
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Подходы к Исследованию чувствительности модели экосистемы шельфа к вариациям ее параметров.
Институт океанологии им. РАН, МФЮА, г. Москва
Разработка математической модели морского шельфа, как решение любой крупной задачи, имеющей природоохранное значение, проводится поэтапно, по мере накопления фактических знаний о системе с помощью данных лабораторных, натурных и дистанционных наблюдений. Для того, чтобы получаемые выводы были достаточно надежными, на каждом этапе моделирования должна проводиться независимая проверка результатов расчета по материалам натурных и дистанционных наблюдений. Предложение о применении данных дистанционных наблюдений для этих целей стало возможным после создания оптического блока модели экосистемы шельфа [1] и реализации его для акваторий различных пространственно-временных масштабов и процессов.
Основными биологическими процессами, учитываемыми в модели экосистемы шельфа, являются процессы трансформации веществ при прохождении ими трофических цепей. Практически во всех работах, посвященных динамике биомассы биологических видов, последняя обычно описывается дифференциальными уравнениями 1-го порядка, правая часть которого состоит из членов, относящихся к процессам ассимиляции, дыхания, отмирания и выедания консументами более высоких трофических уровней [34]:
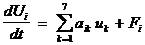 (1)
(1)
где первое слагаемое справа обусловлено внутренними взаимодействиями в системе, а Fi – влиянием внешних границ.
Весьма существенную роль в экосистеме играют процессы перехода вещества от одной компоненты к другой с образованием или трансформацией вещества этих компонент. Основной зависимостью, включаемой в модель при учете этих процессов, является зависимость скорости образования вещества соответствующей компоненты Vi от влияющих на нее факторов [1]:
![]() (2)
(2)
где Xj – концентрация j-го вещества; T- температура; L- освещенность водной среды. В ряде случаев вклад факторов может быть аддитивен и выражение для Vi представлено в виде:
![]() (3)
(3)
Модель экосистемы северо-западного шельфа Каспийского моря [2-4], составляет более высокий уровень обобщения по отношению к моделям процессов (1), поэтому конкретный вид выражений (1) для них не принципиален, а произволен, равно как и для других включаемых в них зависимостей. Имеется в виду, что конкретный вид выражений Vi может вводиться в программную систему при каждом отдельном численном эксперименте в зависимости от постановки задачи диагноза или прогноза состояния экосистемы. Основные учитываемые зависимости Vi относятся к следующим процессам:
- первичное образование биомассы при фото - и хемосинтезе;
- питание живых организмов-консументов, усвоение пищи, траты на жизнедеятельность;
- образование взвешенного и растворенного органического вещества как продуктов жизнедеятельности;
- химические реакции между веществами, протекающие физико-химическим путем.
Конкретные выражения для скоростей Vi , как правило, находятся путем обобщения данных лабораторных экспериментов. Модель, отвечающая скорости образования биомассы при фотосинтезе, может строиться на основании одного из двух биологических принципов – Либиха и Митчерлиха.
Используя предыдущий опыт моделирования для экосистем Черного, Балтийского, Охотского морей, Мексиканского залива, прибрежной акватории Гавайских островов мы считаем одним из важных этапов исследование чувствительности модели к вариациям ее параметров и выявление устойчивых диапазонов изменения ее основных констант.
Серии численных экспериментов с моделью были начаты с исследования влияния скорости фотосинтеза фитопланктона (которая определяется по принципу Либиха) на состояние экосистемы в течение года, а именно с вариаций констант ![]() - максимальных значений скорости деления клеток планктона.
- максимальных значений скорости деления клеток планктона.
При k1M= k4M= k5M=3(среднее значение из диапазона): результат расчета годового хода значений биомасс и концентраций основных компонент экосистемы (фито-, зоопланктона, макрофитов, биогенных элементов, органического вещества, нектона) представлен на рис.1. Годовой ход биомассы фитопланктона имеет два характерных максимума: весенний с максимальной биомассой 1500мг/м3. Летний с максимальной биомассой 3500 мг/м3 и почти сливающийся с летним осенний максимум с максимальной биомассой 1600 мг/м3 (рис1,а). По данным натурных наблюдений величина суммарной биомассы фитопланктона в апреле может достигать 2.5 – 3 г/м3 [5]. Для диатомовых, доля которых преобладает в суммарной биомассе это значение составляет до 2 г\м3. Конец биологической весны приходится на июнь, обусловленный формированием нового состава фитопланктона. Суммарная биомасса по наблюдениям снижается до 1,2 – 2 г/м3 [5]. Модельный расчет также выявил падение значений биомассы фитопланктона в указанный период ( рис.1,а).
С характерным для осени снижением температуры происходит снижение значений биомассы фитопланктона, по многолетним данным натурных наблюдений в октябре составляющее 1 – 2 г/м3 [5]. Рассчитанное по модели для октября значение биомассы фитопланктона 1,5 г/м3 согласуется с данными наблюдений с хорошей точностью, практически до значений самой величины биомассы ( рис. 1,а ). Такой результат расчета отражает правильно выбранный временной диапазон изменения доминирующих видов планктона – теплолюбивых и холодолюбивых.
В отличии от годового хода биомассы фитопланктона в Черном море [1] для Каспия рассчитанные летний и осенний максимумы практически слиты, не имеют между собой ярко выраженного промежутка минимума. Это связано с большим значением фотосинтетически активной радиации, поступающей на поверхность Каспия. Среднемесячное значение солнечной радиации над поверхностью Каспия в июле составляет 500 Дж\м2. Среднегодовое количество солнечной радиации над Северным Каспием по данным атласа составляет 115-130 кКал\см2 в год. А доля фотосинтетически активной радиации по данным сканирования спутниками серии NOAA в гг. в квадратах градус на градус составляет от 40 до 50% от суммарной радиации. При этом сезонный ход эффективности использования фотосинтетически активной солнечной радиации фитопланктоном Северного Каспия составляет в процентах 0.1 – зимой, 0.38 – весной, 0.63 – летом и 0.66 – осенью. При уменьшении уровня освещенности, с которого начинается ингибирования фотосинтеза светом ( в модели снижение коэффициента L1M с 2.5∙104 Лк до 0.5∙104 Лк ) расчет показывает снижение летнего максимума биомассы фитопланктона и четкое выделение летнего и осеннего максимумов, т. е. появление между ними ярко выраженного минимума.
Годовой ход биомассы зоопланктона следует за годовым ходом биомассы зоопланктона с запаздыванием по времени (рис.1,а), что отражает трофическую связь между данными компонентами. И имеет три ярко выраженных максимума: весенний с максимальной биомассой 800 мг/м3, летний - до 500 мг/м3 и осенний – до 450 мг/м3. Весенний максимум фитопланктона, приходящийся на апрель-май, для зоопланктона рассчитан в июне. Летний максимум зоопланктона рассчитан на конец августа, а незначительный осенний максимум пришелся на конец октября.
Годовой ход концентрации органического вещества имеет три выраженных максимума со значениями 3000 мг/м3 весной, 3700 мг/м3 летом и 1600 мг/м3 осенью (рис.1,б). Эти периоды соответствуют максимальным значениям биомасс фито - и зоопланктона (рис.1,а).
Для сезонного хода концентраций соединений азота и фосфора расчет показал падение значений концентраций соединений азота в сезоны интенсивного развития планктона и максимальные значения при минимумах биомассы планктона ( рис.1, в). А концентрация фосфора по расчету имеет незначительную сезонную динамику ( рис.1 в ), т. е. не следует ярко выраженным колебаниям значений биомассы планктона. Это дает основание сделать вывод о том, что соединения фосфора для Каспийского шельфа не являются лимитирующими элементами для фотосинтеза фитопланктона.
Значения биомассы макрофитов по расчету принадлежат интервалу от 50 до 200 мг/м3 (рис.1,г). А сезонная динамика нектона имеет три максимума (рис.1,д), причем последний (осенний) – наибольший; это соответствует периоду осенней путины.
Уточним, что пространственно однородная модель основных компонент является самостоятельным идеализированным объектом, которому приписаны свойства, отвечающие характерным свойствам реальной неоднородной по пространству и сложной по составу экосистемы морского шельфа. Исследование пространственно однородной модели, основанное на численных экспериментах с последующим сравнением с натурными данными, призвано дать ответ на вопрос, какие из свойств реальной системы присущи пространственно однородной системе, а какие обусловлены неоднородностью ее свойств в пространстве. В таком понимании существенный интерес представляют оптические свойства системы, поскольку такие характеристики, как спектральная яркость и цвет, могут определяться дистанционными методами с помощью спутников и аэроносителей, что обеспечивает синхронность соответствующих съемок.
Полученные результаты являются только начальным шагом в исследовании чувствительности модели к вариациям ее параметров. Следующие этапы должны быть связаны с расчетом состояния экосистемы при вариациях параметров в допустимых диапазонах. При этом необходимо соблюдать условие динамической устойчивой модели при фиксированной трофической структуре ее компонент чтобы на расчетном интервале времени выполнялось неравенство: ![]()
Список литературы:
1. Беляев морских систем. Киев: Наукова Думка, 19с.
2. , , Соловьева использование данных натурных, дистанционных наблюдений и математического моделирования для оценки состояния экологической системы Северного Каспия// Защита ОС в нефтегазовом комплексе. 2005. № 5. С. 29–39.
3. , , Соловьева использование результатов математического моделирования, дистанционных и натурных наблюдений для управления состоянием экологической системы шельфа Каспийского моря// Нефтяное хозяйство. 2005. № 6. С. 24–28.
4. , , Соловьева состояния экологической системы Каспийского шельфа на основе результатов моделирования, данных дистанционных и натурных наблюдений// Экология промышленного производства. 2006. № 1. С. 18–26.
5. , Сокольский основы стратегии защиты биологических ресурсов. Астрахань: КаспНИИРХ, 20с.
