

характер всасывания и обусловливает достоверно более низкие абсолютные значения Q (%) во всех тканях по сравнению с в/в и в/б способами введения, которые наблюдаются на протяжении только первых 4 часов при повторении тех пропорций распределения метки, которые характерны для в/в и в/б способов введения препаратов.
Соответственно времени достижения максимума накопления метки, время проявления специфичности к органам согласно значениям КДН составило 0,25 часа при в/в, 0,5 часа при в/б и 1 час при п/к способах введения. Однако уже через 2 часа от момента введения препаратов существенных достоверных различий в значениях КДН при изученных способах введения не наблюдалось.
К группе органов с высокой специфичностю накопления радиоактивного эквивалента бактериальных рибосом [классификация по Т. А. Миронюк, 1992] можно отнести грудину, тимус и лимфоузлы. Так, у грудины: при п/к введении КДН > 3 отмечался на протяжении 45 часов (93,8%) времени наблюдения; при в/в введении 44 часа (91,7%) времени наблюдения и 28 часов (58,3%) времени наблюдения при в/б способе введения препарата. У тимуса КДН > 3 отмечался на протяжении 44 часов (91,7%) времени наблюдения при п/к введении и 38 часов (79,2%) времени наблюдения при в/б способе введения препарата. Кроме того, при п/к введении рибосом КДН > 3 отмечался у лимфоузлов на протяжении 45 часов (93,8% времени) наблюдения.
К группе органов со специфическим накоплением радиоактивного эквивалента 14С-70S рибосом относятся селезёнка при всех способах введения и тимус с лимфоузлами при в/в и в/б способах введения 14С-70S рибосом. У тимуса КДН>2<3 отмечался на протяжении 46 часов (95,8%) времени наблюдения при в/в введении и 10 часов (20,8%) при в/б способе введении препарата. У лимфоузлов при в/б введении КДН>2<3 отмечался на протяжении 46,5 часов (96,9%) времени наблюдения, при в/в введении — 46 часов (95,8%) времени наблюдения. Специфическое накопление 14С-70S рибосом в тканях селезёнки было зафиксировано в течение 46 часов (95,8%) при в/б введении, в течение 45,5 часов (94,8%) при п/к введении и в течение 44 часов (91,7%) времени наблюдения при в/в способе введения препарата. При введении животным свободного 14С-лейцина КДН грудины, лимфоузлов и тимуса не превышали единицу при всех способах введения.
Для подтверждения факта активного накопления рибосомного материала в исследуемых тканях определялась экспозиция меченых веществ вычислением площади фармакокинетической кривой (ПФК). На рис. 1 представлены ПФК за весь период наблюдения. (ПФК0,25-48). Опытным и контрольным животным препараты вводились в одинаковых радиоактивных дозах. Однако суммарные значения ПФК органов при введении 14С-L-лейцина были меньше, чем при введении 14С-70S рибосом, что свидетельствует о более быстром выведении метки из организма животного в составе контроля. Независимо от способа введения препарата ПФК0,25-48 грудины свидетельствовали о максимальной кумуляции изотопа в составе рибосомного материала и достоверно отличались от остальных исследованных органов, кроме тимуса и лимфоузлов, которые занимают второе место по уровню кумуляции рибосом.
![]()
![]()
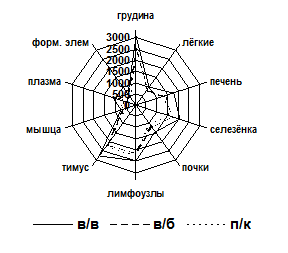
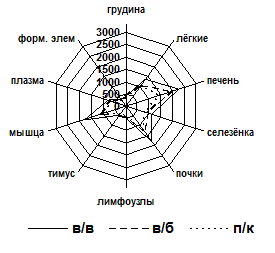
Рис. 1. ПФК экспозиции 14С-70S рибосом E. coli MRE600 (а) и 14С-лейцина (б) в органах и тканях за период от 0,25 до 48 часов
Следует отметить, что ПФК0,25-48 селезёнки не отличается от ПФК0,25-48 печени при всех способах введения.
Таким образом, характер распределения радиоактивного материала существенно не зависит от изученных способов введения бактериальных рибосом, а п/к способ может быть рекомендован в качестве предпочтительного. Вполне вероятно, что внутримышечный путь поступления рибосомного материала, не использованный нами в данной работе, также будет допустимым.
Распределение радиоактивности среди клеток крови экспериментальных животных после введения 14С-70S рибосом
Проведённые исследования показали, что при внутрибрюшинном введении 14С-70S рибосом в периферической крови мышей радиоактивность обнаруживается как во фракции эритроцитов, так и во фракции нейтрофилов. Однако, если через 0,25 часов после введения радиоактивность 106 эритроцитов составляла 123,10±19,866 Cpm, то у 106 нейтрофилов радиоактивность была в 13,55 раз больше (t=4,743, р < 0,01). Через 0,5 часа и 1 час различия не только не уменьшались, но и увеличивались (t=6,383, р < 0,005; t=7,921, р < 0,005 соответственно). Это связано с тем, что удельная радиоактивность фракции эритроцитов со временем практически не изменялась, тогда как в течение всего периода наблюдения удельная радиоактивность нейтрофилов постоянно увеличивалась. Через 0,25 часа у 106 клеток нейтрофилов было всего 1668,440±325,211 сцинтилляций в минуту, тогда как через 1 час уже 5519,110±681,950 (t=4,528, р < 0,05). Что касается сравнения с контролем (введением 14С-лейцина), то статистически значимые различия удельных радиоактивностей имелись у обеих фракций. Уровень значимости различий с контролем фракции эритроцитов в течение всего периода наблюдения оставался постоянным (р < 0,05), тогда как удельная радиоактивность фракции нейтрофилов через 0,25, 0,5 и 1 час имела отличия с контролем разного уровня значимости (t=4,812, р < 0,01; t=6,465, р < 0,005 и t=8,002, р < 0,005 соответственно). Это может говорить о продолжении взаимодействия 14С-70S рибосом и (или) их фрагментов с нейтрофилами как минимум в течении 1 часа после введения препарата в организм (время периода наблюдения).
При введении 14С-лейцина достоверной разницы между радиоактивностями фракции эритроцитов и фракции нейтрофилов не обнаружилось.
Таким образом, удельная радиоактивность фракции нейтрофильных гранулоцитов белых беспородных мышей значительно преобладает над удельной радиоактивностью других форменных элементов крови при внутрибрюшинном введении 14С-70S рибосом E. coli MRE600, но не 14С-лейцина (таблица 4).
Таблица 4
Удельная радиоактивность фракций форменных элементов периферической крови белых мышей при внутрибрюшинном введении 14С-лейцина и 14С-70S рибосом (Cpm /106 клеток) (M±m)
Препарат, вводимый животному | Исследуемые форменные элементы крови | Время после введения, ч | ||
0,25 | 0,5 | 1 | ||
14С-лейцин (контроль) | эритроциты | 57,440 ± 12,625 | 60,850 ± 9,211 | 58,370 ± 9,639 |
НГ | 101,110 ± 18,308 | 97,780 ± 14,677 | 60,830 ± 13,889 | |
14С-70S рибосомы | эритроциты | 123,10 ± 19,866*** | 131,68 ± 21,81*** | 115,82 ± 16,439*** |
НГ | 1668,440 ± 325,211** | 2942,220 ± 439,760* | 5519,110 ± 681,950* |
Примечание: уровни значимости различий по отношению к контролю:
*― p < 0,005; **― p < 0,01; ***― p < 0,05.
Иммунотропная активность 70S рибосом E. coli
в эксперименте in vivo через 1 час при однократном введении
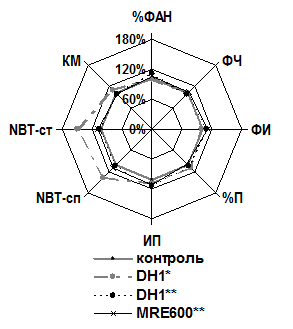
Нами была изучена фагоцитарная активность и кислородзависимый антимикробный потенциал нейтрофильных гранулоцитов мышей, которые за 1 час до исследования получали внутрибрюшинную инъекцию 70S рибосом, полученных из Hly+ и Hly– штаммов кишечных палочек.
В результате исследования выяснилось, что фагоцитарная активность НГ практически не изменялась, однако под воздействием 70S рибосом штамма E. coli DH1, полученных методом гель-фильтрации, стимулировались оксидазные микробицидные системы НГ мышей как в НСТ-спонтанном (t=3,153, р < 0,05), так и в НСТ-стимулированном (t=2,441, р < 0,05) тестах соответственно (рис. 2).
|
|
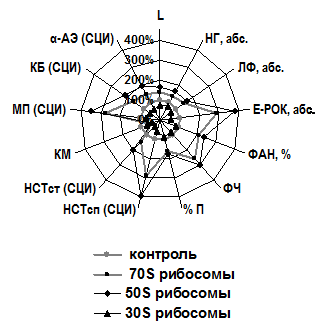
Рис. 2. Иммунотропная активность 70S рибосом E. coli штаммов MRE600 и DH1 через 1 час при однократном введении | Рис. 3. Иммунотропная активность 70S, 50S и 30S рибосом E. coli MRE600 в эксперименте in vivo при трёхкратном введении |
Иммунотропная активность70S, 50S и 30S рибосом E. coli MRE600
в эксперименте in vivo при трёхкратном введении
При трёхкратной иммунизации мышей препаратами 70S, 50S и 30S рибосом E. coli MRE600 получены эффекты, свидетельствующие о взаимосвязи иммунотропной активности рибосом с морфологическим типом рибосомных фракций. Оказалось, что иммуностимулирующая активность, которую проявляли 70S рибосомы, трансформировалась в оппозитные иммунотропные эффекты при использовании больших и малых субъединиц. При этом стимулирующая активность 50S рибосом в отношении абсолютного числа лейкоцитов, лимфоцитов и нейтрофильных гранулоцитов, а также Е-РОК-лимфоцитов была достоверно выше аналогичных показателей в группе животных, иммунизированных 70S рибосомами (t=2,704, p<0,01; t=8,243, p<0,001; t=4,376, p<0,05; t=11,678, p<0,001 соответственно). Напротив, 30S рибосомы угнетающе действовали на перечисленные выше показатели, которые достоверно отличались от соответствующих значений у контрольных животных (t=3,063, p<0,01; t=3,633, p<0,001; t=2,081, p<0,05; t=4,06, p<0,001 соответственно). Под воздействием 70S и 50S рибосом число Е-РОК возрастало в 2,8 и 3,74 раза соответственно, а под влиянием 30S рибосом снижалось в 1,8 раза, тогда как остальные анализируемые в работе показатели колебались менее существенно (в 1,3 ― 1,6 раза) для всех морфологических типов рибосом (рис. 3).
Выраженная стимуляция бактериальными рибосомами уровня НГ в периферической крови сопровождалась усилением их фагоцитарной активности и функции микробицидных систем. Под влиянием 70S и 50S рибосом фагоцитарный индекс НГ мышей возрастал в 5,7 и 7,25 раза, а индекс переваривания ― в 9,1 и 12,6 раза соответственно. Характерно, что эффекты 70S рибосом и их больших субъединиц были однонаправленными. Причём 50S рибосомы более эффективно стимулировали показатели поглотительной и переваривающей функций НГ: долю фагоцитарно-активных НГ, фагоцитарное число, фагоцитарный индекс, долю переваренных микробов и индекс переваривания (t=4,444, p<0,05; t=2,245, p<0,05; t=3,414, p<0,05; t=2,431, p<0,05; t=4,1, p<0,05 соответственно). Малые субъединицы рибосом, напротив, достоверно угнетали данные показатели по отношению к контролю (t=2,041, p<0,05; t=2,381, p<0,05; t=2,342, p<0,05; t=2,43, p<0,05; t=2,203, p<0,05 соответственно). Под воздействием 30S рибосом значения фагоцитарного индекса и индекса переваривания снижались в 1,3 и 1,6 раза соответственно.
В ходе изучения влияние рибосом на кислородзависимые микробицидные системы НГ выяснилось, что 70S рибосомы E. coli MRE600, а также их большие субъединицы достоверно стимулировали оксидазные микробицидные системы НГ мышей как в НСТ-спонтанном (t=8,886, p<0,001; t=13,563, p<0,001), так и в НСT-стимулированном (t=3,589, p<0,001; t=6,116, p<0,001) тестах соответственно. У иммунизированных 70S и 50S рибосомами животных СЦИ НГ в НСT-спонтанном тесте возрастал в 2,85 и 3,85 раза, а в НСT-стимулированном тесте ― в 1,45 и 1,98 раза соответственно по отношению к интактным животным контрольной группы. Однако значения КМ, которые достоверно (t=4,435, p<0,001; t=6,628, p<0,001) уменьшались в 1,9 и 2,2 раза соответственно, свидетельствуют о том, что 70S и 50S рибосомы при неадекватном дозировании способны вызвать истощение ресурсов оксидазных микробицидных систем НГ. Рибосомы 30S, напротив, снижали СЦИ НГ в 1,77 раза и 2,41 раза соответственно по отношению к контролю. Уровень МП НГ у мышей, иммунизированных 70S и 50S рибосомами, достоверно (t=9,573, p<0,001; t=13,598, p<0,001) возрастал в 2,7 и 3,46 раза соответственно. При этом под влиянием 50S субъединиц уровень МП был в 1,28 раза выше по сравнению с влиянием 70S рибосом (t=3,543, p<0,001). У животных, иммунизированных 30S субъединицами, СЦИ МП уменьшался в 1,48 раза (t=2,384, p<0,05).
Результаты исследования состояния кислороднезависимого микробицидного потенциала НГ мышей, иммунизированных 70S, 50S и 30S рибосомами E. coli MRE600, также подтвердили иммуностимулирующую активность 70S и 50S рибосом и депрессивное действие 30S субъединиц. Так СЦИ КБ и АЭ достоверно (t=3,461, p<0,01 и t=3,184, p<0,01) стимулировались при иммунизации 70S рибосомами, возрастая в 1,61 и 1,49 раза соответственно. Под влиянием 50S субъединиц содержание в НГ КБ и АЭ достоверно (t=5,053, p<0,001 и t=6,839, p<0,001) возрастало в 2,13 и 1,88 раза соответственно. При сравнении 70S и 50S субъединиц между собой, в очередной раз 50S рибосомы показали более выраженную стимулирующую активность как в отношении КБ (t=2,11, p<0,05), так и в отношении АЭ (t=2,416, p<0,05). Напротив, 30S субъединицы рибосом E. coli MRE600 способствовали уменьшению содержания указанных выше компонентов кислороднезависимых микробицидных систем НГ по сравнению с контролем более чем в полтора раза: СЦИ КБ ― в 1,68 раза, t=3,7, p<0,001; СЦИ АЭ ― в 1,62, t=3,719, p<0,001.
Иммунологическая специфичность препаратов бактериальных рибосом, различной степени целостности
Взаимодействие сывороточных антител с рибосомным материалом определяли в реакции двойной радиальной иммунодиффузии по [O. Ouchterlony, 1950], учитывая взаимодействие двукратных разведений полученных сывороток с препаратом рибосом в концентрации 4 мг/мл. Окрашивание гелей проводили по J. Fairbanks et al. [1971]. Через 18 часов после постановки реакции обнаруживались полосы преципитации только между сывороткой кроликов и 70S и 50S рибосомами в титре 1:8-1:16, в то время как между иммунной сывороткой, и 30S рибосомами полос преципитации не было (рис. 4).
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()


а) б) в)
Рис. 4. Реакция двойной радиальной иммунодиффузии по Ухтерлони 30S (а), 50S (б) и 70S (в) рибосом штамма E. coli MRE 600 с антисывороткой, полученной иммунизацией кролика препаратом 70S рибосом штамма E. coli MRE 600.
Рибосомы внесены в центральные лунки, двукратные разведения (от 1:2 до 1:64) антисыворотки — в периферические лунки.
Таким образом, гипериммунные кроличьи сыворотки к 70S рибосомам E. coli MRE 600 специфически взаимодействуют с 50S, но не 30S рибосомными субъединицами данного штамма. Это позволяет предположить, что антигенные детерминанты находятся на больших субъединицах и отсутствуют на малых.
ВЫВОДЫ
1. 14С-70S рибосомы E. coli MRE600 и (или) их более мелкие фрагменты, независимо от способа введения в организм, избирательно накапливаются в органах иммунной системы со следующими предпочтениями: прежде всего в грудине (костном мозге), тимусе, затем ― в лимфоузлах, а также в селезёнке.
2. Внутривенный, внутрибрюшинный и подкожный способы введения 14С-70S рибосом E. coli MRE600 белым беспородным мышам обеспечивают одинаковую тропность рибосомного материала к органам иммунной системы.
3. Удельная радиоактивность фракции нейтрофильных гранулоцитов белых беспородных мышей значительно преобладает над удельной радиоактивностью других форменных элементов крови при внутрибрюшинном введении 14С-70S рибосом E. coli MRE600, но не 14С-лейцина.
4. Гипериммунные кроличьи сыворотки к 70S рибосомам E. coli MRE600 специфически взаимодействуют с 50S, но не 30S рибосомными субъединицами данного штамма.
5. Рибосомы E. coli MRE600 70S и 50S в эксперименте обладают иммуностимулирующим эффектом, тогда как малые субъединицы 30S оказывают иммуносупрессивный эффект в отношении абсолютного количества лимфоцитов, лимфоцитов, экспрессирующих Е-рецептор и нейтрофильных гранулоцитов периферической крови нелинейных мышей.
6. Рибосомы E. coli MRE600 70S и 50S обладают иммуностимулирующим эффектом, а малые субъединицы 30S оказывают иммуносупрессивный эффект в отношении фагоцитарной активности (поглотительной и переваривающей функции), компонентов кислородзависимых (активные радикалы кислорода, миелопероксидаза) и кислороднезависимых (катионные белки, неспецифическая эстераза) микробицидных систем нейтрофильных гранулоцитов периферической крови нелинейных мышей.
7. Иммунотропная активность 70S рибосом E. coli MRE600 формируется под влиянием разнонаправленных иммунотропных эффектов большой и малой рибосомных субъединиц. Рибосомы 50S оказывают достоверно более выраженную иммуностимуляцию, чем 70S рибосомы в экспериментах на нелинейных мышах.
ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ
1. При изготовлении иммунобиологических препаратов на основе бактериальных рибосом следует использовать рибосомный материал с сохранённой структурной организацией в соответствии с ранее сформулированным принципом интактности рибосомных частиц.
2. При изготовлении иммунобиологических препаратов на основе бактериальных рибосом следует учитывать, что изолированные 50S субъединицы бактериальных рибосом оказывают иммуностимулирующий, а изолированные 30S иммуносупрессивный эффект.
Список научных работ, опубликованных по теме диссертации
1. П., Г., Ш., А., Скобликов Н. Э., В., В. Бактериальные рибосомы как новый класс иммунобиологических препаратов иммуногенного и иммунотропного действия // Медицинская наука и здравоохранение: материалы научно-практической конференции молодых учёных и студентов Краснодарского края. – Туапсе, 2002. – С.21-24.
2. В., П., Э. Фагоцитарная активность и кислородзависимый антимикробный потенциал нейтрофильных гранулоцитов мышей в условиях тахифилаксии, вызванной внутрибрюшинной инъекцией бактериальных 70S рибосом // IX Всероссийская научно-практическая конференция «Молодые учёные в медицине» посвящённая 190-летию Казанского государственного медицинского университета: тезисы докладов. – Казань, 2004. – С. 131.
3. Э., П., В. Влияние препаратов бактериальных рибосом на фагоцитарную активность и кислородзависимый антимикробный потенциал нейтрофильных гранулоцитов мышей в условиях экспериментальной тахифилаксии // Материалы V Международной научно-практической конференции «Здоровье и Образование в XXI веке» – Москва, 2004, – С. 347.
4. Кошелева C. В., П., Г. Получение 70S рибосом Escherichia coli с высоким содержанием радионуклида 14С // Материалы III региональной научно-практической конференции молодых учёных и студентов «Медицинская наука и здравоохранение». – Краснодар, 2005. – С. 21-24.
5. В., П., Э., Г. Влияние 70S, 50S и 30S рибосом Escherichia coli на функционирование системы нейтрофильных гранулоцитов и популяционный состав лейкоцитов периферической крови мышей // Материалы III региональной научно-практической конференции молодых учёных и студентов «Медицинская наука и здравоохранение». – Краснодар, 2005. – С. 24-27.
6. П., В., Э., Орлов В. Г. Иммунотропная активность 70S, 50S и 30S рибосом Escherichia coli в эксперименте in vivo // Кубанский научный медицинский вестник. – 2006. – №– С. 67-71.
7. П., В., Э., Орлов В. Г. Распределение бактериальных рибосом, меченных радиоактивным углеродом, в организме экспериментальных животных // Кубанский научный медицинский вестник. – 2006. – №10 (91), – С. 43-50.
8. В., П., Э., Г. Распределение в органах и тканях лабораторных животных радиоактивного углерода, введённого в составе 70S рибосом Escherichia coli MRE600 // Материалы Четвёртого съезда Общества биотехнологов России им. Ю. А. Овчинникова. – Пущино, 2006, – С. 116-118.
9. Крылов В. П., Кошелева С. В., Скобликов Н. Э. Распределение радиоактивности среди форменных элементов периферической крови экспериментальных животных после введения меченых 70S рибосом Escherichia coli MRE600 Материалы Четвёртого съезда Общества биотехнологов России им. . – Пущино, 2006, – С. 121-122.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 |
