявлена одна территориальная пара стерха, установлена структура населения канадского журавля, состоящая из 239 особей: 108 пар журавлей, 7 одиночных и 16 неполовозрелых особей в трех группировках из 5 (n=2) и 6 (n=1) птиц (0,59 особ./ км2), эффективность ее размножения: 38 выводков (35,2 %) с одним (n=17) и двумя (n=21) птенцами. Общая продолжительность наблюдений за поведением отдельных пар (n=16) и выводков (n=12) составила 108:34 часа.
В период собственных исследований ( гг.) был окольцован один взрослый канадский журавль (1999 гг.) и 10 птенцов стерха (2005 г. – 8, 2008 г. – 2). При этом на канадском журавле и 2 птенцах стерха (2008 г.) были закреплены спутниковые передатчики (PTT/ Platform Transmitter Terminal) американского производства. В 2008 г. РТТ-100 компании Microwave Telemetry (40 г) фиксировались, как и ранее на канадском журавле, на голени нелётных птенцов стерха специальным кольцом-крепежкой. Птиц отлавливали с помощью вертолета Ми-8 по методу, ранее разработанному и апробированному в Якутии на птенцах стерха (Дегтярев, Лабутин, 1991), кольцевание и их прижизненную обработку проводили по общепринятым методикам (Дольник, 1976; Виноградова, Дольник, Ефремов и др., 1976).
Особенности поведения, питания и демографической структуры стерха в период миграций были проанализированы на основе материалов, собранных как на территории Якутии, так и в северо-восточном Китае, при использовании Сети мониторинговых пунктов наблюдения за состоянием популяции и среды обитания вида на ключевых участках гнездования, пролета и транзитных остановок птиц в Северо-Восточной Азии.
ГЛАВА 3. ЭКОЛОГИЯ СТЕРХА И КАНАДСКОГО ЖУРАВЛЯ В РЕПРОДУКТИВНЫЙ ПЕРИОД ЖИЗНИ
Стерх является узкоспециализированным видом, для поддержания собственного жизнеобеспечения которого как на территории гнездования, так и в местах транзитных остановок и зимовок необходимы условия повышенной обводнённости, учитывающие определённый тип прибрежной и водной растительности (Гермогенов и др., 2002; Meine, Archibald, 1996; Watanabe, 2006). Канадский журавль, напротив, отличается некоторой степенью эврибионтности, и требования вида к гнездовому участку не ограничены жесткими рамками сочетания определённых условий (Владимирцева, Слепцов, 2006). О самых разнообразных типах местообитаний в отношении степени увлажненности при выборе места для гнезда говорит и анализ зарубежной и отечественной литературы (Кищинский, Флинт, Томкович и др., 1982; Винтер, 2002; Walkinshaw, 1965; Boise, 1976). Сравнительный анализ местоположений гнезд журавлей позволяет полагать, что канадский журавль в выборе места для гнезда руководствуется незанятостью местообитаний стерхом. Если в прииндигирской тундре, в условиях соседства со стерхом, оптимальные местообитания канадский журавль находит в умеренно-обводненном ярусе тундры, часто встречаясь на ее сухих возвышенных участках, то в нижнеколымской, где стерх малочислен, он предпочитает гнездиться и водить птенцов в наиболее низких, обводненных местообитаниях. Находки гнезд в обводненных местообитаниях, иногда на мокром субстрате, подтверждают возможность канадского журавля гнездиться в условиях приозерных и межозерных низменностей (Кондратьев, Кречмар, 1982; Кречмар, Андреев, Кондратьев, 1978, Кондратьев, 1988; Владимирцева, Бысыкатова, Слепцов, 2009).
3.1. Пространственное распределение и численность стерха и канадского журавля на гнездовье в прииндигирской тундре
Прямыми доказательствами наличия у стерхов филопатрии являются возврат окольцованных нелетных птенцов и взрослых птиц в район их гнездования. Косвенными доказательствами служит наличие многолетне используемых гнездовых построек, небольшая индивидуальная межгнездовая дистанция, регулярная встречаемость птиц или пар на одних и тех же участках. Окольцованные территориальные птицы возвращались на изначально избранные гнездовые участки, часто занимая одни те же гнездовые постройки.
На основании отлова и кольцевания стерхов, данных РТТ, находок гнезд и выводков, встреч птиц на местах гнездования и зимовки проведен мониторинг местопребывания пар стерха (n=47) на модельной территории (S=1314 км2), установлены их гнездовые участки на основной исследуемой территории в период с 1990 по 2007 гг. Приводимые материалы показывают наличие пространственной устойчивости гнездовых участков стерха и их регулярную занятость, вероятно, одними и теми же парами. Материалы аэро - и наземных исследований основных параметров гнездовых участков стерха подтверждаются данными космического мониторинга за предмиграционными перемещениями 9 взрослых стерхов с РТТ: 1995 г. – 1 и 1996 г. – 8 птиц (из 8 пар), по которым были получены наиболее полные сведения.
3.1.2. Динамика численности. В последние годы гнездящееся население стерха на модельной территории в целом выросло на четыре пары (таб. 1).
Таблица 1
Динамика численности стерха и канадского журавля на модельной территории прииндигирской тундры (S=1314 км2)
Год | Количество особей | Плотность населения, ос./10 км2 | ||
Стерх | Канадский журавль | Стерх | Канадский журавль | |
1996 | 86 | 3 | 0,65 | 0,06 |
2005 | 92 | 110 | 0,70 | 0,83 |
2009 | 94 | 112 | 0,71 | 0,85 |
Выводок канадского журавля из двух взрослых птиц и двух птенцов в прииндигирской тундре впервые зарегистрирован 24 июля 1998 г. на левобережье р. Берелех в 1,5-2 км от кордона «Елонь». 13 июля 1999 г. на этом участке тундры было обнаружено уже две пары с птенцами. В последующем гнездовые пары, находки гнезд и выводки канадского журавля стали чаще отмечать в окрестностях стационара «Джюкарское» на берегу одноименного озера и на всех лодочно-пеших маршрутах. С 1995 по 2009 г. плотность гнездовых пар выросла в 13,3 раз.
Рост численности на гнездовье канадского журавля подтверждается наблюдениями в приколымской тундре, где популяция приближается по этому показателю к таковым в оптимуме ареала вида на Чукотке и Аляске.
3.2. Гнездовой участок журавлей и его использование
Журавли, как и все моногамные виды птиц, обладают определенной гнездовой территорией. Птицы пребывали на гнездовых участках в течение всего периода нахождения в тундре, независимо от их участия в размножении. Предгнездовой период журавлей длится с момента их прилета в середине мая до конца мая – начала июня, когда птицы приступают к яйцекладке. В это время готовящиеся к размножению пары занимают и осваивают гнездовой участок.
3.2.1. Размещение, конфигурация и размеры гнездового участка. Все встреченные и территориально локализированные в прииндигирской тундре пары и выводки стерха, отловленные и окольцованные птицы, обнаруженные и исследованные гнезда находились в приозерных обводненных низменностях и межозерных заболоченных понижениях, не заливаемых во время весеннего половодья. Большинство пар стерха обладают участками площадью от 7,3 до 17 км2 (Гермогенов, Пшенников, Канаи, 2009). В течение всего периода пребывания в на летовке, независимо от наличия или отсутствия птенцов, птицы не покидают индивидуальных участков, а единично фиксируемые небольшие по расстоянию и кратковременные перемещения за пределы их границ отмечены лишь незадолго перед отлетом. Если в прииндигирской тундре, в условиях соседства со стерхом, канадский журавль занимает местообитания в умеренно-обводненном ярусе тундры, часто встречаясь на её возвышенных сухих участках, то в нижнеколымской тундре, где стерх малочислен, он предпочитает гнездиться и водить птенцов в наиболее низких, обводненных местообитаниях, свойственных стерху.
3.2.2. Структура гнездового участка. Данные визуальных наблюдений за территориальными парами стерха и спутниковой пеленгации местопребывания птиц дают основание для выделения в гнездовом участке двух зон -«гнездовой» и «кормовой». В гнездовой зоне устраивается гнездо; здесь находится кормовая база для птенца в его первые дни после выведения из яйца, и для постоянно опекающих в это время его родителей. Особенно заметна структуризация участка в период насиживания, когда птица, сменяемая партнером на гнезде, практически сразу улетает кормиться. Но птица не покидает гнездовую зону, если кормится животными объектами, в частности, рыбой, выходящей на затопленные территории вокруг озер после весенних паводков. Возможно, птицы, кормящиеся растительными объектами вдалеке от гнезда, сохраняют кормовые ресурсы на период первых дней после выведения птенцов, когда возникает необходимость оставаться возле гнезда. У канадского журавля столь чёткого разграничения гнездовой территории на кормовую и гнездовую зоны не проявляется.
3.2.3. Гнезда. При совпадении оптимальных природно-климатических факторов, период гнездования начинается в конце мая - начале июня. Насиживание кладки (28-30 дней у стерха, 29–32 дня у канадского журавля) представляет собой период повышенного стрессового состояния для птиц. Изменчивые и непредсказуемые в различные годы погодные условия, их сезонная неустойчивость могут значительно повлиять на процесс яйцекладки журавлей или, как следствие, негативно отразиться на эффективности их размножения. Совпадение сроков начала насиживания с периодом снеготаяния вызывает необходимость постоянного контроля за уровнем высоты гнезда в случае его подтопления талыми водами. Оставление птицами не насиженной кладки даже на короткий период грозит ее разграблением, главным образом, крупными чайками. Особенно опасными в отношении возможного разорения гнезд являются годы с депрессией численности мышевидных грызунов. В период гнездования на территории исследования в прииндигирской тундре обнаружено 47 гнезд стерха и 16 гнезд канадского журавля.
3.2.4. Месторасположение гнезд. Большая часть обнаруженных гнезд стерха находилась на увлажненных и обводненных участках, занятых преимущественно осоковыми, в межозерных и приозерных низменностях (n=43). Большинство гнезд стерха располагались на расстоянии до 0,5 км (n=21) и 0,5-1 км (n=24) до озера, и лишь два гнезда располагались в 1,5 и 2 км от ближайшего озера. Гнезда канадского журавля располагались на расстоянии 0,4-1,1 км (0,75±0,35 км) до ближайшего среднего или крупного озера. Большая часть обнаруженных гнезд находились в пределах полигональной тундры, реже гнезда находились на низких обводненных или, напротив, на сухих приподнятых участках тундры. Факты обнаружения гнезд (n=2) в значительно обводненных низменностях, расположенных на высоте 16 и 18 м над уровнем моря, что типично для стерха, не позволяют однозначно утверждать, что канадский журавль предпочитает гнездиться на более сухих и приподнятых участках тундры, как принято считать. Расстояние между гнездами стерха и канадского журавля значительно меньше (1,81±0,53 км), чем между гнездами внутри вида, как у стерха (6,79±3,75), так и канадского журавля (3,05±0,61).
3.2.5. Техника (способы) строительства и строительный материал гнезда. Результаты наблюдений за гнездостроительным поведением журавлей показали, что стерх и канадский журавль способны достраивать гнезда в ходе насиживания кладки в случае их размножения в условиях повышенной увлажненности гнездовых участков. Это наблюдается при совпадении сроков начала размножения и интенсивного снеготаяния, особенно в годы с обильными зимними осадками. Журавли практически не достраивают гнезда лишь в случае их расположения в относительно сухих местообитаниях. Прошлогоднее гнездо может быть повторно использовано, если находится в зоне, успевшей освободиться от снега на период начала яйцекладки.
Методы сбора и транспортировки строительного материала к гнезду у трех пар стерха и одной пары канадского журавля, наблюдаемых в 2004 и 2005 гг. на ранней и средней фазах насиживания, хотя и имели один принцип, различались в деталях. Все наблюдаемые пары стерха собирали осоковые растения, в основном, пушицу. Выстилки гнезд канадского журавля состояли из гидрофильной травянистой растительности, лишайников, обломанных веточек карликовых березы и ивы, т. е. использовался материал в непосредственной близости от гнезда.
У обоих видов журавлей сбор строительного материала производился не насиживающим кладку партнером, и состоял из двух этапов: вначале пучки растений выдергивались из почвы и отбрасывались назад (в направлении гнезда) во время перемещения птицы от гнезда. Затем журавль проходил этот же маршрут снова, перебрасывая уже вырванные из почвы на первом этапе пучки растительности ближе к гнезду. Положение корпуса было всегда направлено вперед, лишь шея и голова при броске отводились вбок. Стерхи на каждом шагу меняли направление броска назад пучка растительности, благодаря чему движения птицы напоминали движения косаря. Птица, находящаяся в гнезде, дотягивалась клювом к подкидываемым партнером пучкам, и укладывала их в гнездо. Иногда не насиживающий журавль помогал сидящему партнеру в укладке материала в гнездо.
Различие действий у двух видов журавлей состояло в том, что стерхи при сборе материала двигались всегда строго по прямой линии длиной до 10 м от гнезда, а канадские журавли - по спирали, центр которой представляло гнездо. Такая техника конвейерной транспортировки строительного материала для гнезда представляется наименее энергетически затратной и экономящей время.
3.3. Особенности (характеристика) выводкового периода журавлей
Как указывалось выше, сроки начала откладки яиц зависят от хода весны: первое яйцо может появиться как рано - в последней декаде мая, так и заметно позже, в конце первой-начале второй декады июня.
3.3.1. Переворачивание яиц в гнезде. Во время наблюдения за насиживающими парами стерха было установлено, что в среднем яйца в гнезде переворачивались, в целях равномерного обогревания кладки (Мальчевский, 1959), через 68 мин. Смена партнеров на гнезде у наблюдаемых пар стерха чаще сопровождается переворачиванием яиц, которое производит заменяющий насиживающего партнер, что экономит время вставшей с гнезда птицы. Сменяющий партнер не переворачивает яйца, если они были уже перевернуты незадолго до его возвращения к гнезду насиживающим партнером. Большая часть всех переворачиваний яиц совершается обогревающей кладку птицей (26 из 38 зарегистрированных). В 33,3 % от общего числа отмеченных переворачиваний это происходило при смене партнеров на гнезде (в 73,3 % случаев смен).
3.3.2.Смена партнеров на гнезде . У пар стерха, наблюдаемых в 2005 г., смена партнеров на гнезде не представляла собой сложного ритуала с унисональным дуэтом, как это описывается в литературе (Рощевский, 1973). У пар канадского журавля, напротив, каждая смена на гнезде сопровождалась вокализацией, что оказалось почти единственным вспомогательным фактором при поиске их гнезд. Обнаружено, что на ранних сроках насиживания стерхи сменяли друг друга через более короткие периоды, чем пары, наблюдаемые в конце периода насиживания (рис. 2).
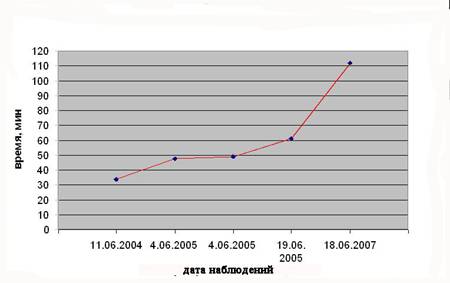
Рис. 2. Изменения продолжительности времени между сменами партнеров у четырех наблюдаемых пар стерха с увеличением сроков насиживания
Все 28-30 дней насиживания кладки являются периодом повышенного стрессового состояния для журавлей; возможно, поэтому в конце этого срока для восполнения энергетических затрат птицам необходимо более длительное время кормления. По данным наблюдений за тремя парами стерхов, партнеры сменяли друг друга на гнезде через периоды от 41 мин до 2,38 час. В 2004 г. у пары № 7 смена партнеров на гнезде в завершающей фазе насиживания в дневные часы 14 июня происходила примерно через равные промежутки времени – в среднем 34 мин. Самец и самка у обоих видов журавлей в равной степени распределяли обязанности насиживания. Такое равноценное распределение может иметь значение в случае гибели одного из партнеров. Так, в 2000 г. одно из обнаруженных гнезд канадского журавля, расположенного восточнее оз. Круглое, с единственным яйцом, оказалось опекаемым лишь одной птицей. Вероятно, ее партнер погиб после того, как гнездо было построено и отложена кладка. Птица, пол которой не было возможности установить, успешно вывела птенца. В итоге, наблюдения за сменами партнеров на гнезде позволили установить, что, вопреки некоторым литературным источникам, самка и самец в парах стерха поровну распределяют обязанности по насиживанию кладки, сменяя друг друга через определенное время (88,35+54,68 мин), как и канадские журавли.
3.3.3. Величина выводка. На территории исследования все встреченные выводки стерха (n=14, гг.) состояли из одного птенца. Количество яиц в кладке стерха не устанавливалось в целях ограничения беспокойства птиц. Известно, что из двух-яйцовой, в большинстве случаев, кладки стерха, в подавляющем большинстве случаев остается лишь один птенец. Высокая степень птенцовой агрессии, проявляемая видом в условиях неволи (Флинт, 1987; Сидоренко, 2002); сведения персонала центра разведения журавлей Международного Фонда Охраны Журавлей, США, Висконсин), как основная причина этого факта, не находит достаточной основы для подтверждения благодаря обнаружению двух парных выводков из 19 встреченных 11-27 августа в 1990 и 1991 гг. в Хромо - Береляхской тундре (Дегтярев, Лабутин, 1999). Проявление птенцовой агрессии у птенцов канадского журавля отмечено лишь во время кормления выводка (Reed, 1998). В устье р. Большая Чукочья 87% всех встреченных выводков с двумя птенцами родители распределяли обязанности – каждая из взрослых птиц водила одного птенца. Разведение птенцов, вероятно, имеет определенную цель – избежание птенцовой агрессии во время кормления.
Анализ литературных и собственных данных по канадскому журавлю показывает преимущественное количество кладок, состоящих из двух яиц (1,89±1,1) и выводков из двух птенцов (1, 71±1,11). Таким образом, несмотря на довольно частое присутствие второго яйца в кладках стерха (величина кладки 1,59±0,90), выводки в подавляющем большинстве случаев состоят из одного птенца (величина выводка 1,03±0,28). У канадского журавля выживаемость птенцов гораздо выше: величина кладки составляет 1,89±1,1, выводка - 1, 71±1,11.
3.3.4. Рост и развитие птенцов. Промеры птенцов восточной популяции стерха были взяты во время работ по кольцеванию и установке спутниковых передатчиков на северо-востоке Якутии. Оказалось, что масса диких птенцов стерха несколько превышает вес птенцов, выращенных в неволе, что соответствует мнению об ускоренных темпах роста и развития в субарктических условиях (Постельных, Кашенцева, 2005; Potapov, 1992; Watanabe, 2006). Как выяснилось, значительные различия в росте и развитии птенцов могут присутствовать внутри одной и той же популяции в границах одного сезона. Птенцы одной из 38 выводковых пар, наблюдаемых в устье р. Большая Чукочья 13 и 14.07, были крупнее (вполовину размера взрослой птицы), чем птенцы большинства пар (треть размера взрослой птицы). Кроме того, очень крупным, более половины размера взрослой птицы, был единственный птенец пары, встреченной в бассейне р. Эмэскхэвэм (приток р. Большая Чукочья). Возможно, некоторые пары, достигшие территории гнездования во время оттепели, в период 9-25 мая, сразу приступили к насиживанию. Птенцы пар, которые смогли сохранить кладку в период заморозков и периодически случавшейся пурги, длящийся 26 мая – 8 июня, имели значительный разрыв в возрасте с птенцами журавлей, имевших возможность приступить к гнездованию лишь по окончанию неблагоприятного периода. Более старший возраст подтверждается более самостоятельным поведением крупных птенцов, наблюдаемых большую часть времени кормящимися без помощи родителей, на достаточно удалённом от них расстоянии, иногда до 50 м. Следовательно, более быстрое развитие журавлят в субарктической зоне, как увеличение размеров, так и приобретение навыков к полету являются адаптацией, необходимой в условиях с возможными долгосрочными запаздываниями начала гнездования.
3.3.5.Перемещения выводка на территории гнездования. Сильная привязанность пар стерха с птенцами к собственным гнездовым участкам подтверждается их поведением во время работ по кольцеванию и установке спутниковых передатчиков. Пары стерхов с птенцом передвигаются при кормлении в основном в размеренном темпе, пересекая по нескольку раз небольшой участок за сутки, обычно не превышающий 500 м2. Выводки канадского журавля оказались гораздо более мобильными, особенно в бассейне Индигирки. Здесь канадский журавль находится в условиях более низкой плотности популяции в периферийной зоне ареала размножения и разделения территории гнездования со стерхом. В результате постоянных перемещений выводка наблюдения оказываются краткосрочными. Наблюдаемые здесь выводки (n=2, 1998 и 2000 гг) скрывались за короткий период наблюдения за пределы досягаемой видимости, проходя значительные дистанции (Lim max 8 км/ 11:03 ч, 1998 г.). Ситуация масштабных перемещений канадских журавлей с птенцами, сопровождающихся постоянным кормлением, наиболее схожа с описанием : «выводки начинают широко кочевать по тундре» после того, как покидают гнездо (Флинт, 1987). Однако в оптимуме численности гнездового ареала субарктической Северной Америки и Чукотки перемещения выводков ограничены собственными участками. Несколько менее обширные перемещения, но гораздо большая склонность к ним, чем у пар стерха, наблюдалась у выводков в устье р. Большая Чукочья, территории с крайне низко выраженным антропогенным фактором и отсутствием стерха на гнездовье. Пары канадских журавлей, несмотря на наличие птенцов, передвигаются при кормлении в ускоренном темпе, перемещаясь за день до 1,5 км, не придерживаясь определенного участка. Очевидно, что выводок не придерживался ограниченного собственного охраняемого участка, как это происходит у стерха. Пары, как с птенцами, так и без птенцов, свободно перемещались по обширной территории, могли встречаться, соединяться на незначительное время в группы до восьми особей. Ни разу не наблюдалось каких-либо проявлений агрессии между парами или отдельными птицами. Вероятно, в случае недостатка кормовых ресурсов, внутривидовая агрессия должна была бы проявляться в той или иной степени. Возможно, кормовые ресурсы здесь, напротив, достаточно обильны для вида, ввиду более низкой численности периферийной популяции и отсутствия стерха на гнездовье, что снижает проявления территориальности здесь, по крайней мере, в выводковый период.
3.4. Кормодобывание и питание
Если канадский журавль известен как активный потребитель корма животного происхождения, то стерх в литературе описывается как наиболее склонный к вегетарианскому питанию журавль. Наблюдения за кормодобыванием стерха в ареале его гнездования и на остановках во время осенних и весенних миграций не показали значительного предпочтения видом растительных кормов. Добыча мелкой рыбы и привлечение на приваду стерхов в 2005 г. может объясняться периодом труднодоступности растительного корма до схода снежного покрова и протайки верхнего почвенно-растительного слоя. Но в 2007 г., когда в середине июня растительность уже была доступна, стерхи гнездовой пары в период насиживания тратили большую часть времени кормления (90%) на высматривание, ловлю и поедание мелкой и крупной рыбы трёх видов (таб. 3).
Таблица 3
Видовой состав и количество рыбы, отлавливаемой
гнездовой парой cтерхов за сутки, 24 ч
Вид рыбы | Количество добытой рыбы, % | Длина рыбы, Lim см | |||
2007 г. | 2008 г. | ||||
% | n | % | n | ||
Девятииглая колюшка Pungitius pungitius L | 78,7 | 91 | 69,0 | 48 | 3-5 |
Щука Esox lucius L. | 6,0 | 7 | 11,9 | 8 | 20-30 |
Пелядь Coregonus peled Gm. | 15,5 | 18 | 19,0 | 13 | 20-50 |
Наблюдения показали, что корм животного происхождения использовался и при кормлении птенца – при изобилии мелкой рыбы растительный корм был полностью исключен из потребления за время наблюдений в 3,5 часа.
Целенаправленный поиск и активное поедание сибирских углозубов на весенней транзитной остановке также указывает на выбор калорийной пищи животного происхождения при ее нахождении в тех обводненных условиях, к которым привязан этот узкоспециализированный вид журавля, по крайней мере, в весенне-летнее время. В определенные периоды года, такие, как весеннее половодье, в результате которого вокруг крупных озер образуются неглубокие затопления с выходящей в них озерной рыбой в ареале гнездования стерха, или сезон икрометания у сибирских углозубов в середине мая на территориях транзитных остановок, стации обитания стерхов пополняются энергоемким видом корма. Зазубрины на дистальных частях подклювья и надклювья стерха должны обеспечивать захват собственно скользких подводных кормовых объектов, как животного, так и растительного происхождения, что естественно для вида, привязанного к наиболее низким и обводненным стациям.
Принципиальных различий между канадским журавлём в устье р. Большая Чукочьея и в бассейне р. Индигирка в характере питания и кормовом поведении не обнаружено, однако большую часть времени кормления (68%) журавли в устье р. Большая Чукочья, проводили на наиболее низких и обводненных уровнях рельефа, выкапывая корневища осоковых. В низовьях Индигирки, напротив, канадских журавлей наиболее часто наблюдают кормящимися на умеренно-обводненных участках и на поднятиях рельефа (Watanabe, 2006). Очевидно, что обводненные зоны не исключаются канадским журавлем для кормления, и в бассейне Индигирки вид вытесняется стерхом в другую экологическую нишу на более высоких ярусах тундры, что не оказывает значительного воздействия благодаря пластичности и толерантности канадского журавля.
Более протяженные перемещения как пар без птенцов, так и выводков канадского журавля при кормлении в гнездовом ареале могут объясняться несколькими причинами: низкой территориальностью вида в выводковый период; толерантностью к местообитаниям в отличие от привязанного к обводненным местообитаниям стерха; и более поверхностным сбором корма (включающего наземные части растений, насекомых, мелких млекопитающих и птенцов мелких птиц), что требует охвата больших площадей.
Стерх и канадский журавль могут проявлять конкурентные взаимоотношения в середине мая – начале июня до схода снежного покрова при недостатке единственно полноценного корма в это время – мышевидных грызунов. Очень незначительное количество растительного корма в это время представляют остатки ягод с прошлого сезона, прошлогодние съедобные мерзлые части растений и проростки растений на участках, освободившихся от снега.
В результате наблюдения за кормовым поведением журавлей в период насиживания кладки показали преимущественное (90%) питание стерха животным, а не растительным кормом, - молодью и взрослой рыбой, вопреки нескольким литературным источникам. По литературным и собственным сведениям установлено 16 животных кормовых объектов стерха и канадского журавля в гнездовом ареале.
3.5. Бюджет времени журавлей в выводковый период.
Отличительной особенностью канадского журавля является постоянное перемещение, сопряженное с кормлением. За весь период времени наблюдения в 1998 г. канадские журавли бассейна Индигирки предоставили возможность отдохнуть своим птенцам лишь два раза, примерно по полчаса, в то время как птенец стерха отдыхает каждые два часа. Птенцам канадских журавлей в устье Чукочьей также предоставлялся более продолжительный отдых - до 70 мин (n=4), у некоторых выводковых пар до 4 раз в день, во время которого одна (n=1) или две взрослые птицы стояли рядом с птенцами или на расстоянии до 50 м (n=1) от них. Канадские журавли выводковой пары, наблюдаемые в бассейне Индигирки, в очаге наиболее высокого воспроизводства стерха, находились в состоянии настороженности в 14 раз больше по времени, чем журавли устья р. Большая Чукочья, и почти в 6 раз дольше находились в позах тревоги, что может объяснять крайне незначительное антропогенного беспокойство и отсутствие стерха на гнездовье в устье Большая Чукочья.
Стерхи, имеющие птенца, по крайней мере, в течение суток перемещаются в диапазоне не более 1 км. Пар, имеющих птенца, можно было идентифицировать по неторопливому передвижению, с остановками, во время которых птица поджидает птенца или партнера, и возвращениями к партнеру или птенцу. Выводок передвигается при кормлении, совершая маятникообразные переходы назад и вперед, от 100 до 300 м. Птенец большую часть времени проводит рядом с матерью; самка чаще кормит птенца. Кормление птенца родителями занимает 12,9% времени суток. В течение 47 мин непрерывного наблюдения за одной из пар, родители покормили птенца 11 раз, из них 4 раза самец, 7- самка. За 28 мин наблюдения птенец находился на расстоянии 1-3 м от самки в течение 15 мин (53,6 % времени наблюдения), 4 мин. в нейтральной зоне между самкой и самцом (14,3%) и 9 мин. (32,1%) – на расстоянии более чем 15 м от обоих родителей, но ближе к самке, чем к самцу. Все птенцы канадских журавлей, наблюдаемые в устье р. Большая Чукочьей, кормились как самостоятельно, так и при помощи родителей, но более мелкие птенцы чаще (примерно на 10%) подбегали к родителям за кормом. Самостоятельно птенцы могут добывать насекомых и молодые побеги на верхушках осоковых. Родители кормят птенцов выкапываемыми корневищами осоковых и фрагментами тела леммингов и полёвок. Птенцы сами подбегают к родителям и склёвывают корм у них с клюва. Насекомых птенцы схватывают самостоятельно.
3.6 Поведение журавлей в местах размножения
3.6.1.Территориальное поведение. Наблюдения взаимоотношений журавлей субарктической тундры показало, что территориальное поведение в наибольшей степени проявляется между парами отдельными особями стерха. Значительно реже регистрируются социальные конфликты между стерхом и канадским журавлём (рис. 3). Преследование территориальными парами групп молодых птиц из трех-шести особей у канадского журавля наблюдалось лишь в период насиживания кладки, 16-30 июня 2011 г. в дельте Колымы, участке «Походская Едома». В периоды строительства гнезда и насиживания, наиболее потенциальные для территориальных столкновений, большинство канадских журавлей, встречаемых в бассейне Индигирки, находились в парах или отдельными особями, т. е. провоцирующие ситуации были исключены из наблюдения. Атака в полете одиночного канадского журавля стерхом, чей партнер находился на гнезде, наблюдалась один раз. Примеры нейтрального и даже положительного отношения стерха к канадскому журавлю более многочисленны. Так, в 2000 г. наблюдалось совместное кормление стерха и канадского журавля из соседних территориальных пар, в то время пока их партнеры насиживали каждый свою кладку.
Отсутствие проявления внутривидовой территориальности у канадских журавлей, наблюдаемых в выводковый период 12-23 июля 2010 г., в устье р. Большая Чукочья может находить объяснение в достаточном количестве здесь энергоёмких кормовых ресурсов, расположенных в наиболее низких и обводненных участках тундры, значительно занятых в прииндигирской тундре стерхом.
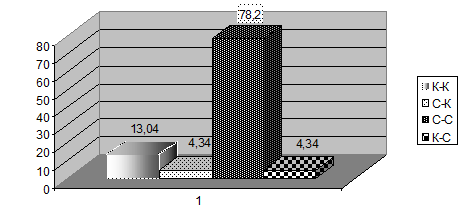 |
Рис. 3. Соотношение межвидовых и внутривидовых конфликтов (n=18) за период исследований на территории гнездования: С-С – конфликты между особями стерха; С-К – атака стерхом канадского журавля; К-С – атака канадским журавлем стерха; К-К –конфликты между особями канадского журавля
3.6.2. Защита гнезда и гнездового участка. Особенно опасными в отношении возможного разорения гнезд являются года с депрессией размножения мышевидных - леммингов и полевок. За время исследований ( гг.) отмечено два случая отпугивания стерхом одиночного северного оленя от гнезда внезапным принятием вертикального положения с резким раскрытием крыльев; три случая атаки стерха одиночных особей серебристой чайки, бургомистра и тундрового лебедя; и 5 атак песца. Следовательно, перечисленные виды выдают себя как несущие угрозу кладкам стерха. Потенциальными грабителями кладки стерха и малого канадского журавля могут быть заходящие в тундру росомаха, волк, медведь; собаки, сопровождающие стада кочующего через его местообитания домашнего оленя, а также человек. Кладки и выводки из пуховых птенцов вспугнутых и оставивших их без присмотра журавлей могут быть разорены также крупными чайками и поморниками, бургомистрами. В июле 1998 г. наблюдался самец канадского журавля, защищавший свой выводок от щенков песца низким перелетом с последующим ударом ногами. Зарегистрирован случай разорения песцом гнезда канадского журавля. Дистанция вспугивания человеком стерха с гнезда в ночное время суток составляет 900±200м (n=4), днем – до 1,5±0,5км (n=6); канадского журавля –60±40м (n=12). Привыкнув к присутствию человека, стерхи могут подлетать к объекту опасности и идти навстречу, приближаясь до 200 м., принимая позы угрозы. Дистанция вспугивания малого канадского журавля гораздо меньше: птицы приближаются к человеку, сразу не опознав его, на 50-100 м. При обнаружении одного из гнезд малого канадского журавля 3 июня 2005 г. сидящая на гнезде птица подпустила человека на 10 м.
3.6.3.Социальная структура гнездящихся журавлей. Основу населения стерха и канадского журавля в гнездовой период составляют пары (80-90%), но регулярно встречаются одиночные особи и небольшие группы не участвующих в размножении птиц. Соотношение выделяемых социальных субъединиц довольно существенно меняется по годам.
3.6.4. Периоды стайного образа жизни журавлей. Стайный образ жизни журавлей в период миграций и зимовок обеспечивает создание социальных связей и репродуктивных пар. Установлено, что в парах стерха замена погибшего партнера может произойти достаточно быстро, за период менее года. В двух из трех случаях замены в парах стерха выжившая особь обрела нового партнера уже, очевидно, во время первой же зимовки после потери прежнего.
Молодые птицы обоих рассматриваемых видов, по всей вероятности, приобретают независимость уже на первом году жизни, и до обретения партнера держатся небольшими группами в 3-7 птиц, или парами, время от времени отмечаемыми на территории гнездования.
ВЫВОДЫ
1. Различные экологические ниши стерха и канадского журавля в прииндигирской тундре нивелируют конкурентные взаимоотношения между видами и позволяют им реализовывать имеющийся потенциал к росту популяций.
2. Стерх и канадский журавль добавляют растительный материал в гнездо с уже насиживаемой кладкой при повышении увлажненности гнездовых участков в период снеготаяния способом, выстроенным в плане максимального сохранения времени и энергии. Прошлогоднее гнездо может быть использовано, если находится в зоне, успевшей освободиться от снега на период начала насиживания кладки.
3. В период насиживания пары стерха не используют растительные кормовые ресурсы в непосредственной близости от гнезда - в «гнездовой» зоне участка, сохраняя корм на период первых дней после появления птенца. До появления птенца стерхи употребляют растительные корма в отдаленных от гнезда «кормовых» зонах; вблизи гнезда птицы питаются преимущественно животными кормами. У канадского журавля подобного зонирования гнездового участка не выявлено.
4. Наблюдения за насиживанием кладки яиц показало, что самец и самка стерха и канадского журавля равное количество времени уделяют этому процессу. Такое равноценное распределение обязанностей имеет особое значение в случае гибели одного из партнеров. Для стерха определено, что продолжительность насиживания кладки каждым партнером между сменами увеличивается к началу выклева птенца.
5. Изучение питания в период насиживания журавлей показало преимущественное (90%) питание стерха рыбой, вопреки большинству литературных источников, сообщающих о предпочтении растительных кормов этим видом.
6. Преимущественное кормление канадского журавля на низких и обводненных уровнях рельефа в нижнеколымской тундре показывает, что эти зоны не исключаются им для кормления, и в бассейне р. Индигирка он вытесняется стерхом на более высокие ярусы тундры. Вытеснение канадского журавля в эту экологическую нишу не оказывает на него значительного воздействия благодаря пластичности и толерантности вида.
7. Масштабные перемещения выводков канадского журавля в прииндигирской тундре могут объясняться, во-первых, толерантностью вида к местообитаниям, в отличие от привязанного к обводненным местообитаниям стерха; во-вторых, поверхностным сбором корма, доступного на территориях более возвышенного рельефа, что требует охвата больших площадей.
Список основных публикаций
В журналах списка ВАК
1. , Основные этологические аспекты стерха (GRUS LEUCOGERANUS) и малого канадского журавля (GRUS CANADENSIS CANADENSIS) в период насиживания кладки / , // Зоол. ж., 2009. - Т. 88.- Вып. 2. - С. 221-227
2. , , Особенности использования гнездовой территории канадским журавлем в Якутии / , , // Сиб. экол. журн., 2009. –- Т. 16. Вып. 3. – С. 423–427.
3. , , Весенние миграции стерха в Якутии / , , . // Сиб. экол. журн, 2010. –Вып. 1. - С. 121-125
В иностранных изданиях
4. Germogenov N. I., Solomonov N. G., Pshennikov A. E., Degtyarev A. G., Sleptsov S. M., Yegorov N. N., Vladimirtseva M. V., Bysykatova I. P., Okoneshnikov V. V. Eastern population of Siberian Crane: structure, habitats, nesting and migration. Integrative Zoology / N. I. Germogenov, N. G. Solomonov, A. E. Pshennikov, A. G. Degtyarev, S. M. Sleptsov, N. N. Yegorov, M. V. Vladimirtseva, I. sykatova, V. V. Okoneshnikov // X International Congress of Zoology - 2008. - Р 7.
5. Germogenov N. I, Degtyarev A. G., Labutin Yu. V., Solomonov N. G., Pshennikov A. E., Kanai Yu., Sleptsov S. M., Egorov N. N., Bysykatova I. P., Vladimirtseva M. V., Archibald G., Harris Jim, Nagendran M., Chan S., Veta M. Banding and monitoring of Siberian Cranes in Yakutia / N. I. Germogenov, .G. Degtyarev, Yu. V Labutin., N. G. Solomonov, A. E. Pshennikov, Yu. Kanai, S. M. Sleptsov, N. N. Egorov, I. sykatova, M. V. Vladimirtseva, G. Archibald, J. Harris, M. Nagendran, S. Chan, M. Veta // China Crane News. China Crane and Waterbird Specialist Group. - Felburn, 2007. - Vol. 11. - No 1. - P. 26-35.
6. Important Bird Areas in Asia: Key Sites for Conservation. Editors: S. Chan, M. J. Crosby, M. Z. Islam and A. W. Tordoff. Birdlife Conservation Series // Important Birds Areas in Asia. 2004. В списке исполнителей от Якутии: N. Germogenov, A. Degtyarev, M. Vladimirtseva
7. Solomonov N. G., Germogenov N. I., Isayev А.P., Nakhodkin N.А., Degtyarev V. G., Yegorov N. N., Sleptsov S.М., Оkoneshnikov V. V., Vladimirtseva М.V., Bysykatova I. P. Rare and endangered species of birds of eastern Yakutia taiga and tundra regions / N. G. Solomonov, N. I. Germogenov, А. P. Isayev, N. А.Nakhodkin, V. G. Degtyarev, N. N.Yegorov, S. М. Sleptsov, V. V. Оkoneshnikov, М. V. Vladimirtseva, I. sykatova // 46th Animal Meeting of the Society for Cryobiology.- Japan. Hokkaido, 2009. - P. 241.
8. Germogenov N., Pshennikov A., Vladimirtseva M. Present distribution and state of crane populations of Yakutia North-East / N. Germogenov, A. Pshennikov, M. Vladimirtseva // V European crane ConferenceP. 7.
В сборниках
9. , , 2011. Кормодобывание и питание стерха и канадского журавля в период гнездования и миграций / , , // Тр. ин-та систематики и экологии животных СО РАН. 2011. - Вып. 47. - С.
11. Материалы по поведению и бюджету времени стерха и канадского журавля в Якутии / // Журавли Евразии (распределение, численность, биология). Журавли Евразии (распределение, численность, биология). М.: Россельхозакадемия, 2002. – С. 234-239.
12. , Наблюдения за гнездостроительной деятельностью стерха / , // Инф. бюлл. РГЖЕ. –М., 2005. –Вып. 9 - С.29-30.
13. , , Кормодобывание стерха во время миграций и гнездования / , ., , // III Межд. конф. по мигрирующим птицам севера тихоокеанского региона. – Якутск, 2007. С. 28-30.
14. , Определенная степень толерантности канадского журавля к местообитаниям для гнездования на северо-востоке Якутии / , // Cб. III Международной Орнитологической конференции «Современные проблемы орнитологии Сибири и Центральной Азии». - Улан – Уде, 2006. – Ч. 2. - С. 48-51.
15. , Описание метода строительства нового гнезда парой стерхов на северо-востоке Якутии / , // Орнитологические исследования в Северной Евразии.- Ставрополь, 2006. — С. 116-117.
16. , К поведению стерха Grus leucogeranus в период насиживания кладки / , // Сб. тр. науч. молодежной конф. «ЭРЭЛ-2005». –Якутск, 2005. - С. 112-116
17. , , К вопросу территориальных взаимоотношений стерха и канадского журавля / , , // Сб. тр. науч. молодежной конф. «ЭРЭЛ-2007». –Якутск, 2008. - С. 176-179
18. , Специфика межвидовых взаимоотношений журавлей северо-востока Якутии / , // III Межд. конф. по мигрирующим птицам Севера Тихоокеанского региона. - Якутск: Изд-во ЯНЦ СО РАН, 2007. - С. 26-28
19. , И. Миграции стерхов в долине р.
Индигирки, Якутия / , Е. И Ильяшенко // Инф. бюлл РГЖЕ. – М., 2007.- Вып. 10. - С. 29-30.
20. , Наблюдения за осенним пролетом стерха в 2008 г. на территории и в окрестностях села Охотский Перевоз на юго-востоке Якутии / , // Материалы Всероссийской научно-практической конференции с межд. участием «Экология, эволюция и систематика животных». – Рязань, 2009. - С. 192-193.
Подписано в печать 23.09.2011. Формат 60х 84/16.
Бумага тип. №2. Гарнитура «Таймс». Печать офсетная.
Печ. л. 1,5. Уч.-изд. л. 1,87. Тираж 100 экз. Заказ.
Издательско-полиграфический комплекс
Северо-Восточного федерального университета
8.
 |
Отпечатано в типографии ИПК СВФУ
