


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Гемагглютинация, вызываемая вирусами, не является иммунологической реакцией, так как здесь нет системы антиген – антитело. С помощью этой реакции легко выявить присутствие гемагглютинирующего вируса, но невозможно его идентифицировать. Вирусы, обладающие гемагглютинирующими свойствами, имеют на поверхности гемагглютинины, с помощью которых происходит склеивание эритроцитов. По своей химической природе гемагглютинины являются глико - или липопротеидами.
В процессе взаимодействия вирусов с эритроцитами различают стадии адсорбции, агглютинации и элюции. Адсорбция вирусов на эритроцитах начинается как неспецифическая, а затем переходит в специфическую вследствие сродства гемагглютининов к рецепторам эритроцитов. На одном эритроците адсорбируется множество вирусов, они образуют мостики между эритроцитами, изменяется электростатический заряд эритроцитов и в результате наступает следующая стадия взаимодействия – гемагглютинация, на которой процесс может остановиться. Некоторые вирусы способны к элюции – освобождению с поверхности эритроцитов. Элюция происходит под действием вирусных ферментов (например, нейраминидазы) на рецепторы эритроцитов. Элюированные вирусы при этом не повреждаются, эритроциты же не способны повторно адсорбировать вирус вследствие разрушения рецепторов.
При постановке РГА используют те эритроциты (птиц, животных, человека), с которыми гемагглютинирующие свойства данного вируса проявляются лучше (например, эритроциты кур). Задерживающее действие на РГА могут оказывать ингибиторы, содержащиеся в тканях, где размножался вирус, и других биологических субстратах. Для снятия ингибиторного действия вируссодержащие материалы прогревают, обрабатывают трипсином, ацетоном и др.
2.2.1. Техника постановки РГА
На плексигласовой доске готовят двукратные возрастающие разведения в объёме 0,5 мл исследуемого вируссодержащего материала, начиная с 1:10 и до 1:1280. в луночки с разведениями добавляют равные объёмы 1% взвеси эритроцитов кур. Доску осторожно встряхивают и оставляют на 30-45 мин при комнатной температуре. Учёт результатов проводят по трёхкрестовой системе в зависимости от внешнего вида осадка эритроцитов. При гемагглютинации на «+++» осадок имеет форму зонтика и занимает всё дно лунки. При отсутствии агглютинации эритроциты оседают в центре лунки в виде компактного диска.
Титром вируса называется то наибольшее его разведение, при котором ещё наблюдается выраженная гемагглютинация («+++» или «++»). В нашем примере титр вируса 1:320 (табл. 4). Количественное содержание вируса в разведении, равном титру, называется гемагглютинирующей единицей (содержится в 0,5 мл разведения 1:320). Определение этой величины необходимо для постановки РТГА, в которой в качестве рабочей дозы вируса берут 4 гемагглютинирующие единицы.
Таблица 4
Схема постановки реакции гемагглютинации при диагностике гриппа
Ингре- диенты РГА (в мл) | Номера лунок разведения вируссодержащего материала | ||||||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
1:10 | 1:20 | 1:40 | 1:80 | 1:160 | 1:320 | 1:640 | 1:1280 | К | |
Вирус- содержащий материал | 0,1 | 0,5 | 0,5 вылить | ||||||
Физиоло- гический раствор | 0,9 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 |
1% взвесь эритроцитов | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 |
Возможный результат | +++ | +++ | +++ | +++ | +++ | ++ титр | + | - | - |
Если предварительно соединить вирус со специфической иммунной вируснейтрализующей сывороткой и потом добавить эритроциты, то такой инактивированный вирус теряет способность вызывать гемагглютинацию. Эта реакция получила название реакции торможения гемагглютинации (РТГА) и может быть использована для окончательной идентификации неизвестного гемагглютинирующего вируса – по специфической диагностической сыворотке, а также для выявления титра неизвестных антител в сыворотке больного – по известным вирусным антигенам-диагностикумам.
Затрудняет применение РТГА наличие в сыворотках крови человека и животных неспецифических вирусных ингибиторов, которые могут вызвать неспецифическое торможение гемагглютинации. Выпускают противовирусные диагностические сыворотки, освобожденные от ингибиторов. РТГА ставят обычно на досках с лунками по методике, сходной с РГА.
2.2.2. Техника постановки РТГА
РТГА с целью идентификации вируса гриппа рода А. На плексигласовой доске готовят три ряда двукратных возрастающих разведений (в объёме 0,25 мл) диагностических противогриппозных сывороток А(Н1N1), А(Н2N2) и А(Н3N2) от 1:10 до 1:320 (до титра сывороток). Во все лунки, кроме контрольной, добавляют по 0,25 мл рабочей дозы вируса (разведение 1:40), в котором содержится 4 гемагглютинирующие единицы (табл. 5). После выдерживания реакции в течение 1 часа при комнатной температуре во все лунки добавляют по 0,5 мл 1% взвеси эритроцитов кур. Учёт результатов проводят через 30-40 минут. В примере торможение произошло сывороткой А(Н3N2), следовательно выделен вирус гриппа, принадлежащий к подтипу А(Н3N2).
Таблица 5
Схема постановки РТГА для определения типовой принадлежности
вируса гриппа А
Ингредиенты РТГА в мл | Номера лунок, разведения иммунной сыворотки | ||||||
1 | 2 | 3 | 4 | 5 | 6 | 7 | |
1:10 | 1:20 | 1:40 | 1:80 | 1:160 | 1:320 | Контр. | |
Диагностические противогриппозные сыворотки в разведении 1:5 | 0,25 | 0,25 | 0,25 | 0,25 вылить | |||
Физиологический раствор | 0,25 | 0,25 | 0,25 | 0,25 | 0,25 | 0,5 | |
Вируссодержащая аллантоисная жидкость в разведении 1:40 | 0,25 | 0,25 | 0,25 | 0,25 | 0,25 | 0,25 | - |
Взвесь эритроцитов кур 1% | Выдерживание 1 час при комнатной температуре | ||||||
0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | 0,5 | |
Возможный результат Диагности - А(Н1N1) +++ +++ +++ +++ +++ ++ - ческие А(Н2N2) +++ +++ +++ +++ +++ +++ - сыворотки А(Н3N2++ - (нейтрализация) |
РТГА ставят и для определения наличия и титра антител в сыворотке больного. Для этого готовят последовательные двукратные разведения сыворотки в объёме 0,25 мл и соединяют их с 0,25 мл известного антигена (содержащего 4 гемагглютинирующие единицы), чтобы произошла нейтрализация вируса. Затем в пробирки вносят по 0,5 мл взвеси эритроцитов. За титр сыворотки принимают то наибольшее её разведение, в котором ещё наблюдается торможение гемагглютинации (1:160).
2.2.3. Реакция нейтрализации вирусов in vivo
Основными составляющими компонентами этой реакции являются: иммунная вируснейтрализующая (опытная) и нормальная (контрольная) сыворотки одного и того же вида животного или человека; содержащий живые вирусы материал, являющийся антигеном; биологические объекты (мыши, куриные эмбрионы), на которых обнаруживают вируснейтрализующее действие иммунной сыворотки. Применяют два варианта постановки реакции нейтрализации. При одном из них берут постоянную дозу сыворотки и различные разведения вируса, при втором – постоянное разведение вируса и различные разведения сыворотки. Во всех случаях последовательность проведения реакции следующая:
1) предварительный контакт вируссодержащего материала с соответствующей иммунной сывороткой (один из этих компонентов заведомо известен, наличие другого определяется);
2) введение этой смеси в чувствительную биологическую систему (белым мышам или куриным эмбрионам);
3) учёт наличия или отсутствия нейтрализации.
Сыворотки получают либо от больных людей, либо применяют диагностические сыворотки от иммунизированных животных. Перед постановкой реакции сыворотки инактивируют, прогревая при температуре 56-600С.
Антигены (вируссодержащие взвеси) получают из материала от больных вирусными инфекциями, готовят из органов и тканей заражённых вирусом животных, куриных эмбрионов, а также из инфицированных клеточных культур и подвергают очистке. Перед употреблением антиген титруют для определения его активности.
Биологические объекты выбирают с учётом свойств вируса, чувствительности самого объекта к действию вируса, а также задач и условий исследования.
Постановка реакции нейтрализации (I вариант)
Ставят для идентификации выделенного вируса. Для постановки реакции берут два ряда пробирок, из которых первый ряд является опытным, а второй – контрольным. В первый ряд пробирок наливают определённое количество неразведённой диагностической иммунной сыворотки, во второй – контрольный – такое же количество неразведённой нормальной сыворотки того же вида животного. Отдельно в пробирках готовят несколько разведений вируса, чтобы при добавлении их в пробирки опытного, контрольного ряда получились последовательные 10-кратные разведения вируса (например, 10-1, 10-2, 10-3 и т. д.). Сыворотки и добавляемые разведения вируса должны быть в одинаковых объёмах, например по 0,2 мл. Пробирки с приготовленной смесью ставят в термостат при температуре 370С обычно на 1-2 часа. После этого опытные и контрольные смеси вводят мышам или в куриные эмбрионы. Каждым разведением для достоверности опыта заражают не менее 4 мышей или куриных эмбрионов.
За заражёнными животными (опытными и контрольными) устанавливают наблюдение в течение 2-х недель, регистрируют заболевших и погибших мышей. Учёт реакции проводят путём сопоставления количества погибших опытных и контрольных мышей от соответствующего разведения вируса. Определяют 50%-ную смертельную дозу – LD50, проводя подсчёт по Риду и Менчу. Высчитывают индеек нейтрализации (ИН) по формуле, определяющей количество доз, нейтрализуемых сывороткой:
Титр вируса в опыте с нормальной сывороткой
ИН = --
Титр вируса в опыте с иммунной сывороткой
ИН менее 10 – отрицательный;
ИН – 11-49 – сомнительный;
ИН равный 50 и выше – положительный.
Заражённые эмбрионы инкубируют при температуре 35-370С от 48 до 72 ч. Затем яйца вскрывают, аллантоисную или амниотическую жидкость отсасывают и в ней определяют наличие вируса при помощи реакции гемагглютинации. Если гемагглютинация наступила в контроле (смесь нормальной сыворотки с разведениями вируса) и отсутствует в опыте (смесь иммунной сыворотки с разведениями вируса), то сыворотка обладает нейтрализующим действием, Конечный титр вируса в контроле и в опыте определяют путём статистической обработки.
Постановка реакции нейтрализации (II вариант)
Реакцию ставят с различными разведениями исследуемой сыворотки и постоянной дозой известного живого вируса. Для этого к двукратным возрастающим разведениям сыворотки добавляют равные объёмы определённого разведения вируса (рекомендуется брать 100 или 1000 LD50 вируса). Далее поступают как и в I варианте, то есть выдерживают опытные и контрольные смеси 1-2 часа в термостате, после чего вводят их мышам или куриным эмбрионам.
При постановке реакции нейтрализации на мышах за титр изучаемой сыворотки принимают разведение, дающее защитный эффект у 50% мышей. В реакции на куриных эмбрионах титром сыворотки считается то наибольшее её разведение, которое ещё способно задержать гемагглютинирующее действие вирусов у 50% заражённых куриных эмбрионов.
2.3. Молекулярно-генетические методы исследования в диагностике вирусных инфекций
К таким методам относятся молекулярная гибридизация и полимеразная цепная реакция, которые позволяют обнаружить присутствие в исследуемом материале даже единичных копий генов определяемых вирусов (ДНК или РНК с определённой нуклеотидной последовательностью), и тем самым доказать наличие соответствующей инфекции.
2.3.1. Молекулярная гибридизация (метод молекулярных зондов)
Метод основан на способности двухспиральной ДНК к денатурации и ренатурации. Денатурация – это расхождение цепей при нагревании ДНК до 80-1000С или обработке её щёлочью. Ренатурация – воссоединение цепей с помощью водородных связей при снижении температуры до 40-600С (отжиг) и приобретение ДНК первоначальной двухспиральной структуры. Разъединённые цепи ДНК способны к гибридизации с фрагментами других односпиральных ДНК, имеющих комплементарные участки расположения нуклеотидов. К гибридизации комплементарных нитей способна также РНК, образуя комплексы ДНК – РНК или РНК – ДНК.
Необходимые для молекулярной гибридизации фрагменты ДНК или РНК, с помощью которых выявляют наличие в исследуемом материале комплементарных нитей нуклеиновой кислоты, называются молекулярными зондами. Молекулярные зонды готовят из нуклеиновых кислот, выделенных из различных вирусов, иногда используют вирусную и-РНК, но чаще зондом служит клонированная рекомбинантная ДНК. Имеются наборы молекулярных зондов для определения многих вирусов.
При постановке реакции молекулярной гибридизации зонды метят радиоактивной (Р32), флюоресцентной или биотиновой меткой, соединяют с исследуемым материалом, содержащим определяемую нуклеиновую кислоту, подвергающуюся денатурации. Если зонд комплементарен цепи определяемой нуклеиновой кислоты, происходит гибридизация в комплементарном участке.
После отжига зонд оказывается включённым в ренатурированную нуклеиновую кислоту и может быть обнаружен по имеющейся метке. Выявление наступившей молекулярной гибридизации позволяет установить природу определяемого вируса. Чувствительность этого метода составляет 10-14 г/мл.
2.3.2. Полимеразная цепная реакция (ПЦР) или локальная
амплификация нуклеиновых кислот (ЛАНК)
ПЦР, как и молекулярная гибридизация, основана на способности ДНК к денатурации и ренатурации и на комплементарности цепей ДНК. Важным принципом реакции является использование термостабильной ДНК-полимеразы, при участии которой происходит амплификация – умножение определяемых генов или фрагментов с определённой нуклеотидной последовательностью ДНК. В результате реакции исследуемый генетический материал накапливается в значительном количестве и может быть легко выявлен и идентифицирован. Чувствительность этой реакции составляет 10-18 г/мл.
В реакции участвуют следующие ингредиенты:
· определяемая ДНК вирусов в испытуемом биологическом материале, который предварительно концентрируется;
· праймеры 2-х типов (олигонуклеотиды) – короткие цепочки ДНК с нуклеотидной последовательностью комплементарной 3,-концам каждой из двух цепей определяемой ДНК. Праймеры получают из нуклеиновых кислот различных вирусов, их нуклеотидную последовательность определяют методом секвенирования;
· свободные нуклеитиды (дезоксирибонуклеозидтрифосфаты 4-х типов с различными азотистыми основаниями) – необходимый материал для осуществления амплификации;
· фермент термостабильная ДНК-полимераза – производит достройку комплементарных цепей ДНК из пула свободных нуклеотидов. Фермент выделяют из бактерий Thermus aquaticus или получают генно-инженерным методом.
Сущность ПЦР состоит в том, что определяемая ДНК, находящаяся в тестируемом биологическом материале, подвергается денатурации. Затем праймеры 2-х типов в условиях отжига присоединяются, вследствие их комплементарности, к 3,-концам каждой из антипараллельных цепей, восстанавливая на этом участке двухспиральную структуру ДНК. Праймеры служат «затравками» для последующей достройки цепей ДНК, осуществляемой термостабильной ДНК-полимеразой, которая использует для этой цели свободные нуклеотиды.
В результате одного цикла ПЦР количество молекул определяемой ДНК удваивается, то есть происходит амплификация ДНК. Обычно проводят 25-40 повторных циклов амплификации и в итоге за 2-3 часа получают миллионы копий специфического фрагмента ДНК вирусов.
ПЦР проводят в 0,5-1,5 мл микроцентрифужных пробирках в амплификаторах, которые автоматически регулируют смену температуры. Каждому из 3-х этапов цикла амплификации – денатурации ДНК, отжига и достройки – необходима инкубация образцов при различном температурном режиме.
1. Денатурация – разъединение определяемой двухспиральной ДНК на две изолированные цепи при нагревании до 90-950С в течение 0,5-1,0 мин.
2. Отжиг – восстановление двухцепочечной структуры определяемой ДНК в области присоединения комплементарного праймера – проводится при температуре 40-600С 0,5 мин.
3. Удлинение (элонгация) – достройка каждой цепи определяемой ДНК доисходного двухцепочечного состояния с помощью термостабильной ДНК-полимеразы – проводится при температуре 70-750С в течение 2-5 мин.
Наличие ДНК после повторных циклов амплификации определяют с помощью электрофореза в полиакриламидном геле, авторадиографией или другими методами.
С помощью ПЦР возможно определение не только нуклеотидной последовательности ДНК, но также РНК и, следовательно, выявление РНК-вирусов (для этого в реакцию вводят обратную транскриптазу). ПЦР особенно ценен для диагностики латентных вирусных инфекций и ВИЧ-инфекции.
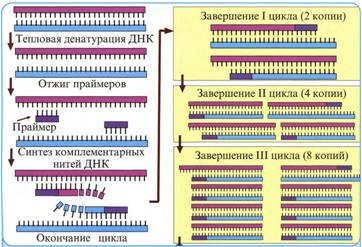
Рис. 8. Схема полимеразной цепной реакции
Контрольные вопросы
1. Индикация вируса по его цитопатическим действиям (ЦПД) и определение титра вируса. Идентификация вируса по нейтрализации ЦПД.
2. Метод гемадсорбции, его практическое применение, идентификация вируса в реакции задержки гемадсорбции.
3. Метод бляшек. Идентификация вирусов методом бляшек, титрование антител.
4. Метод цветной пробы, методика постановки, практическое применение.
5. Реакция нейтрализации по цветной пробе, её сущность, применение.
6. Реакция гемагглютинации (РГА), вызываемая вирусами. Практическое применение РГА. Методика постановки.
7. Сущность реакции торможения гемагглютинации (РТГА). Методика постановки. Практическое применение.
8. Реакция нейтрализации вирусов in vivo, способы постановки. Практическое применение.
9. Молекулярно-генетические методы исследования, их сущность. Принцип метода молекулярной гибридизации.
10. Полимеразная цепная реакция (ПЦР). Ингредиенты реакции, их характеристика, значение амплификации. Сущность ПЦР.
11. Техника постановки ПЦР, практическое применение и оценка, как экспресс-метода диагностики вирусных инфекций.
Тематика докладов
1. Современные методы диагностики распространённых вирусных инфекций.
2. Индикация и идентификация вирусов гриппа.
3. Молекулярно-генетические методы анализа полиморфизма генов.
4. Молекулярно-генетические методы анализа экспрессии генов.
ЛИТЕРАТУРА
1. Биргер по микробиологическим и вирусологическим методам исследования. 3-е изд. – М.: Медицина, 1982.
2. Большой словарь медицинских терминов / Сост. – М.: , 2007.
3. Вирусология. Методы / Под ред. Б. Мейхи. – М.: Мир, 1988.
4. А., , Широбоков и санитарная микробиология. – М.: АСАDEMA, 2003.
5. Голубев по применению клеточных культур в вирусологии. – Л., 1986. Электронный учебник.
6. ПЦР «в реальном времени» / Под ред. ; 2-е изд., испр. И доп. – М.: БИНОМ. Лаборатория знаний, 2009.
7. Вирусология. Т. 1-3. – М.: «Мир», 1989.
8. www. *****/rus/all/mvb journals.shtml
9. www.
10. www. medmaster.net
СОДЕРЖАНИЕ
ВВЕДЕНИЕ…………………………………………………………………….... 3
Раздел 1. Культивирование вирусов ……………………………………………4
1.1. Подготовка материала для вирусологических исследований…………….4
1.1.1. Правила работы в вирусологической лаборатории…………………....4
1.1.2. Материалы, исследуемые при вирусных инфекциях…………….........5
1.1.3. Обработка вируссодержащего материала………………………………5
1.1.4. Микроскопические методы исследования в вирусологии…………….6
1.2. Культуры клеток в вирусологии и методы их получения………………..9
1.2.1. Питательные среды…………………………………………………….11
1.2.2. Типы клеточных культур………………………………………………12
1.3. Культивирование вирусов в развивающихся куриных эмбрионах……...16
1.3.1. Строение куриного эмбриона………………………………………….17
1.3.2. Заражение куриного эмбриона на хорионаллантоисную оболочку....18
1.3.3. Заражение в аллантоисную полость……………………………...........20
1.3.4. Заражение в полость амниона………………………………………….20
1.4. Культивирование вирусов путём заражения лабораторных животных…22
1.4.1. Интраназальное заражение………………….……………………….....23
1.4.2. Интрацеребральное заражение…………………………………............24
Контрольные вопросы………………………………………………………..….25
Тематика докладов……………………………………………………………….25
Литература………………………………………………………………………..25
Раздел 2. Индикация и идентификация вирусов………………………………26
2.1. Методы индикации и идентификации вирусов в клеточных культурах..26
2.1.1. Цитопатическое действие (ЦПД) вируса в культуре клеток…….......27
2.1.2. Реакция гемадсорбции…………………………………………………29
2.1.3. Метод бляшек…………………………………………………………..30
2.1.4. Цветная проба…………………………………………………………..32
2.2. Применение реакции гемагглютинации (РГА), реакции
торможения гемагглютинации (РТГА) и биологических
моделей для индикации и идентификации вирусов……………………...35
2.2.1. Техника постановки РГА………………………………………………36
2.2.2. Техника постановки РТГА……………………………………………..37
2.2.3. Реакция нейтрализации вирусов in vivo………………………………39
2.3. Молекулярно-генетические методы исследования
в диагностике вирусных инфекций……………………………………..41
2.3.1. Молекулярная гибридизация (метод молекулярных зондов)………42
2.3.2. Полимеразная цепная реакция (ПЦР) или локальная
амплификация нуклеиновых кислот (ЛАНК)……………………...........43
Контрольные вопросы ………………………………………………………...45
Тематика докладов……………………………………………………………...46
Литература ……………………………………………………………………..46

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
