

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Как видно на приведенных рисунках, даже за сравнительно короткий период времени в водохранилище заметно растут размеры зоны аноксии (содержание кислорода менее 2 мг/л на рисунке выделено черным цветом). В июле в поверхностных слоях наблюдается пересыщение воды кислородам в результате активного фотосинтеза в дневные часы.
Осеннее перемешивание приводит к выравниванию содержания растворенного кислорода по глубине и его повышению от 60-70% до 90-95% насыщения в момент замерзания водохранилищ. Зимой вновь устанавливается кислородная стратификация. В период ледостава из-за практически полного отсутствия фотосинтеза и прекращения газообмена с атмосферой концентрация в воде уменьшается и в конце зимы в придонном слое глубоководных участков оно меньше 10% насыщения. Таким образом, бескислородные условия в гиполимнионе высокопродуктивных водохранилищ достаточно типичны и для зимнего периода. Отсутствие внутриводоемных источников кислорода в зимний период позволяет ориентировочно оценить масштабы расходования кислорода на процессы деструкции органических веществ в водохранилище. Расчетами кислородного баланса в Можайском водохранилище зимой показано, что наиболее активно зимнее самоочищение воды происходит в первый месяц после образования ледяного покрова, когда расход растворенного в воде кислорода на деструкцию органического вещества может достигать 0.3-0.4 мг О2/(л×сут) (Пуклаков и др., 2002). На деструкцию органического вещества в этом водохранилище в зимний период 1983/1984 гг. было израсходовано более 500 т растворенного кислорода. В кислородном балансе глубоководных и стратифицированных водохранилищ важную роль играет продольная циркуляция, порождаемая плотностным течением, которое способствует интенсификации самоочищения придонных слоев воды и ослаблению в них дефицита кислорода. В Можайском водохранилище плотностной поток переносит до 20% массы растворенного в воде кислорода, поступающего в приплотинный район в результате внутреннего водообмена, что замедляет формирование там гипоксидной зоны.
Особенности изменения содержания растворенного кислорода по глубине в течение вегетационного периода демонстрируют данные регулярных наблюдений на русловой вертикали Красновидовского плеса Можайского водохранилища в 1999 году (рис. 4.14).

Рис. 4.14. Распределение кислорода с мая по сентябрь (по оси «х» сутки, начиная от 1 мая. По оси «у» - глубина, м) в Красновидовском плесе Можайского водохранилища в 1999 году.
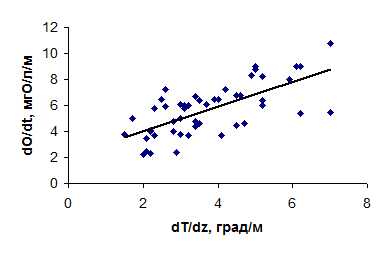
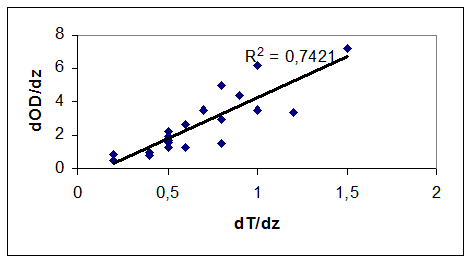
При общности экологических механизмов функционирования экосистем озер и водохранилищ существенные различия в морфологии и гидрологическом режиме последних обусловливают заметную специфику пространственно-временных изменений содержания кислорода в водохранилищах в вегетационный период. Для водохранилищ в летний период характерно клиноградное распределение кислорода по вертикали, обусловленное плотностной стратификацией. Как показывают наблюдения, между максимальными градиентами содержания кислорода и температуры существует тесная зависимость, не зависящая от природной зоны, в которой расположено водохранилище (рис. 4.15 )
 |
а)
 |
б)
Рис. 4.15. Зависимость между максимальными градиентами температуры воды и содержания растворенного кислорода в Можайском водохранилище (а) и в водохранилищах северо-востока Бразилии (б).
Пространственное положение слоя максимальных градиентов кислорода в водохранилищах почти горизонтально и в соответствие с асимметрией ложа толщина гиполимниона плавно увеличивается к плотине. Как положение термоклина и оксиклина, так и градиенты в них не остаются постоянными, и, в зависимости от синоптических условий, испытывают короткопериодные колебания, непосредственно влияющие на характер вертикального распределения кислорода.
Наиболее общие черты кислородного режима в водохранилище – образование и постепенное в течение лета заглубление термоклина и оксиклина, его относительно горизонтальное положение вдоль водоема и короткопериодные колебание градиентов, в целом сохраняются от года к году. Однако, характерные в отличие от озер колебания уровня водохранилищ как в отдельные годы, так и в течение вегетационного периода вносят определенные изменения. В многоводные годы в водохранилищах многолетнего регулирования при высоком, как правило, уровне процессы расслоения водной массы выражены менее отчетливо, градиенты в слое термоклина меньше и вероятность появления зон аноксии в гиполимнионе мала. В маловодные годы при низком уровне и особенно в условиях жаркой погоды, сопровождающей обычно маловодье, продукционно-деструкционные процессы протекают более напряженно и градиенты пространственных изменений кислорода возрастают.
В водоемах тропической зоны главным фактором кислородного режима выступают активные биологические процессы в течение всего года. Температурного и плотностного расслоения в этих полимиктических водоемах не наблюдается, но в вертикальном распределении кислорода клиноградность сохраняется и даже усиливается. Причиной столь резких изменений кислорода в этих водоемах выступает очень высокая скорость биохимических процессов вследствие высоких температур воды и высокой продуктивности водоемов. В периоды активного фотосинтеза пересыщение воды кислородом в поверхностном слое, достигающее 200% - явление обычное, также как обычным явлением можно считать и широкое распространение бескислородных зон в глубоких слоях водоемов. Отсутствие ярко выраженного плотностного расслоения и, соответственно, привычного для водоемов умеренной зоны деления водной массы на эпилимнион, металимнион и гиполимнион лишает всякого смысла использование в этих водоемах показателя скорости потребления кислорода в гиполимнионе. Вместе с тем в этих водоемах вертикальные изменения содержания кислорода хорошо отражает особенности продукционных процессов в водоемах.
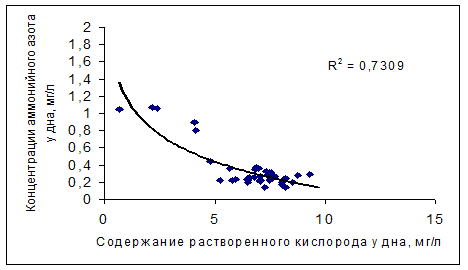
Как уже указывалось, бескислородные условия в гиполимнионе водохранилищ, кроме непосредственного влияние на особенности развития ихтиофауны водохранилищ, играют важную роль в круговороте биогенных веществ, способствуя резкому увеличению потока их минеральных форм из донных отложений, тем самым, поддерживая на высоком уровне внутреннюю биогенную нагрузку экосистем. Влияние кислородного режима на содержание биогенных веществ в москворецких водохранилищах иллюстрируют связи минеральных форм биогенных элементов (аммонийный азот и фосфор) с содержанием растворенного кислорода в придонных слоях водохранилищ, полученные по данным гидролого-гидрохимических съемок водохранилищ в летний период (рис. 4.16)
 |
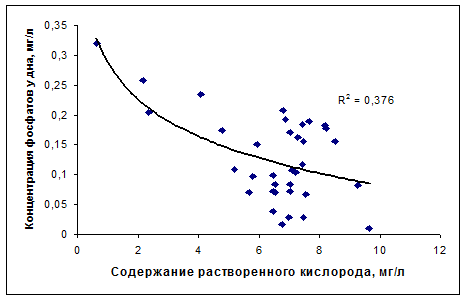 |
Рис. 4.16. Связи концентраций минеральных форм биогенных веществ с содержанием кислорода в придонных слоях москворецких водохранилищ в летний период.
Тесная связь обнаруживается между содержанием растворенного кислорода и таким важным гидрохимическим показателем как величина рН. Генетическая обусловленность этой зависимости связана с процессом фотосинтеза. Как известно, при интенсивном фотосинтезе происходит подщелачивание среды и увеличение значений рН и в тоже время растет содержание кислорода, выделяемого фитопланктоном. Анализ таких связей в москворецких водохранилищах показал, что они носят линейный характер, не зависят от синоптических условий и характеризуются высокими значениями коэффициентов корреляции.
Напряженность и изменчивость продукционно-деструкционных процессов в водоеме отражает в самом общем виде запас кислорода в водной массе, определяемый как произведение концентрации на объем водоема. По мнению , отношение запаса кислорода в водоеме через определенные промежутки времени может служить мерой устойчивости экосистемы [Якушко, Мысливец, 1989]. Резкие колебания запаса кислорода свидетельствуют о низкой устойчивости экосистемы и ее неспособности сопротивления антропогенному воздействию. Однако, использование такого показателя ограничено условиями динамической стабильности водной массы водоема в течение расчетного периода (неизменности уровня, процессов водообмена). Кроме того, само понятие «устойчивость экосистемы» пока точно не определено. Сравнение изменений запасов кислорода может быть успешно применено для ориентировочных сравнительных оценок трофического состояния близко расположенных водоемов (например, отдельных озер в озерных районах).
Как было показано в разделе 1, эффективным показателем состояния экосистемы по режиму растворенного кислорода в озере может служить кислородный индекс (ИК), определяемый как средневзвешенное значение отклонений концентраций кислорода от равновесных. Однако, использование кислородного индекса для оценки трофического состояния может быть эффективно только в стратифицированных водоемах, поскольку только при расслоении водной толщи режим и вертикальное распределение кислорода отражает уровень продукционных процессов. Расходование кислорода на деструкцию органических веществ в гиполимнионе тем больше, чем больше органики продуцируется в водоеме и поступает на дно. При наличии термоклина возникающие дефициты кислорода сохраняются и увеличиваются, поскольку подпитка гиполимниона водами верхних слоев, аэрированными кислородом воздуха, крайне затруднена. Степень этого вертикального препятствия определяется градиентом плотности воды в слое температурного скачка или вертикальной устойчивостью водной массы. Влияние стратификации на величину кислородного индекса проявляется в существовании тесной связи между ИК и величиной устойчивости водной массы, полученной нами по результатам учащенных наблюдений за распределением температуры и растворенного кислорода в летний период 2001 года на Можайском водохранилище (рис. 4.17 )
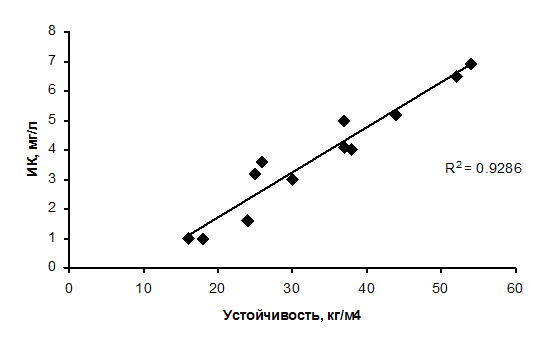
Рис. 4.17. Связь кислородного индекса с величиной устойчивости водной массы
Индекс кислорода применим не только для водоемов умеренной зоны. Цикл наблюдений за содержанием растворенного кислорода, проведенный на водохранилищах северо-востока Бразилии показал, что между ИК и наиболее простым и распространенным показателем трофического состояния – прозрачностью воды по диску Секки наблюдается тесная зависимость (рис. 4.18)
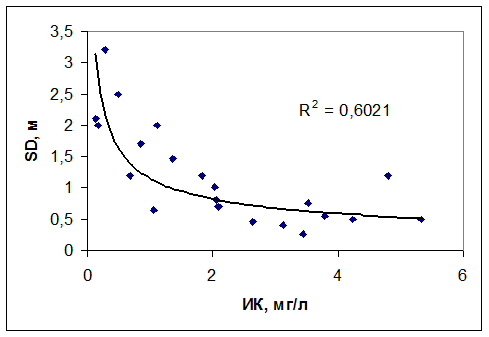
Рис. 4.18. Связь прозрачности воды с кислородным индексом для водохранилищ шт. Сеара (Бразилия).
Следует отметить, что использование осредненных значений индекса для всего водохранилища не всегда обосновано, т. к. например, в период заполнения водохранилища его верховья находятся под сильным влиянием речных вод, содержащих большое количество взвесей и характеризующихся низкой прозрачностью, не связанной с трофическими характеристиками.
4.5. Органические вещества в озерах и водохранилищах
Главные источники органических веществ в воде водоемов – приток органических веществ с речным стоком (аллохтонное органическое вещество) и продукция водных организмов (автохтонное органическое вещество). Аллохтонное вещество в водохранилищах представляет собой важный источник энергии для экосистемы [Richey, Wissmar, 1979]. Обнаруженное во многих водохранилищах превышение деструкции органического вещества над продукцией обусловлено этим избытком аллохтонного вещества.
Автохтонное органическое вещество образуется в водохранилищах в результате процессов биологического продуцирования и, в первую очередь, продуцирования фитопланктона. Из косвенных показателей органического вещества особое значение имеет определение величины БПК. Помимо потребления кислорода в процессе окисления лабильной (в основном фитопланктонной и бактериальной) органического вещества, в периоды активного фотосинтеза определенный вклад в величину БПК вносит дыхание водных организмов.
С увеличением продуктивности водоема возрастает роль автохтонной составляющей в общем балансе органического вещества водоема. Так, в близком к эвтрофному состоянию Можайском водохранилище автохтонное органическое вещество в годовом балансе в 10 раз превышает аллохтонную составляющую [Сахарова, Левшина, 1992]. Автохтонная органика относится к типу легкоразлагаемого лабильного органического вещества. Ориентировочные количественные оценки состава органического вещества водоема могут быть сделаны по соотношениям косвенных показателей [Скопинцев, 1950]. Значения отношения БПК5 / ПО (ПО – перманганатная окисляемость воды) выше 0.15 указывают на наличие значительного количества автохтонного органического вещества в водоеме, при значениях менее 0.10 в водоеме доминирует стойкое органическое вещество, в основном аллохтонного происхождения. Значение этого отношения в водохранилищах закономерно повышается в вегетационный период и минимально в зимний период. Типовой график изменений соотношения БПК и ПО во входном створе (Пестово) и в нижнем бьефе Учинского водохранилища, полученный осреднением многолетних данных () представлен на рис. 4.19.
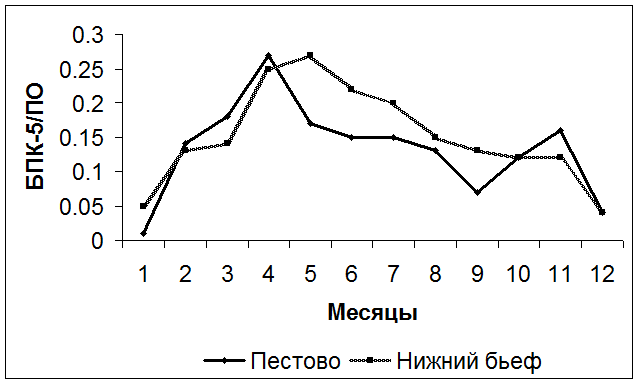
Рис. 4.19. Внутригодовой ход соотношения БПК/ПО в Учинском водохранилище.
В балансе органического вещества водоемов замедленного водообмена большую роль играет процесс седиментации. При образовании в результате фотосинтеза в верхнем фотическом слое взвешенного органического вещества (сестона), частицы которого имеют несколько больший удельный вес, чем вода, возникает вертикальный седиментационный поток. Чем больше взвешенного органического вещества продуцируется в водной массе водоема, тем более интенсивен седиментационный поток. Влияние продуктивности водоема на седиментацию органического вещества проявляется в существовании тесной зависимости между величиной первичной продукции и накоплением органического вещества на дне водоема. Линейный характер этой зависимости не изменяется для водоемов, характеризующихся различным соотношением аллохтонного и автохтонного органического вещества [Мартынова, 1988].
Процесс седиментации органического вещества в водоемах сопровождается непрерывной минерализацией его в водной толще. От соотношения интенсивности седиментации и минерализации зависит доля взвешенного органического вещества, достигающая дна водоемов. Многочисленные экспериментальный исследования, проведенные на континентальных водоемах, показывают, что доля минерализующегося в водной толще органического вещества колеблется в пределах 60-90% [Романенко, 1985]. Вертикальный поток неравномерен и находится в сильной зависимости от степени стратифицированности водоема. В термоклине скорость осаждения резко падает, а минерализация достигает максимальной интенсивности. Поскольку минерализация органического вещества приводит к регенерации биогенных элементов с последующим их включением в продукционный процесс, стратифицированность водоема может рассматриваться как важный фактор поддержания уровня первичной продукции водоема.
Наиболее надежный метод количественной оценки трансформации ОВ в водоеме замедленного водообмена - расчет баланса ОВ. Простым и в то же время достаточно информативным показателем влияния водоема на трансформацию стока ОВ может служить относительная доля изменения массы ОВ в водоеме в процессе замедления стока, или коэффициент удержания ОВ ( КRов ) (по аналогии с известным коэффициентом удержания фосфора)
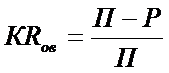
![]() где П и Р – сумма приходных и расходных составляющих баланса ОВ в водоеме.
где П и Р – сумма приходных и расходных составляющих баланса ОВ в водоеме.
Главными факторами трансформации стока ОВ как в водохранилищах, так и озерах выступает степень замедления стока, характеризуемая коэффициентом внешнего водообмена или временем пребывания воды в водоеме, и структура баланса ОВ (соотношения аллохтонной и автохтонной составляющей в приходной его части). В свою очередь структура баланса ОВ также косвенно связана с интенсивностью водообмена. Чем выше проточность водоема, чем большую роль в балансе ОВ в нем играет приток аллохтонного ОВ. Связи между этими величинами проанализированы нами с использованием данных, обобщенных М. Сакамото [1996] по некоторым наиболее изученным водоемам мира, и дополненных нами информацией о некоторых водохранилищах России (табл. 4.5).
Таблица 4.5
Характеристики органического вещества, его удержания и водообмена некоторых озер и водохранилищ мира (по [Sakamoto, 1996]).
ОВалл | ОВавт | ОВалл/ ОВавт | Кв | Hср | КRов | |
г/м2 год | год-1 | м | % | |||
ОзераБалатон (Венгрия) | 11.5 | 250 | 0.05 | 0.27 | 3.2 | |
Миррор (США) | 36 | 100 | 0.36 | 1.00 | 5.6 | 85 |
Лоуренс (США) | 50 | 342 | 0.15 | 1.33 | 5.9 | 80 |
Красное (Россия) | 84 | 269 | 0.31 | 0.87 | 6.6 | 86 |
Вингра (США) | 104 | 1200 | 0.09 | 2.09 | 2.4 | 95 |
Киннерет (Израиль) | 121 | 1312 | 0.09 | 0.21 | 24 | 99 |
Пааярви (Россия) | 126 | 174 | 1.7 | 0.30 | 14.4 | 64 |
Финдли (США) | 164 | 10 | 16.4 | 7.16 | 7.8 | 15 |
Марлон (США) | 1580 | 160 | 9.9 | 66.36 | 2.4 | 14 |
ВодохранилищаДи Грей (США) | 99 | 145 | 0.68 | 0.98 | 9.0 | 74 |
Рыбинское (Россия) | 232 | 152 | 2.28 | 1.27 | 5.6 | 74 |
Вако (Япония) | 384 | 626 | 0.61 | 5.07 | 4.0 | 65 |
Киевское (Украина) | 408 | 418 | 0.98 | 8.90 | 4.0 | 58 |
Кременчугское (Украина) | 410 | 1000 | 0.41 | 2.25 | 6.0 | - |
Иваньковское (Россия) | 917 | 340 | 2.7 | 8.30 | 3.4 | 57 |
Изабелла (США) | 2500 | 1000 | 2.5 | 9.61 | 2.6 | 36 |
Слапы (Чехия) | 3361 | 183 | 34.8 | 1.81 | 22.3 | 28 |
Можайское (Россия) | 234 | 500 | 0.47 | 1.15 | 7.5 | 38 |
Прямая связь между соотношением аллохтонного и автоохтонного органического вещества в водоемах и коэффициента водообмена обусловлена влиянием проточности на приток органического вещества (рис. 4.20). В главе 3 было показано, что продукционные характеристики водоемов, а, следовательно, образование автохтонного ОВ в водоеме, непосредственно не связаны с интенсивностью внешнего водообмена. Следует отметить, что в водохранилищах и озерах структура баланса органического вещества нередко существенно различается. В водохранилищах, как правило, приток аллохтонного органического вещества значительно выше, чем в озерах, и как следствие, в этих водоемах выше величина деструкции ОВ. Связь соотношения аллохтонного и автохтонного органического вещества с водообменом иллюстрируется рисунком, построенным по опубликованным данным. На приведенном графике отдельно показаны водохранилища, точки которых располагаются в основном выше озерных.
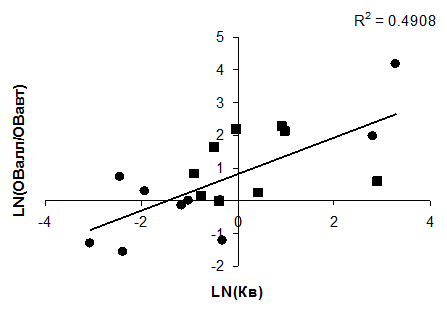
Рис. 4.20. Связь между соотношением аллохтонного и автохтонного ОВ с коэффициентом водообмена в водоемах (точки прямоугольные – водохранилища, точки круглые – озера)
Анализ колебаний самоочищающей способности водоемов от ОВ показывает, что уменьшение содержания ОВ в водоемах также связано с интенсивностью его внешнего водообмена (рис. 4.21).
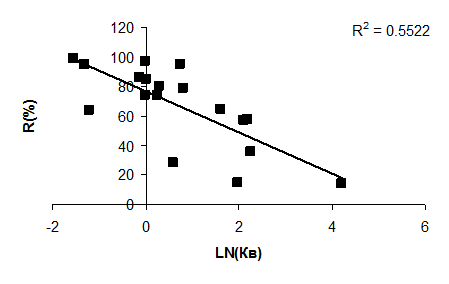
Рис. 4.21. Зависимость величины снижения содержания ОВ в водоеме от коэффициента его водообмена
В слабоводообменных водоемах более глубоко и полно происходит биоседиментация ОВ аллохтонного происхождения, усиливается физическая седиментация взвешенных ОВ. Продуцируемое в результате фотосинтеза ОВ относится к лабильному (легкоокисляемому) ОВ, большая часть которого окисляется непосредственно в водной толще, не достигая дна при осаждении [Романенко, 1985]. Эта составляющая баланса ОВ представляет собой главный энергетический резерв экосистемы водоема. Вследствие относительно быстрой трансформации органического вещества планктонного происхождения внутри водоема ее доля в сбросе из водоема оказывается небольшой. Кроме того, в высокопродуктивных водоемах интенсифицируются биохимические процессы окисления ОВ аллохтонного происхождения, поэтому такие водоемы характеризуются высокой самоочищающей способностью в отношении ОВ. Это подтверждается наличием тесной связи между соотношением автохтонной и аллохтонной органики и удержанием ОВ в водоеме (рис. 4.22).
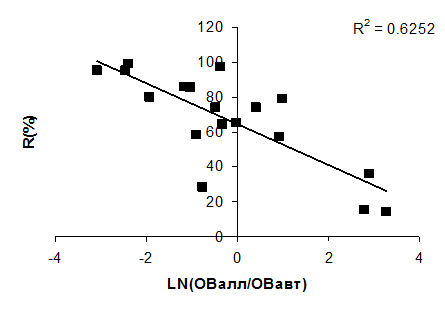
Рис. 4.22. Связь снижения ОВ в водоеме с соотношением автохтонного и аллохтонного ОВ.
Два главных фактора самоочищающей способности водоема от ОВ – интенсивность водообмена и соотношение автохтонной и аллохтонной составляющих в балансе ОВ - можно считать относительно независимыми, поскольку, как было отмечено, продукция автохтонного ОВ прямо не определяется проточностью водоема. Уравнение множественной регрессии (с коэффициентом корреляции 0.90) в этом случае имеет вид
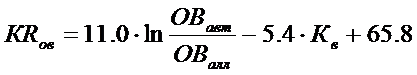
Проверка приведенного выше уравнения связи удерживающей способности водоемов по ОВ проведена нами на независимом материале по балансам ОВ различных озер и водохранилищ, не использованных при вычислении параметров зависимостей [Рыбинское…, 1972, Куйбышевское…, 1983, Экосистема…, 1989]. Результаты расчетов удержания органического вещества по приведенной выше формуле показаны в табл. 4.6.
Таблица 4.6.
Результаты расчета коэффициента удержания органического вещества в водоемах.
(В скобках – год, за который имеются данные о балансе органического вещества)
Водоем | ОВаллохт гС/м2год | ОВавтохтонн | ОВавт/ОВалл | Кв | КRфакт по балансу | КRрасч по формуле |
оз. Плещеево () | 14 | 283 | 20.2 | 0.17 | 97 | 100 |
вдхр. Куйбышевское (1960) | 822 | 169 | 4.9 | 4.2 | 70 | 61 |
вдхр. Рыбинское (1957) | 316 | 88 | 3.6 | 1.4 | 71 | 72 |
вдхр. Куйбышевское (1957) | 1105 | 271 | 4.1 | 4.2 | 63 | 59 |
Результаты расчетов по материалам, представленным в таблице 4.5, показывают, что при наличии информации о структуре приходной составляющей ОВ в водоеме можно по величине его водообмена оценить трансформацию органического вещества в нем.
Проведенный анализ характеризует влияние водоемов на трансформацию органического вещества по среднемноголетним условиям в водоемах, подверженных эвтрофированию, т. е. с высокой долей автохтонной органики. Естественно, степень этой трансформации может заметно изменяться в отдельные годы вследствие колебаний водности притоков и изменчивости внутриводоемных абиогенных и биогенных факторов первичной продуктивности.
Длительность пребывания воды в водохранилищах определяет также интенсивность окисления аллохтонных органических веществ, включающих гуминовые вещества, определяющие цветность воды. Последняя, представляя собой важный органолептический показатель качества воды, нормируемый в источниках водоснабжения и питьевых водах, оказывает также существенное влияние на оптические характеристики водных масс водоема. В этом отношении цветность вод привлекает внимание как один из факторов, определяющих толщину фотической зоны и развитие продукционных процессов.
Процессы трансформации гумусовых соединений в природных водах изучены пока явно недостаточно. Основные процессы, определяющие снижение содержания придающих воде окраску органических веществ в природных водоемах – коагуляция гумусовых соединений, соосаждение органических коллоидов с гидроокисью железа, фотохимическое окисление и, возможно, некоторую роль играет биохимическое окисление водного гумуса [Рощупко, 1970]. Важно, что все эти процессы зависят от времени пребывания воды в водохранилище, т. е. от его водообмена. Обобщение результатов наблюдений на Учинском водохранилище показали, что среднемноголетний эффект обесцвечивания вод в нем составляет 24.6±6.4%.
Осредненные для сглаживания межгодовых колебаний по пятилетним периодам значения величины снижения цветности в Учинском водохранилище сопоставлены со средними за эти же пятилетки значениями коэффициента водообмена (рис. 4.23).
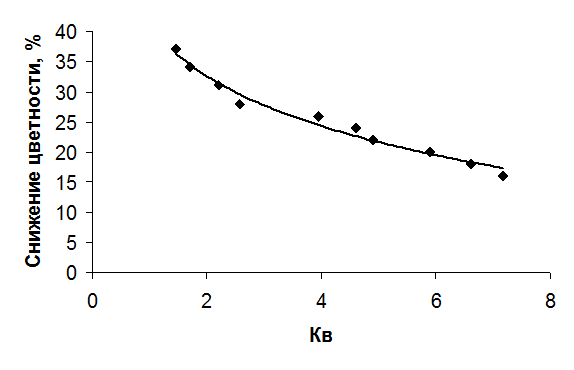
Рис. 4.23. Связь снижения цветности в Учинском водохранилище с коэффициентом его водообмена.
Очевидная закономерность снижения эффекта обесцвечивания вод с ростом проточности водохранилища хорошо аппроксимируется простой формулой [Даценко, 1987]:
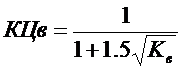
KЦв – коэффициент снижения цветности в водохранилище (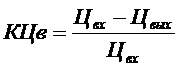 ), Кв – коэффициент водообмена водохранилища.
), Кв – коэффициент водообмена водохранилища.
Эффект снижения цветности различен в разные сезоны. Построенный типовой график снижения цветности представленный на рис.4.24, показывает, что максимальное снижение цветности наблюдается в летний период, минимальное – зимой.
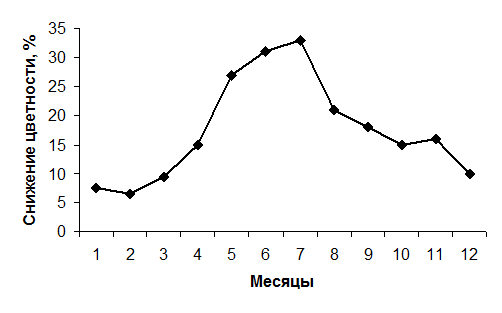
Рис. 4.24. Внутригодовое изменение величины снижения цветности в Учинском водохранилище по среднемноголетним данным.
Снижение цветности хорошо коррелирует с величиной мутности воды (коэффициент корреляции – 0.82). Это подтверждает мнение , что заполнение водохранилища мутными водами половодья резко стимулирует очистной эффект водохранилища по цветности [Францев, 1959]. По-видимому, мелкодисперсные взвешенные вещества способствуют увеличению интенсивности процессов сорбции и коагуляции окрашенных коллоидов, что усиливает эффект обесцвечивания вод. В результате закономерное для рек бассейна Верхней Волги повышение цветности в период половодья у водозаборов водопроводных станций Учинского водохранилища выражено слабо, а в некоторые годы вообще не отмечается.
Наблюдения за состоянием источников водоснабжения г. Москвы убедительно показывают, что во многих случаях водохранилища можно рассматривать как звенья технологической цепи очистки питьевой воды, осуществляющие предварительную очистку вод в результате действия природных процессов самоочищения, суммарный вектор которых направлен на улучшение качества воды.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 |
