

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
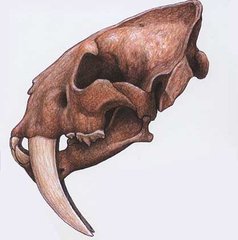


Смилодон.
Смилодон. Эта форма занимает следующее место вслед за Ужасным волком по количеству особей, найденных в асфальтовых лужах и сильно превосходит все другие виды кошек. Миллер (1968) насчитал 2100 особей смилодонов из Ранчо Ла Бреа (коллекция Page Museum), основываясь на черепах и их фрагментах. Смилодон приближался в размере к африканскому льву, хотя его тело и конечности имели несколько иные пропорции. У смилодона задние конечности были относительно легкими, в то время как передние конечности были сильными и мощными (рис. 18). Крепость и сила обнаруживаются также в ребрах и грудине. Нижние части конечностей (локтевая кость, большая берцовая) относительно короче в сравнении с аналогичными костями пещерных львов.
Смилодон имел пропорции конечностей, которые были наиболее схожи с пропорциями конечностей ягуара (если сравнивать с современными крупными кошками). Оба этих вида разделяли характерные черты задних конечностей с стопоходящими хищниками. Кажется более вероятным, что смилодон использовал засаду, нежели быструю погоню за добычей (Gonyea, 1976b). Любопытной чертой этого животного является короткий хвост. В этом смилодон демонстрирует внешнее сходство с рысью. Фундаментальные отличия между смилодоном с одной стороны и львом и пумой с другой стороны наиболее рельефно проявляется в черепе и расположении зубов (рис. 19). Долго думали, что смилодон обладал относительно маленьким мозгом, хотя Джерисон (Jerison, 1973) показал, что размер мозга смилодона только чуть-чуть отличается от размера мозга крупных кошек. Череп сильно изменен в плане приспособления к сильно развитым кинжалообразным зубам верхней челюсти. О некоторых из этих изменений стоит упомянуть. Носовое отверстие несколько отступало от его типичного расположения в черепах кошек. Твердое небо обнаруживает выпуклый костистый гребень, который идет по всей длине этой поверхности.
Череп смилодона демонстрирует отличительные признаки в росте mastoid, представляющего большую площадку для прикрепления мускулов, сильно давящих вниз голову. Элементы нижней челюсти, в противоположность нижним челюстям современных кошек, слабее. Судя по развитию структур, к которым крепились важные мускулы, нижняя челюсть отклонялась на широкий угол, когда рот открывался для атаки и сила укуса могла быть несколько ослаблена. Зубной ряд смилодона указывает на несколько необычную специализацию. У этой кошки зубной ряд взрослых особей обычно насчитывал 26 зубов, в то время как лев или пума имеют 30 зубов. Другими словами, в сравнении с современными кошками саблезуб потерял передние премоляры на каждой стороне верхней и нижней челюсти. Верхние зубы являлись большими кинжалообразными, значительно удлиненными и сплющенными в поперечнике. Передний и боковой край были слегка зазубрены. Название «смилодон» буквально означающее «зубы –сабли» очень хорошо подходит к этому животному. Нижние клыки уменьшились в размере и походили на нижние резцы. Коренные зубы были сдавлены в поперечнике и хорошо приспособлены для режущих действий. Атакуя крупных животных, таких как мамонты, мастодонты или земляные ленивцы смилодон, вероятно, должен был искать уязвимые места своих жертв, хватать жертву мощными передними лапами и быстро наносить удар верхними клыками, оставляя рваные раны. При такой атаке нижняя челюсть была способна отводиться вниз, позволяя значительно шире раскрывать рот. Мощная голова и шейная мускулатура обеспечивали сильный удар. Низкое положение nasal, вероятно, давало возможность животному дышать тогда, когда голова была глубоко погружена в бок жертвы. Сильно рифленые десны, покрывающие края твердого неба могли служить для стока крови, как полагал Сток, но более вероятным представляется укрепление черепа, необходимое для противодействия давлению, которому череп подвергался во время борьбы. Акерстен считал, что смилодон использовал клыки, вонзая их в живот крупных жертв, аргументируя это тем, что  относительно хрупкая природа длинных и тонких клыков не могла быть эффективно использована в области тела, где кости появлялись почти сразу после кожи. Недавнее сравнение износа зубов смилодона с образцами 8 других существующих хищников наводит на мысль, что смилодон редко ел кости, но иногда участвовал в поедании падали ( Van Valkenburgh et al., 1991). Любопытной чертой, отмеченной при исследовании большой коллекции скелетных останков смилодонов, является высокая частота повреждений отдельных элементов. В целом, патологические нарушения иногда отмечаются в черепах и зубах. Они особенно очевидны в костях конечностей и в поясничной области. Кроме того, помимо переломов, которые зажили в течение жизни, в ряде костей животных и птиц из асфальтов были найдены аномалии, преобладающие среди саблезубых кошек. Согласно доктору R. L. Moodie, который большое внимание уделил изучению болезней, поражавших в Ла Бреа животных и птиц, наибольшие патологии были отмечены для ран с последующими инфекциями. Были выявлены случаи вывихов и артритов. Не удивительно найти нарушения нормального роста костей у таких свирепых созданий как смилодон и ужасный волк. Раны, несомненно, часто наносились в стычках, которые происходили, когда большое число этих животных собиралось у асфальтовых луж. Менее очевидна причина, которая способствовала аномалиям в позвоночном столбе у смилодонов. В поясничной области (особенно 2-й или 4-й позвонки) позвонки могли соединяться, образуя более или менее сплошную трубу, благодаря развитию излишней костной ткани вдоль стороны и основания позвоночных соединений (рис. 20). Эта болезнь похожа на патологии у человека. Так или иначе, специфические повадки этих созданий были в определенной мере ответственны за это необычное слияние, которое не было предопределено, хотя состояние некоторых останков такое, что позволяет легко отличить эту группу хищников от других животных. Патологические раны, которые случались наиболее часто у смилодонов в области шеи, грудной клетки и поясничной области согласуются с травмами, полученными во время охоты и борьбы с жертвами на земле. Септические повреждения согласуются с укусами или другими открытыми ранами. Выживание особей (достаточно долгое время после получения раны, делающей животное неспособным охотиться) было интерпретировано социальным поведением этого вида (Heald 1989, Shaw 1991). Особенно это заметно на черепах смилодонов, у которых один или оба клыка были сломаны при жизни. Эти раны, видимо, причиняли боль во время еды. У таких образцов сломанные края клыков демонстрируют стертые, ровные поверхности, ясно указывая на факт, что кошка долго использовала зубы после получения раны. У животных со сломанными при жизни клыками коренные зубы также демонстрируют чрезмерную изношенность, позволяя предположить, что животное находило большую необходимость в этих зубах после потери клыков.
относительно хрупкая природа длинных и тонких клыков не могла быть эффективно использована в области тела, где кости появлялись почти сразу после кожи. Недавнее сравнение износа зубов смилодона с образцами 8 других существующих хищников наводит на мысль, что смилодон редко ел кости, но иногда участвовал в поедании падали ( Van Valkenburgh et al., 1991). Любопытной чертой, отмеченной при исследовании большой коллекции скелетных останков смилодонов, является высокая частота повреждений отдельных элементов. В целом, патологические нарушения иногда отмечаются в черепах и зубах. Они особенно очевидны в костях конечностей и в поясничной области. Кроме того, помимо переломов, которые зажили в течение жизни, в ряде костей животных и птиц из асфальтов были найдены аномалии, преобладающие среди саблезубых кошек. Согласно доктору R. L. Moodie, который большое внимание уделил изучению болезней, поражавших в Ла Бреа животных и птиц, наибольшие патологии были отмечены для ран с последующими инфекциями. Были выявлены случаи вывихов и артритов. Не удивительно найти нарушения нормального роста костей у таких свирепых созданий как смилодон и ужасный волк. Раны, несомненно, часто наносились в стычках, которые происходили, когда большое число этих животных собиралось у асфальтовых луж. Менее очевидна причина, которая способствовала аномалиям в позвоночном столбе у смилодонов. В поясничной области (особенно 2-й или 4-й позвонки) позвонки могли соединяться, образуя более или менее сплошную трубу, благодаря развитию излишней костной ткани вдоль стороны и основания позвоночных соединений (рис. 20). Эта болезнь похожа на патологии у человека. Так или иначе, специфические повадки этих созданий были в определенной мере ответственны за это необычное слияние, которое не было предопределено, хотя состояние некоторых останков такое, что позволяет легко отличить эту группу хищников от других животных. Патологические раны, которые случались наиболее часто у смилодонов в области шеи, грудной клетки и поясничной области согласуются с травмами, полученными во время охоты и борьбы с жертвами на земле. Септические повреждения согласуются с укусами или другими открытыми ранами. Выживание особей (достаточно долгое время после получения раны, делающей животное неспособным охотиться) было интерпретировано социальным поведением этого вида (Heald 1989, Shaw 1991). Особенно это заметно на черепах смилодонов, у которых один или оба клыка были сломаны при жизни. Эти раны, видимо, причиняли боль во время еды. У таких образцов сломанные края клыков демонстрируют стертые, ровные поверхности, ясно указывая на факт, что кошка долго использовала зубы после получения раны. У животных со сломанными при жизни клыками коренные зубы также демонстрируют чрезмерную изношенность, позволяя предположить, что животное находило большую необходимость в этих зубах после потери клыков.
 Очевидно, что такие особенности были помехой в борьбе за существование. Однако интересные находки Ван Валкенбурга показывают, что несмотря на относительную хрупкость клыки смилодона ломались менее часто, чем клыки живущих сейчас хищников. Извлеченные полные и полу целые скелеты смилодонов из отложений, обнаруженных во время строительства Page Museum указывают на то, что метаподии и фаланги стали постепенно уменьшаться от 2-го к 5-му пальцу, в отличие от большинства прошлых реконструкций, основанных на неполных или несвязанных скелетных материалах. Все онтогенетические стадии смилодона, за исключением очень молодых особей, были обнаружены в Ла Бреа. Отсутствие незрелых форм может указывать на то, что район логова не был локализован вблизи асфальтовых луж. Саблезубые кошки имели длинную и богатую историю в Северной Америке (от раннего олигоцена до позднего плейстоцена). Саблезубые кошки, тесно связанные с образцами из Ла Бреа, обнаружены во Флориде, Небраске, Мехико, Техасе, а схожие формы известны из плейстоцена Южной Америки и Западной Европы. Берта (1985) свела число видов смилодонов к 2 – Smilodon gracilis (от позднего плиоцена до среднего плейстоцена) и Smilodon populator (включая S. californicus, S. floridanus, S. fatalis от среднего до позднего плейстоцена). Но ее интерпретация пока еще не всеми принята. Мы здесь следуем таксономии Чарчера (Churcher), объяснявшего происхождение североамериканских и южноамериканских видов. Подвид S. fatalis brevipes был предложен Мэрриамом и Стоком (1932) за необычно короткий machairodontid метаподии из Ла Бреа. Или этот таксон представляет особый подвид смилодона или это вторгнувшийся S. populator (характеризовавшийся укороченными дистальными сегментами конечностей), вторжение которого уже было доказано.
Очевидно, что такие особенности были помехой в борьбе за существование. Однако интересные находки Ван Валкенбурга показывают, что несмотря на относительную хрупкость клыки смилодона ломались менее часто, чем клыки живущих сейчас хищников. Извлеченные полные и полу целые скелеты смилодонов из отложений, обнаруженных во время строительства Page Museum указывают на то, что метаподии и фаланги стали постепенно уменьшаться от 2-го к 5-му пальцу, в отличие от большинства прошлых реконструкций, основанных на неполных или несвязанных скелетных материалах. Все онтогенетические стадии смилодона, за исключением очень молодых особей, были обнаружены в Ла Бреа. Отсутствие незрелых форм может указывать на то, что район логова не был локализован вблизи асфальтовых луж. Саблезубые кошки имели длинную и богатую историю в Северной Америке (от раннего олигоцена до позднего плейстоцена). Саблезубые кошки, тесно связанные с образцами из Ла Бреа, обнаружены во Флориде, Небраске, Мехико, Техасе, а схожие формы известны из плейстоцена Южной Америки и Западной Европы. Берта (1985) свела число видов смилодонов к 2 – Smilodon gracilis (от позднего плиоцена до среднего плейстоцена) и Smilodon populator (включая S. californicus, S. floridanus, S. fatalis от среднего до позднего плейстоцена). Но ее интерпретация пока еще не всеми принята. Мы здесь следуем таксономии Чарчера (Churcher), объяснявшего происхождение североамериканских и южноамериканских видов. Подвид S. fatalis brevipes был предложен Мэрриамом и Стоком (1932) за необычно короткий machairodontid метаподии из Ла Бреа. Или этот таксон представляет особый подвид смилодона или это вторгнувшийся S. populator (характеризовавшийся укороченными дистальными сегментами конечностей), вторжение которого уже было доказано.
Об обнаружении останков саблезубой кошки в Южной Америке впервые было объявлено Ландом (Lund) в 1842 г. Это сообщение было основано на анализе останков позднеплейстоценовой - голоценовой фауны на востоке Бразилии в Лагоа Санта (Lagoa Santa). Принадлежность саблезубой кошки смилодону была установлена по отдельным зубам и связанным друг с другом метаподиям. В начале, Ланд (1839 г.) ошибочно отнес некоторые из этих материалов к гиене (Hyaena neogaea). Впоследствии, другие плейстоценовые саблезубые кошки были обнаружены в Аргентине, Боливии, Эквадоре, Уругвае и Перу. Большинство из этих находок были определены как останки смилодонов, хотя некоторые были описаны как останки, принадлежащие новому роду или подроду (Smilodon (Prosmilodon) Rusconi, 1929; Smilodontidation Kraglievich, 1948). Мендес Алзола (Mendez Alzola) в 1941 г. отнес аргентинский материал к Smilodon bonaeriensis. Паула Коуто (Paula Couto) в 1955 г. предложил различать южноамериканский подвид S. populator populator и североамериканский подвид S. populator californicus. В 1967 г Чёрчер (Churcher), изучив некоторые аргентинские материалы и присоединив дополнительную информацию о некоторых других южноамериканских образцах, отнес эти находки к Smilodon neogaeus. Хоффстеттер (Hoffstetter) в 1952 г. дополнил наши знания о смилодоне находками из Эквадора, а Чёрчер описал превосходный экземпляр из Перу (в Талара). В настоящее время, выделяют два вида смилодонов. Маленький, с легким скелетом S. gracilis, жил в позднем плиоцене ― среднем плейстоцене (поздний Бланкан ― поздний Ирвингтониан) в Северной Америке. Крупный, с массивным, тяжелым скелетом, и более часто встречающийся, S. populator (его синонимы: S. floridanus Leidy, 1889; S. californicus Bovard, 1907; S. fatalis sensu Slaughter, 1963) жил в среднем и позднем плейстоцене (поздний Ирвингтониан ― Ранчолабреа) в Северной и Южной Америках.
Географическое распространение.
А. Берта указывает, что смилодоны были найдены во многих странах Южной Америки: в Бразилии (Лагоа Санта), Аргентине (Буэнос-Айресе, Мар дель Плата и его окрестности), Боливии (Тариха, Нуапуа), Эквадоре (Ла Каролина, Пунин), Уругвае и Перу (Талара). К списку стран, который приводит А. Берта, необходимо добавить еще и Венесуэлу. Там останки смилодонов были найдены после 1985 г.
Возраст находок. Самые древние останки смилодона были найдены в Аргентине в районе Мар дель Плата. Их датировали поздним плиоценом. Однако, согласно Д. Скаглиа (G. Scaglia), этот материал был собран не на том месте, которое датируется Чападмалалием (поздний плиоцен), но скорее в отложениях у основания скалы, которые более соответствуют Энсенадию (среднему плейстоцену). В Мар дель Плата и Мирамар (Аргентина) были найдены останки смилодонов, датированные Укием (ранний плейстоцен). Скаглиа, однако, поставил под сомнение эту датировку, и интерпретировал материал как относящийся к отложениям Энсенадия. Если оставить в стороне эти спорные датировки, то придется признать, что хорошо документированные следы присутствия смилодона относятся к Энсенадию ― Луханию (средний ― поздний плейстоцен). Останки смилодонов, найденные в Боливии (в Тариха, Нуапуа) датируются средним ― поздним плейстоценом. В данном случае определение возраста находок опирается на коллекцию фауны (вероятно, руководящие формы млекопитающих, позволяющие достаточно точно датировать возраст сомнительных находок). К позднему плейстоцену относятся находки из Перу, Эквадора и Уругвая. Весьма многочисленны позднеплейстоценовые находки в Аргентине. Исчезновение смилодона в Южной Америке приходится на поздний плейстоцен ― голоцен. Именно к этому периоду относятся находки из Бразилии (Лагоа Санта), имеющие самые поздние датировки.
Некоторые морфологические особенности
Череп очень крупный. Лицевая часть короткая и широкая. Сагиттальный гребень рельефно поднимается за слегка выпуклой передней частью. Затылочная часть характеризуется высокой степенью индивидуальной изменчивости. Верхние резцы крупные, острые и отогнуты вперед. Верхние клыки длинные, узкие, с большой кривизной и зазубренные. Клыки южноамериканских смилодонов длиннее, чем клыки североамериканских смилодонов из Ранчо Ла Бреа (Паула Коуто сравнил клык из Бразилии длиной 277,5 мм с клыками из Ранчо Ла Бреа (200 ― 262 мм)). Р3 сравнительно маленький и весьма изменчив по форме. У верхних хищнических зубов метаконид всегда длиннее, чем параконид. Протоконид обычно сильно редуцирован или отсутствует. Протоконид редко встречается и у североамериканских экземпляров из Ранчо Ла Бреа, на что обратили внимание еще Мэрриам и Сток (А. Берта в своем исследовании подтверждает выводы американских ученых). Экземпляры с неизношенными зубами демонстрируют наличие зазубрин вдоль режущей кромки. М1 сильно редуцирован. Диастема между клыком и Р4 сравнительно короткая у большинства южноамериканских образцов. Её длина варьируется и, возможно, это связанно с возрастом животных. Нижние резцы широкие, отогнутые назад. Нижние клыки имеют зазубрины с 2-х краев. Р3 обычно присутствует у экземпляров датируемых энсенадием и в плане морфологии варьируется очень сильно (коэффициент вариации = 23,4). Потеря Р3 чаще встречается у экземпляров датируемых луханием и ранчолабреа (Мэрриам и сток отмечали, что этот зуб присутствует лишь у 6 % экземпляров из Ранчо Ла Бреа). Коронки Р4 и М1 отклонены назад. У М1 параконид короче, чем протоконид (обе кромки имеют зазубрины).
Дискуссии по вопросам систематики
Как уже говорилось, название Smilodon было установлено Ландом в 1842 г., описавшим вид S. populator из пещер Лагоа Санта в Бразилии. Муньиз (Muñiz) в 1845 г. описал скелет саблезубой кошки из Аргентины под именем Munifelis bonaerensis. Однако это родовое имя недействительно, т. к. оно было опубликовано Муньизом в газете и потому не удовлетворяет международным требованиям, предъявляемым к публикациям. Позже, некоторые авторы некорректно именовали саблезубых кошек под именами Smilodon bonaerensis (Амегино (Ameghino), 1907; Рускони (Rusconi), 1931; Мендес Алзола, 1941). Амегино в 1888 г. описал вид Smilodon ensenadensis (= Machaerodus ensenadensis), основываясь на верхних клыках, найденных в отложениях энсенадия в Аргентине. Рускони в 1929 г. записал маленькую саблезубую кошку из энсенадия Аргентины в подрод Smilodon (Prosmilodon). Краглиевич (L. Kraglievich) в 1934 г., следуя за Рускони, использовал название Prosmilodon. В 1947 г. он же определил существование Smilodon (Prosmilodon) на основе маленького размера и наличия Р3 ( еще раз подчеркивает вариабильность и редкое наличие этого зуба, который, как она считает, может использоваться для отличия более ранних смилодонов из энсенадия и ирвингтониана от более поздних смилодонов лухания и ранчолабреа). Находка из Боливии, первоначально описанная Амегино (1902) как Machaerodus ensenadensis была отнесена Краглиевичем к Smilodon (Prosmilodon) ensenadensis ferox. Этот подвид был выведен на основе своего маленького размера, отсутствию зазубрин на верхних клыках и наличию Р3. Но, по мнению А. Берты, исследование этого материала показывает, что верхний клык был сильно изношен вдоль передней и задней кромки, что и стало причиной исчезновения зазубрин, которые, по ее мнению, безусловно, были. Второй подвид Smilodon (Prosmilodon) ensenadensis minor был выделен Краглиевичем в 1948 г. опять же на основе маленького размера и массивного посткраниального скелета. Однако, дополнительное исследование показало, что этот скелет находится в пределах нижней границы размеров смилодона. А. Берта рассматривает находки, отнесенные Краглиевичем к Smilodon (Prosmilodon), как принадлежащие Smilodon populator.
Smilodon crucians был описан Амегино (1904) на основании его маленького размера, длинной диастемы и узкой ветви нижней челюсти. А. Берта (ссылаясь на Мэрриама и Стока) замечает, что все это может быть отнесено к онтогенетическим вариациям, и указывать на то, что Smilodon crucians вполне мог быть молодой особью Smilodon populator. Как и ожидалось, высокий коэффициент вариации для длины диастемы среди смилодонов Северной и Южной Америки существенно снижает возможности использования данного признака в качестве таксономической характеристики. Исследование Smilodon crucians показало наличие зазубрин на нижнем клыке и щечных зубах, которые поддерживают предложение об отождествлении этого вида с Smilodon populator.
Новый род саблезубых ― Smilodontidation ― был предложен Краглиевичем в 1948 г. Основанием для этого стали кости конечностей, найденные в Аргентине. В целом, этот род был выделен на основании маленького размера костей и небольших отличий в морфологии. По мнению Берты, эти особенности вполне можно рассматривать как результат индивидуальной и возрастной изменчивости. Анализ эпифизов бедренной и большой берцовой костей позволил считать эти кости принадлежащими взрослому, но молодому животному. Берта согласна с Чёрчем, что Smilodontidation должен быть отождествлен с Smilodon populator.
Самое старое название смилодона в Северной Америке ― Felis (Trucifelis) fatalis ― было предложено Джозефом Лейди в 1868 г. Позже Лейди описал фрагмент черепа с отсутствующими зубами как новый вид европейского рода Machairodus ― M. floridanus. Во всех поздних работах он был отнесен уже к смилодону.
Бовард (Bovard) в 1907 г. предложил название Smilodon californicus, отличая этот вид от S milodon floridanus на основании того, что у последнего короче лицевая часть черепа и имеются особенности в форме неба. Позже Мэрриам и Стокк, более тщательно изучая материалы из Ранчо Ла Бреа (1932), к числу отличий между двумя видами добавили и отличия в мастоидной области.
Б. Куртэн (1965), изучив флоридских саблезубых, пришел к выводу, что S. floridanus и S. fatalis являются синонимами. Вебб (Webb) в 1974 г. при оценке дополнительного флоридского материала пришел к выводу о синонимичности Smilodon californicus и S. floridanus, используя в качестве доказательств результаты промеров черепов, нижних челюстей и зубов (результаты сильно перекрывали друг друга). Куртэн и Андерсон (1980), исследуя происхождение Smilodon fatalis, пришли к выводу, что многочисленные виды смилодонов (за исключением S. gracilis) не более чем подвиды. Чёрчер (1984), вслед за Слотером (Slaughter), отнес к S. fatalis следующие виды: Smilodontopsis conardi Brown, 1908; Smilodontopsis troglodytes Brown, 1908; Smilodon nebraskensis Matthew, 1918; Smilodon trinitensis Slaughter, 1960.
Род Smilodontopsis, предложенный Брауном в 1908 г., основывался на находках из Арканзаса. Отличием этого рода от Smilodon стали особенности плечевой кости. Два вида (Smilodontopsis troglodytes и Smilodontopsis conardi) были описаны на основе различий в морфологии верхних хищнических зубов.
Smilodon nebraskensis, как вид, основывался на находке нижней челюсти в Небраске. Его отличием от Smilodon californicus стала «решительно короткая диастема» (Matthew, 1918). А. Берта подчеркивает сомнительность таких оценок и подходов в вопросах систематики. Smilodon trinitensis был выделен Слотером в 1960 г. на основании находки нижней челюсти в Далласе (Техас). Она отличалась от челюстей ранее описанных видов крупными щечными зубами (относительно размеров самой челюсти) и отсутствием Р3.
Таксономическая путаница вокруг североамериканского смилодона спровоцировала дискуссию, результатом которой стала критическая оценка S. floridanus и S. fatalis (Slaughter, 1963). Как показал Слотер в своем исследовании, все характерные черты S. floridanus также свойственны и южноамериканским смилодонам (Smilodon populator). Среди большинства наиболее часто используемых характеристик можно назвать следующие: крупный череп с широкой лицевой частью, большая кривизна верхних клыков, зазубрины на верхних клыках и щечных зубах, сильно редуцированный протоконид у Р4, сильно редуцированный выступ нижней челюсти. Именно эти черты позволяют отличить Smilodon populator от Smilodon gracilis для которого как раз характерны небольшая кривизна верхних клыков, хорошо развитый протоконид у Р4, меньший размер черепа и посткраниального скелета, очень маленькие зазубрины на клыках и щечных зубах и т. д.
Smilodon populator
Южноамериканский S. populator всегда привлекал внимание своими большими размерами. В работе Пера Христиансена и Джона Харриса (Body Size of Smilodon (Mammalia: Felidae) // Journal of Morphology 2005, № 000.), посвященной определению массы смилодона, подчеркивается, что по массе популятор может быть сопоставим только с гигантскими плейстоценовыми львами Северной Америки. Масса S. populator была определена ими в 220-360 кг. При этом, ученые подчеркивают, что хотя в их работе и присутствовали крупные кости (все исследование построено на тщательном анализе плечевой, локтевой, бедренной и большой берцовой костей смилодонов и сопоставлении их с соответствующими значениями существующих крупных кошек), но они сознательно не включили самые крупные экземпляры. Однако, касаясь вопроса о максимально возможной массе смилодона популятора, они пишут, что особо крупные особи могли весить значительно более 400 кг.
Сравнение некоторых промеров костей популятора с уссурийским тигром массой 221 кг (масса была определена сразу после смерти животного).
Тигр:
Плечевая кость: длина ― 372,5 мм, наименьшая окружность диафиза ― 104, переднезадний диаметр диафиза ― 36,4 мм
Локтевая кость: длина ― 390,5 мм, переднезадний диаметр диафиза ― 37,7 мм
Бедренная кость: длина ― 429,5 мм, наименьшая окружность диафиза ― 102,5 мм, переднезадний диаметр диафиза ― 31,2 мм
Большая берцовая кость: длина ― 365,5 мм, наименьшая окружность диафиза ― 93 мм, переднезадний диаметр диафиза ― 27,5 мм
S. populator:
Плечевая кость(4экземпляра):
1. Длина ― 387,5 мм, наименьшая окружность диафиза ― 155 мм, переднезадний диаметр диафиза ― 54 мм.
2. Длина ― 333,5, наименьшая окружность диафиза ― 138,5 мм, переднезадний диаметр диафиза ― 49,6 мм.
3. Длина ― 374,5мм, наименьшая окружность диафиза ― 149 мм, переднезадний диаметр диафиза ― 51,5 мм.
4. *Длина ― 369,5 мм, наименьшая окружность диафиза ― 145 мм, переднезадний диаметр диафиза ― 51,4 мм.
Локтевая кость (2 экземпляра):
1. Длина ― 350,8 мм, переднезадний диаметр диафиза ― 49,9 мм.
2. *Длина ― 348,5 мм, переднезадний диаметр диафиза ― 36,3 мм.
Бедренная кость (2 экземпляра):
1. Длина ― 390,2 мм, наименьшая окружность диафиза ― 134,5 мм, переднезадний диаметр диафиза ― 40,4 мм
2. *Длина ― ?, наименьшая окружность диафиза ― 120 мм, переднезадний диаметр диафиза ― 35,8 мм
Большая берцовая кость:
1. *Длина ― 286 мм, наименьшая окружность диафиза ― 108 мм, переднезадний диаметр диафиза ― 32,7 мм
* означает, что эти кости принадлежат одному животному.
Как уже говорилось, Христиансен и Харрис не использовали в своей работе самые крупные экземпляры. Так, Берта указывает, что длина плечевой кости популяторов из позднего плейстоцена Южной Америки доходила до 395 мм, длина бедренной кости ― до 40 мм, длина большой берцовой ― до 294,2 мм. Впрочем, не факт, что и это есть верхняя граница, т. к. здесь учтены только целые кости, тогда как некоторые сломанные кости могут указывать на еще большие размеры.
Некоторые другие находки костей популяторов впечатляют ничуть не меньше
Строение в целом: Смилодон из Ла Бреа достигал размеров современного африканского льва, но имел очень мощные передние конечности, соразмерно крупную голову и короткий хвост. Передние конечности с их крупными, втягивающимися когтями должны были очень хорошо подходить для того, чтобы валить и обездвиживать крупных животных. Структура и пропорции конечностей и ступни, а также структура позвоночника указывают на то, что смилодон не принадлежал к животным долго преследующим добычу, но, вероятно, был способен на короткую стремительную атаку. Большинство современных авторов согласны, что смилодон, вероятно, близко подкрадывался к своим потенциальным жертвам, а затем стремительно преодолевал короткую дистанцию. Я полагаю, что массивные передние конечности употреблялись для захвата верхней части тела жертвы сбоку и сбивания ее на землю для дальнейшей атаки зубами (таким образом, чтобы брюшная полость оставалась незащищенной). Этот метод сваливания жертвы схож с методом, который часто используется многими крупными живущими кошками, когда они атакуют крупную жертву, за исключением того, что последние могут также использовать свои зубы во время этой атаки.
Область атаки: Мартин (1980) рассматривал атаку саблезубых кошек, особенно кинжалозубых форм подобных смилодону, в брюхо как очень мало вероятную. Он аргументировал это тем, что жертва не могла быть тот час же убита и, пока не наступил шок, постаралась бы убежать от атакующего, который не был приспособлен преследовать убегающих животных. Он также заявил, что брюхо может быть защищено головой жертвы, и что удар по широкому, покатому брюху едва ли был бы лучшим. Я согласен с последним пунктом; однако, зубы смилодона отлично приспособлены для атаки в брюшную полость. Кроме того, трудно представить себе как формы, которые были потенциальными жертвами смилодона в Ранчо Ла Бреа, могли использовать свои головы для защиты своего живота, кроме как растянувшись на земле. На рисунке 10 смилодон валит молодого мамонта и вонзает зубы в его живот. – p. 18.
Верхние зубы: Морфология клыков смилодона была описана Мэриамом и Стоком (1932). Краткий обзор: смилодон имел длинные, узкие, загнутые назад саблеобразные зубы с крайне тонкой внешней эмалью (фигура 4). Эмаль простиралась до линии десны вдоль заднего края, но ее толщина отличалась от толщины эмали остальных зубов в среднем на 2/3. Эта очень тонкая эмаль простиралась от кончика до десен на заднем крае и от кончика до середины или 2/3 длины по переднему краю. Задний край более сжатый, чем передний, особенно у основания, выступающей наружу части. Здесь передний край становился полностью круглым, приближаясь к окончанию резцов. Зубы часто стирались, вплоть до гладкой поверхности (у старых особей). Уже это указывает на то, что зубы не использовались просто для украшения. Я полагаю, что острота зубов и очень тонкая эмаль, покрывающая клыки смилодона, демонстрируют относительно нечастое использование клыков (использовались во время убийства, но не регулярно во время еды). Более частое использование должно было потребовать увеличения толщины эмали и более сильного развития зубов. Крупные корни верхних зубов, до 40 % их общей длины, несомненно, отражают необходимость поддержки во время напряженного использования.
Виды жертв: Вообще, предполагалось, что смилодон и другие саблезубые были приспособлены для атаки крупных и толстокожих животных, таких как земляной ленивец или хоботные. Непосредственные данные, имеющиеся в распоряжении, идут только из позднеплейстоценовой фауны пещеры Friesenhahn в Техасе. Эта пещера была логовом гомотерия. Множество молодых хоботных, преимущественно мамонтов, было найдено здесь, но взрослые особи являлись редкостью. Эта очевидная связь логова и останков молодых хоботных предполагает, что гомотерий охотился, преимущественно, на молодых хоботных и приносил их останки в свое логово. Возможно, грубо зазубренные грани сабель гомотерия и другие зубы способствовали расчленению скелета для более легкого получения мяса. Эти резцы были особенно акулоподобными – с очень зазубренными краями. Если стадное и родительское поведение вымерших хоботных было схоже с поведением современных африканских слонов, охота на молодых особей могла быть опасным занятием. Многие, если не все, стада слонов защищают и помогают молодым; они могут даже продолжать защищать тело несколько дней, после смерти животного. Даже, если поведение мамонтов было подобно поведению индийских слонов с отдельным выкармливанием и защитой не стадом, а одной особью, как предположил Грэхам, атакуя хищники должны были быть осторожными. В любом случае, гомотерии выжидали идеальную возможность и ретировались немедленно, даже после успешной атаки на молодых (до оставления места охоты взрослыми хоботными). Конечно, соблазнительно продолжить эту интерпретацию на смилодона и других саблезубых, но имеются два существенных отличия в морфологии, хотя бы отличия порядка следования зубов. В дополнение, укус смилодона мог быть равным образом эффективен и для более мелких тонкокожих жертв. Мощное строение смилодонов указывает, что они, вероятно, могли успешно атаковать любое крупное травоядное в Ла Бреа, за исключением взрослых хоботных. Их, безусловно, привлекали попавшиеся в ловушку нехоботные и поэтому, они так обычны во многих отложениях Ла Бреа, в которых в целом ощущается недостаток останков хоботных. Я сомневаюсь, что смилодон ел исключительно членов какого-либо одного вида – среди живущих крупных хищников таких нет, - но они могли чаще охотиться на молодых хоботных, чем это делали другие хищные, найденные в Ла Бреа.
