

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
В составе флороценотического комплекса приморских экосистем четко видно господство Poaceae — 26 видов (23%), затем идут Сyperaceae — 17 видов (15%) и Asteraceae — 8 видов (7%). Высока роль сем. Chenopodiaceae — 7 видов (6%) и велико число семейств, имеющих по 1—2 вида в составе общей ППФ, — 23 семейства (72%). Наиболее богато представлены в общей ППФ морей следующие рода: Carex — 12 видов (10%), Puccinellia — 11 видов (7%), Atriplex — 4 вида (3,5%), по 3 вида (2,7%) имеют рода Eleocharis, Juncus, Cochlearia, Potentilla, Plantago, по 2 вида (по 1,7%) — Ruppia, Zostera, Triglochin, Agrostis, Deschampsia, Dupontia, Leymus, Salicornia, Cerastium, Saxifraga, Rhodiola, Senecio, Empetrum, Primula. Род Puccinellia — типично приморский, большинство его видов приурочены к засоленным местообитаниям береговых и континентальных районов. Виды рода Carex предпочитают влажные и слегка заболоченные элементы ландшафта, но с успехом приспособились и к специфическим условиям приморской полосы.
Эколого-географический анализ
Виды, входящие в состав ППФ, различаются по широте экологической амплитуды и объединены в следующие экологические группы:
I. Среди облигатных галофитов (эвгалофитов): а) гемиэвритопные виды, имеющие эколого-ценотический оптимум на эвгалогенных экотопах, но довольно обычны в мезогалогенных экотопах первичных и вторичных маршей; б1) гемистенотопные виды, предпочитающие эвгалогенные, реже мезогалогенные экотопы, с илистыми отложениями на первичных и на вторичных маршах; б2) гемистенотопные виды, предпочитающие эв-, реже миогалогенные экотопы песчано - и песчано-илистых отложений на вторичных маршах; в1) стенотопные виды, растущие на илистом аллювии эвгалогенных экотопов в устьях рек; в2) стенотопные виды, растущие на песчано - и песчано-илистых отложениях по берегам бессточных засоленных озер на вторичных маршах.
II. Среди факультативных галофитов (мезогалофитов): а) гемиэвритопные виды, предпочитающие задернованный илистый аллювий мезогалогенных экотопов на первичных, реже вторичных маршах; б) гемистенотопные виды, растущие на песчаном и песчано-галечном аллювии мезо-, реже миогалогенных экотопов приморских баров и кос (псаммофитон); б1) на песчаных пляжах; б2) на гребнях берегового вала; б3) на обратных скатах берегового вала; в) стенотопные виды, строго приуроченные к мезогалогенным экотопам, уклоняющимся в сторону некоторой нивальности местообитаний.
III. Толерантные виды (миогалофитов) включают группу гемиэвритопных видов, обычных на мио - и мезогалогенных экотопах, но часто встречающихся вне зоны засоления — на дренированных склонах в пойме рек, у подножия коренного берега, на речном аллювии, на рудеральных местообитаниях.
Анализ распределения эколого-ценотических групп приморских видов в ППФ морей российской Арктики по крупных выделам экотопов — илистые маршевые осушки (осушки), песчаные и песчано-галечные пляжи (псаммофитон), песчаные, песчано-галечные валы, косы и бары (валы) и экотонные, переходные зоны (экотон) — выявил абсолютное доминирование на осушках Puccinellia и Carex, в псаммофитоне — преобладание Leymus, Honckenya, Mertensia, на валах преобладают виды из родов Deschampsia, Armeria, Potentilla (рис. 2).
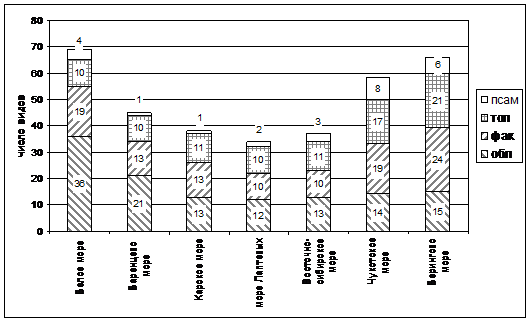
Рис. 2. Распределение эколого-ценотических групп приморских видов в парциальных приморских флорах арктических морей российской Арктики.
Примечание: псам — псаммофиты, тол — толерантные виды, фак — факультативные галофиты, обл — облигатные галофиты
Географический анализ парциальных приморских флор побережий морей российской Арктики проведен в соответствии с «принципом биогеографических координат», разработанным А. И. Толмачевым (Толмачев, 1932, 1939, 1986) и Б. А. Юрцевым (Юрцев, 1968, 1983, 1991; Юрцев, Камелин, 1991; Юрцев, Кучеров, 1999) для анализа северных флор. Сравнение соотношения широтных географических элементов показывает, что в общей ППФ преобладают виды арктической и бореальной фракций (рис. 3). В арктической фракции ППФ всех морей преобладают виды с циркумполярным ареалом, являющиеся автохтонными элементами приморской флоры — Carex subspathacea, Puccinellia phryganodes, Stellaria humifusa, Cochlearia arctica, Hippuris tetraphylla.
Число видов гипоарктической фракции практически не меняется в составе приморских флор, но варьирует от 16 (Белое море), 10 видов (Берингово море) до 4 видов (Карское море, море Лаптевых, Восточно-Сибирское море). Арктобореальная и бореальная фракции представлены в основном факультативными приморскими видами — 1—2 вида на побережьях Карского моря, моря Лаптевых, Восточно-Сибирского и Чукотского, 7—10 видов — Баренцево и Берингово моря и 23 вида — побережье Белого моря.
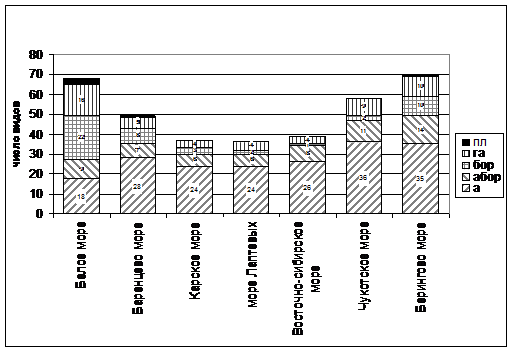
Рис. 3. Спектр широтных географических групп парциальных приморских флор морей российской Арктики. Примечание: пл — плюризональные виды, га — гипоарктические виды, бор — бореальные виды, абор — арктобореальные виды, а — арктические виды
Сравнение соотношения долготных географических элементов показывает, что в парциальных приморских флорах побережий морей российской Арктики преобладают виды с циркумполярным типом распространения. Среди циркумполярных видов господствуют арктические виды, в группе видов с азиатским типом распространения преобладают бореальные и гипоарктические. Существенную роль играют и амфиокеанические виды, доля которых несколько выше в приморских флорах на побережье Белого моря, чем на побережьях остальных арктических морей. Если рассматривать соотношения долготных элементов в 3 группах видов, имеющих разную экологическую амплитуду в парциальных флорах, то дифференциация выражена более значительно. Среди облигатных галофитов на побережьях арктических морей преобладают виды с циркумполярными и европейскими ареалами. Виды Puccinellia maritima, P. pulvinata, Bolboschoenus maritimus отмечены только на побережье Белого моря. Среди факультативных галофитов доля европейских и амфиокеанических видов наибольшая. Среди толерантных галофитов преобладают виды с евразиатским типом ареала и виды с циркумполярным распространением.
В ведущих семействах — Poaceae, Cyperaceae, Caryophyllaceae — практически одинаково число циркумполярных видов, сем. Chenopodiaceae богато представлено видами европейской фракции. Семейства с небольшим числом видов (1, 2, 3, 4) — Rosaceae, Primulaceae, Plantaginaceae, Asteraceae — лишены видов с амфиокеаническими ареалами.
В результате проведенного таксономического и эколого-географического анализа можно констатировать значительное сходство приморских парциальных флор побережий всех арктических морей. Наибольшее сходство отмечается в таксономической структуре ведущих семейств, которая остается практически одинаковой. Бóльшие расхождения имеются в широтной географической структуре, где доля бореальных видов в приморской флоре побережья Белого моря в 1,8 раза больше, чем на побережьях всех арктических морей.
На основании географического анализа видов ППФ можно предположить, что в формировании галофитного флороценотического комплекса основную роль играли древние эоарктические виды (шельфовые элементы) и их более поздние атлантические и берингийские дериваты (Тихомиров, 1944, 1946б; Толмачев, 1939, 1962; Юрцев, 1966, 1968, 1976б; Chapman, 1960, 1964; Каплин и др., 1991; Каплин, Селиванов, 1999; Павлидис и др., 1998). Кроме того, эта гипотеза находит свое подтверждение в результатах изучения распределения географо-флористических групп по типам местообитаний.
Полиплоидия
На настоящий момент изучены хромосомные числа большинства (более 80% состава флоры) видов сосудистых растений Чукотки (Юрцев и др., 2010), поэтому анализ уровней плоидности проведен нами у видов ППФ Чукотки. Выделено 3 группы видов, характеризующихся разными уровнями плоидности (в анализ не включались виды р. Carex и Pucciphippsiа czukczorum, характеризующиеся анеуплоидией): диплоиды, тетраплоиды и гексаплоиды и виды с более высоким уровнем плоидности. Среди приморских видов больше всего диплоидов — 17 видов (42% от видов, включенных в анализ), тетраплоидов — 16 видов (39%), гексаплоидов и видов с более высоким уровнем плоидности — 8 видов (19%). Среди эвгалофитов, являющихся пионерами зарастания приморских маршей, преобладают диплоиды; среди мезо - и миогалофитов, поселяющихся на вторичных маршах и на песчано-галечных косах, преобладают тетраплоиды и полиплоиды более высоких уровней плоидности. При рассмотрении географического распространения полиплоидов в пределах Чукотки среди арктических видов преобладают диплоиды — 10 видов (60%); тетраплоиды — 4 вида (23%) и видов с более высоким уровнем плоидности 3 (17%). У наиболее крупной в долготном отношении группы циркумполярных видов диплоиды преобладают: 8 диплоидов (62%) и 5 тетраплоидов (38%). Гексаплоиды и виды с более высоким уровнем плоидности отсутствуют. В главе также представлен конспект видов галофитного флористического комплекса. Для каждого вида приведены номенклатурная цитата, эколого-ценотическая и географическая характеристики. Пример цитаты:
Puccinellia angustata (R. Br.) Rand. et Redf. 1894, Fl. of Mount. Desert. Isl. : 181; Hulten, 1968, Fl. Alaska a. neighb. terr. : 159; Цвелев, 1964, Аркт. фл. СССР, 2 : 195. Bocher, 1978, Grønland Flora : 289; Elven, 2005, Nørsk flora : 1087; Поспелова, 2007, Флора сосуд. раст. Таймыра и сопред. терр. : 128; Юрцев и др., 2010. Конспект фл. Чукот. тундры: 108. — Poa angustata R. Br. 1824, Suppl. to App. Parry’s First. Voy., Bot. : 287. — Puccinellia angustata subsp. angustata, Цвелев, 1976, Злаки СССР: 509.
Глава 6. Критико-систематические исследования приморских видов из родов Puccinellia, Suaeda, Cochlearia, Honckenya, Potentilla, Arctanthemum
5.1. Puccinellia Parl.
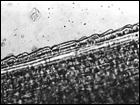
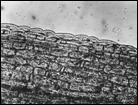
Помимо анализа морфометрических признаков (характеристика побегов, размеры листьев, метелки, колоска, цветковых чешуй, размеры и форма колосковых и цветковых чешуй, пыльников) изучено строение эпидермиса верхней стороны листовой пластинки стеблевых листьев. Выявлено 6 типов строения эпидермы (рис. 4): 1 — эпидерма правильно ячеистая с многочисленными сосочками, побережье Охотского моря, Аляска, Гренландия, 2 — эпидерма правильно ячеистая с более мелкими клетками и хорошо выраженными сосочками, Камчатка, побрежье Охотского моря, о-ва Б. и М. Шантар, 3 — эпидерма правильно ячеистая с довольно ровными клетками и очень редкими сосочками, Новая Земля, п-ов Канин, 4 — эпидерма неправильно ячеистая с немногими, но ясно выраженными сосочками, побережье Чукотского моря, Берингов пролив, 5 — эпидерма неправильно ячеистая с несколько вздутыми клетками и немногочисленными сосочками и бугорками, Таймыр, Новосибирские о-ва, о. Б. Ляховский, о. Айон, о. Врангеля, побережье Берингова пролива, 6 — эпидерма неправильно ячеистая с немного ромбовидными клетками, большей частью без сосочков, Шпицберген, о. Колгуев, о. Котельный, низовья реки Лены, мыс Барроу, западное побережье Гренландии. Показано, что современные подвиды рода Puccinellia Parl. — Р. phryganodes subsp. phryganodes, P. phryganodes subsp. geniculata, и P. phryganodes subsp., sibirica (рис. 4) имеют одинаковое строение эпидермальной ткани: она неправильно ячеистая, клетки с хорошо выраженными обильными сосочками. Наши данные подтверждают предположение Н. Н. Цвелева (Цвелев, 1967) о том, что подвиды P. phryganodes s. l. сформировались во время миграции предковой формы типа P. phryganodes subsp. geniculata на восток и на запад от Берингова пролива по северным побережьям Евразии и Америки.
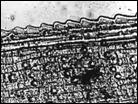 | 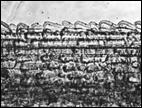 |  |  |
1 | 2 | 3 | 4 |
Рис. 4. Эпидермис верхней поверхности стеблевых листьев Puccinellia phryganodes subsp. phryganodes. По образцам: 1 — Охотское море (typus: Puccinellia geniculata V. Krecz.), 2 — Гренландия, 3 — Шпицберген, 4 — п-ов Канин
5.2. Suaeda Forssk. ex J. F. Gmel.
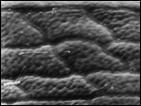
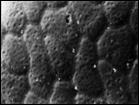
Suaeda arctica Jurtz. et Petrovsky — арктический приморский эвгалофитный вид, известный только из одного, типового местонахождения на Чукотском п-ве. Для уточнения таксономических связей S. arctica изучены наиболее близкие к ней виды: S. maritima (L.) Dumort., распространенный на атлантическом побережье Европы, и S. corniculata (С. A. Mey.) Bunge, обычный на солончаках в центральной Якутии. На основании изучения всего комплекса признаков, а в особенности ультраструктуры поверхности семени, сделан вывод, что S. arctica обнаруживает более тесные родственные связи с S. maritima, чем с S. corniculata. На рис. 5 (1, 2) показанa ультраструктура поверхности семени типового образца Suaeda arctica.
 |  |  |  |
1 | 2 | 3 | 4 |
Рис. 5. Микроструктура поверхности семени Suaeda arctica Jurtz. et Petrovsky: 1 — Typus! Западная Чукотка, Певек, х 240, 2 — там же, х 860, 3 — Suaeda maritima (L.) Dumort, из приатлантической Европы, х 860, 4 — Suaeda corniculata (C. A. Mey.)
Bunge из центральной Европы (Париж) х 360
5.3. Cochlearia L.
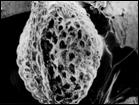
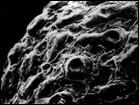
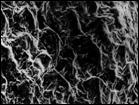
На побережьях российской Арктики род Cochlearia L. представлен двумя близкородственными видами: С. groenlandica L. и С. arctica Schlecht. ex DC, последний состоит из 2 подвидов: subsp. arctica u subsp. oblongifolia (DC.) Petrovsky (Аркт. флора СССР, 1975). Нами установлено, что основные различия между растениями состоят в строении чашелистиков, плодов и семян, а также в ультраструктуре поверхности семян. По нашим данным, у С. arctica чашелистики лиловатые, стручочки невздутые, семена яйцевидные, рыжеватые, в числе 14—16; ультраструктура поверхности семени в виде мелкоскладчатых конических, плотно (но не рядами) расположенных пологих бугорков с углублением на верхушке. У С. oblongifolia чашелистики почти прозрачные, стручочки вздутые, семена яйцевидные, реже яйцевидно-шаровидные, темно-бурые, в числе 10—15; ультраструктура поверхности семени в виде мелкоскладчатых, параллельно расположенных рядов, вытянутых пологих выступов без углубления на верхушке. У С. groenlandica чашелистики беловатые с широкими прозрачными краями, стручочки вздутые, семена шаровидные, реже яйцевидно-шаровидные, светло-рыжие, в числе 8—12; ультраструктура поверхности семени имеет вид очень плотно расположенных мелкоскладчатых чешуек (рис. 6). Все названные признаки рассмотренных таксонов являются выдержанными, что дает нам основание считать С. oblongifolia самостоятельным видом, а не подвидом С. аrctica.
 |  |  |  |
1 | 2 | 3 | 4 |
Рис. 6. Микроструктура поверхности семени Cochlearia arctica (1, 2), C. groenlandica (3), C. oblongifolia (4). По образцам: 1, 2 — северное побережье Чукотки, 3 — юго-восточное побережье Чукотки, 4 — южное побережье Чукотки
5.4. Potentilla L.
Весь просмотренный материал по Potentilla egedei subsp. egedei показал большую изменчивость растений по форме зубцов листочков и опушению нижней стороны листьев. Гренландские и лапландские экземпляры, а также растения с Кольского п-ва имеют округлые листочки в основном с неопушенной нижней поверхностью и городчатыми или почти округлыми зубцами. У них ярче выражена антоциановая окраска листочков и чашечки. Растения с арктического побережья Сибири (окрестности пос. Хараулах) очень маленькие, с совсем голыми листьями. Почти все чукотские и американские экземпляры приближаются по форме к типовому изображению, т. е. это более крупные растения с заостренными зубцами листочков и более густым опушением их нижней стороны. Таким образом, в типовом подвиде намечаются три слабо обособленные морфологические группы; признаки у растений этих групп весьма расплывчаты и не могут, на наш взгляд, служить основанием для выделения разновидностей (Hulten, 1968). Нами предложена новая комбинация Potentilla egedii subsp. pacifica (Howell) L. Sergienko comb, nov. — P. pacifica Howell, 1898, Pl. North-West. Amer. 1 : 179. В результате изучения диапазона изменчивости приморских видов группы P. anserina L. s. l. в Евразии и Северной Америке мы пришли к выводу о произрастании только 2 видов: P. anserina и P. egedei с 2 подвидами: subsp. egedei с арктическим в основном ареалом и subsp. pacifica (Howell) L. Sergienko, распространенному на тихоокеанском побережье Азии и Северной Америки и атлантическом побережье Северной Америки (40°—60° с. ш.).
5.5. Honckenya Ehrh.
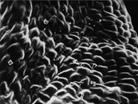
Для циркумполярного арктического побережья авторами различных обработок приводится от 2 до 5 видов рода Honckehya Ehrh. Были изучены признаки: длина междоузлий, форма и размер пластинки листа, толщина стенок плода-коробочки, форма, цвет, типы и строение поверхности семян. Выделены 2 группы, различающиеся по размеру, форме, цвету и структуре поверхности семян (рис. 7). На основании полученных данных сделан вывод об отсутствии различий между H. oblongifolia, H. diffusa и H. frigida по всем признакам. Мы не считаем возможным различать эти виды в качестве самостоятельных, поэтому Н. frigida и H. diffusa включаем в синонимы к H. oblongifolia.
 |  |  |  |
1 | 2 | 3 | 4 |
Рис. 7. Микроструктура поверхности семени Honckenya diffusa (Hornem.) A. Love: 1 — побережье Белого моря (Кемь), 2 — Сахалин, 3 — Якутия, губа Буорхая, 4 — Тип Honckenya frigida Pobed.: Берингов пролив, мыс Дежнева
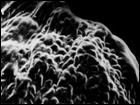 | ||
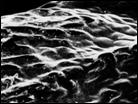 | 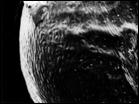 | 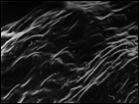 |
1 | 2 | 3 | 4 |
Рис. 8. Микроструктура поверхности семени Honckenya diffusa (Hornem.) A. Love (1, 2)
и Honckenya peploides (L.) Ehrh. (3, 4): 1 — побережье Баренцева моря (Териберка), 2 — Тазовский п-ов, (3, 4), 3, 4 — побережье Балтийского моря, Финский залив
5.6. Arctanthemum (Tzvel.) Tzvel.
Для российской Арктики и Дальнего Н. Цвелев (1987) приводит 3 подвида облигатного галофитного вида Arctanthemum arcticum (L.) Tzvel.: subsp. arcticum, subsp. polare (Hult.) Tzvel. и subsp. kurilense (Tzvel.) Tzvel. Мы считаем, что A. arcticum subsp. kurilense представляет собой серию форм, связанную с A. arcticum subsp. arcticum и A. arcticum subsp. polare постепенными переходами, и поэтому его нельзя считать самостоятельным подвидом. Мы рассматриваем A. arcticum subsp. kurilense в качестве синонима типового подвида A. arcticum subsp. arcticum .
Глава 7. Структура и динамика растительного покрова
приморских экосистем российской Арктики
Развитие и формирование растительного покрова приморских экосистем российской Арктики обусловлено направленным действием одного или комплекса узко и четко отграниченных в пространстве факторов среды (заливание и подтопление морской водой и структура почвенного слоя). Первичные сукцессии данных растительных сообществ возникают на лишенной растительности и не имеющих фитоценотической среды поверхности и являют собой основополагающую часть общего процесса динамики растительности приморской полосы.
Среди всех морей российской Арктики Белое и Баренцево отличаются наибольшим видовым богатством маршевой флоры. Береговая линия побережья Белого моря извилиста и очень разнообразна, поэтому распределение маршей по побережью весьма неравномерно. Для Кандалакшского залива на западе характерны скалистые абразионные и абразионно-аккумулятивные берега, в то время как на востоке преобладают обширные аккумулятивные формы. Широкие глинистые или песчаные осушки до двух километров есть в устьях рек Мезень, Несь, Чижа. По побережью Онежского залива, у мыса Лопшеньгского, распространены огромные песчаные пляжи с дюнами шириной до 500 м и длиной 200—300 км. На побережье Белого моря зона, занятая маршевой растительностью, представляет собой три уступа, полого спускающихся к морю. В нижней, или пионерной, зоне, покрытой морской водой более четырех часов в сутки, растения не образуют сомкнутых сообществ. Эта часть марша обычно представляет собой открытый илистый субстрат с редким покрытием из водорослевых корочек и пионерного вида Eleocharis uniglumis (рис. 9). В устьях некоторых рек к нему добавляется Salicornia europaea и Bolbochoenus maritimus. Средняя, наиболее задернованная, часть марша занята приморской растительностью, где видовое разнообразие галофитов достаточно велико, а в роли основного доминанта выступает Carex subspathacea. На участках морского побережья, покрытых свежими наносами песка, обычна Honckenya oblongifolia, на более удаленных от моря располагается полоса Leymus arenarius и Festuca rubra. При переходе маршей от среднего уровня к высокому местами появляются следы накопления торфа. Верхняя часть маршевой зоны представляет собой экотонный участок, куда проникают виды местной флоры, способные выносить незначительное засоление почвы.
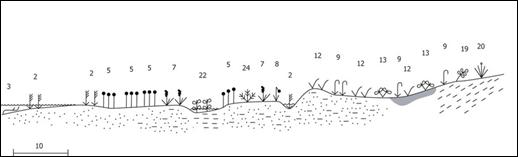
Рис. 9. Эколого-динамический ряд приморской маршевой растительности на Поморском и Карельском берегах Белого моря. Условные обозначения: приморские сообщества с доминированием: 2 — Bolboshoenus maritimus, 3 — Zostera marina, 5 — Eleocharis uniglumis, 7 — Triglochin maritima, 8 — Tripolium vulgare, 9 — Juncus gerardii, 12 — Carex subspathacea, 13 — Potentilla egedei s. l., 19 — Lathyrus japonicus ssp. pubescens,
20 — Phragmites australis, 22 — Hippuris tetraphylla, 24 — Stellaria humifusa
В Баренцевом море на Мурманском побережье берега, как правило, скалистые, поэтому участков с выраженной приморской растительностью немного, их можно найти только в защищенных от волнения кутовых частях губ. Наиболее разнообразные маршевые сообщества (отчасти похожие на беломорские) встречаются на побережье Чешской губы. На берегах Печорского моря из-за действия прибоя и морских льдов низкие маршевые зоны часто лишены растительности. Луга низкого уровня, длительно и часто заливаемые, расположены полосами вдоль илистых берегов. Их растительный покров исключительно однообразен: это ярко-зеленый плотный, но низкий (2—4 см) ковер из побегов Puccinellia phryganodes с примесью Carex subspathacea и Stellaria humifusa. Теми же растениями покрыты илистые осушки в северной, низменной, части Малоземельской тундры между Печорской и Колоколковой губами, сложенной передутыми песками. На восточном, илистом, берегу Колоколковой губы располагаются обширные, до четырех километров шириной, приморские луга нижнего уровня с теми же доминантами в растительном покрове. На лугах среднего уровня развиты простые по своему строению луговые ассоциации из почти чистых зарослей Carex subspathacea и Dupontia psilosantha. Все производные группировки лугов данного уровня связаны с Carex subspathacea, Calamagrostis deschampsioides, Stellaria humifusa и Potentilla egedеi. В большинстве баренцевоморских заливов и соленых лагун марши высокого уровня занимают сообщества c доминированием Carex subspathacea, Festuca rubra и Parnassia palustris. Здесь может развиваться моховой покров, чего, как правило, не бывает в зонах, расположенных ниже.
Характерные черты побережья Карского моря — геологоморфологическая неоднородность, повсеместное распространение многолетней мерзлоты и долгий срок жизни припайных льдов, которые существенно ограничивают воздействие волн на берег. Вдоль высоких абразионных берегов западного побережья Ямала приморская растительность тянется узкой полосой, окаймляет острова Белый и Шокальского, а также северное побережье Гыданского п-ва. Ее участки встречаются на берегах Обской губы и в Енисейском заливе, куда доходят соленые воды Карского моря. На юго-западном побережье Карского моря основу приморских маршевых сообществ составляют арктические циркумполярные виды — Puccinellia phryganodes, Carex glareosa, Carex subspathacea, Stellaria humifusa, а также Calamagrostis deschampsioides. В районе Харасавея на п-ве Ямал найден Triglochin maritimа, характерный для атлантической приморской флоры. Полоса приморской растительности с характерным для Байдарацкой губы пространственным распределением тянется вдоль всего полуострова Марре-Сале, расширяясь в местах впадения ручьев. Вся кутовая часть Обской губы заполнена почти пресными и солоноватыми водами (не более 5—8‰), по берегам произрастает обычная тундровая прибрежно-водная растительность с незначительной примесью Dupontia psilosantha и Arctophila fulva.
На Таймыре почти повсеместно распространены ледниковые формы рельефа, как денудационные (не измененные морем), так и аккумулятивные. Побережье начинается с песчано-илистого пляжа, ежедневно заливаемого морскими водами, на котором до 30% площади покрывают озерки-лужицы, а маршевая растительность сложена монодоминантной группировкой Puccinellia phryganodes. За пляжем следует менее сырая низина, периодически заливаемая морскими водами. Грунт там более стабильный, растительность представлена осоковыми галофитными сообществами. Выше расположена полоса, заливаемая лишь изредка, при сильных нагонных ветрах, где сырые пониженные участки перемежаются с песчаными повышениями. Маршевые биотопы и сообщества находятся под охраной в государственном заповеднике «Ненецкий», а также в нескольких региональных ООПТ.
В море Лаптевых приливные явления выражены слабо (высота прилива не более 0,8 м), но очень велики штормовые нагоны и сгоны, амплитуда которых достигает 2 и даже 5 м. Небольшие участки маршевой растительности имеются в Хатангском заливе, в районе Терпей-Тумуса (Анабарский район) и дельты реки Яны. Дельта реки Лены, крупнейшая на побережье Северного Ледовитого океана, имеет площадь 28,5 тыс. км2. Восточная часть дельты состоит из множества островов и проток. Во время половодья многие острова затопляются, из-за накопления на них илистых наносов и их уровень может повышаться до 5 м, однако на берегах островов сохраняются злаково-разнотравные приморские сообщества, хотя и с низким, всего лишь до 20%, проективным покрытием маршевых видов, таких как Calamagrostis deschampsioides, Triglochin maritima, Dupontiа psilosantha, Stellaria humifusa, Potentilla egedei, Armeria maritima, Arctanthemum arcticum ssp. polare. В западной части побережья Восточно-Сибирского моря широко распространены термоабразионные берега; крупные реки, в частности, Индигирка, Колыма, Алазея, Большая Куропаточья, Чукочья (протока Колымы) выносят в море огромное количество взвеси, из-за чего в устьях маршевая растительность развита слабо и выражена только на тех участках устьевой области, которые защищены от действия волновой эрозии. В устье реки Большая Куропаточья илистые наносы легко перемещаются, а берег отступает со скоростью около 1,06 м/год (Григорьев, Куницкий, 2000), из-за чего маршевые сообщества постоянно находятся на ранних стадиях сукцессионного процесса. К востоку от дельты Колымы берег выровнен и состоит из отрезков абразионных клифов, чередующихся с низкими аккумулятивными пляжами и ветровыми осушками. На абразионных берегах растительности нет, поскольку грязевые потоки с береговых клифов целиком покрывают обнажающийся пляж. Низкие осушки в некоторых местах слегка зарастают Puccinellia phryganodes, опять же с примесью Carex subspathacea, Stellaria humifusa, но эту группировку часто стирают наползающие на берег морские льды или смывают грязевые потоки. Берег наиболее глубоко вдающегося в сушу залива Восточно-Сибирского моря, Чаунской губы представляет собой холмистую равнину. Вдоль побережья губы тянется полоса шириной 10—15 км, подверженная засолению. При нагонах все устье реки Чаун осолоняется на 10—20 км вверх по течению, и поэтому приморские маршевые виды заходят далеко вверх по реке.
Отличительная черта побережья Чукотского моря — это цепочка кос, отделяющих от моря многочисленные лагуны. Сама береговая линия на большем своем протяжении выровненно-абразионная с аккумулятивными фрагментами. За исключением Колючинской губы на ней нет ни глубоко врезанных в сушу заливов, ни крупных мысов. В Анадырском заливе Берингова моря береговая линия, наоборот, изрезанная, с двумя крупными заливами — Креста и Анадырским лиманом. Морфогенетическое разнообразие берегов отражается на структуре и длительности существования первичных растительных сообществ приморской полосы Чукотского п-ва, между которыми в большинстве случаев нет четких границ. Формирование марша начинается обычно с поселения на илистой почве Puccinellia phryganodes, в устьевых зонах многочисленных рек выше собственно устья ее сменяет Dupontia psilosantha, менее устойчивая к засолению. Потом к пионерным видам подселяются и достигают большого обилия арктические галофиты: Carex glareosa и Arctanthemum arcticum ssp. polare (рис. 10).
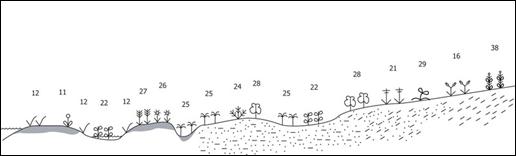 |
Рис. 10. Эколого-динамический ряд приморской маршевой растительности на илистых осушках северного побережья Чукотского п-ва (Чукотское море). Условные обозначения: приморские сообщества с доминированием: 11 — Puccinellia phryganodes, 12 — Carex subspathacea, 16 — Leymus villosissimus, 21 — Festuca rubra, 22 — Hippuris tetraphylla, 24 — Stellaria humifusa, 25 — Dupontia psilosantha, 26 — Arctanthemum arcticum ssp. polare, 27 — Calamagrostis deshampsioides, 28 — Salix ovalifolia, 29 — Carex glareosa, 38 — Rhodiola atripurpurea

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
