


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
АТОПКИН Дмитрий Матвеевич
Молекулярная эволюция и систематика мышей Sylvaemus uralensis Pallas, 1811 и Apodemus agrarius Pallas, 1771 (Rodentia, Muridae)
03.00.08 - Зоология
03.00.15 - Генетика
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Владивосток
2007
![]() Работа выполнена в лаборатории Паразитологии Биолого-почвенного института ДВО РАН
Работа выполнена в лаборатории Паразитологии Биолого-почвенного института ДВО РАН
Научный руководитель:
доктор биологических наук, с. н.с
Официальные оппоненты:
доктор биологических наук, с. н.с.
доктор биологических наук, с. н.с.
Ведущая организация: Институт биологии развития им. РАН
Защита состоится "9" октября 2007 г. в 13 часов на заседании диссертационного совета Д 005.003.03 при Биолого-почвенном институте ДВО РАН г. Владивосток, проспект 100-лет Владивостоку, 159,
С диссертацией можно ознакомиться в Центральной научной библиотеке ДВО РАН
Автореферат разослан: " " сентября 2007 г.
 |
Ученый секретарь диссертационного совета,
доктор биологических наук, с. н.с.
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность. Видообразование, как одна из центральных проблем в биологии, существует не одну сотню лет. Основным достижением в данном научном направлении является возникновение теории эволюции, существовавшей изначально в виде теории естественного отбора Ч. Дарвина и преобразовавшейся в современную синтетическую теорию эволюции (СТЭ). Однако все еще остается проблема критерия вида, которая привела к кризису в современной систематике и эволюционной теории. Поэтому многие из современных исследований, подкрепленных новыми методами, направлены в основном на изучение процессов, происходящих в природных популяциях, признанных единицами эволюции, так как понимание механизмов микроэволюционных процессов может пролить свет на решение одной из основных биологических проблем: вида и видообразования. Одной из наиболее удобных групп организмов для изучения молекулярной эволюции является семейство мышевидных грызунов Muridae, в частности – широкоареальные виды, где наиболее полно могут быть отображены мультифакторные процессы становления вида. Эта особенность является одной из основных причин, по которым в качестве объекта исследования были выбраны два вида грызунов: малая лесная мышь Sylvaemus uralensis Pallas, 1811 и полевая мышь Apodemus agrarius Pallas, 1771.
Малая лесная мышь – один из широкоареальных видов грызунов, обитающих в разных высотных и зоогеографических частях, что делает его удобным для исследования закономерностей географического видообразования. Внутри вида существуют таксономические проблемы, связанные с неоднозначной интерпретацией кариологических различий популяционных групп и требующие новых, более точных и информативных методов исследования. Полевая мышь – не менее интересный вид мелких грызунов. Её ареал пролегает по всему евразийскому континенту от Центральной Европы вплоть до тихоокеанского побережья Дальнего Востока с дизъюнкцией в районе озера Байкал. Данные о популяционной генетике полевой мыши на уровне ДНК крайне скудны и ограничены локальными исследованиями. Вместе с тем такая информация необходима для решения существующих вопросов внутривидовой систематики и изучения становления современного ареала этого вида. Молекулярно-генетические исследования внутривидовой изменчивости полевой мыши только начинаются, причём наблюдается полное отсутствие данных из некоторых районов.
Цель и задачи исследования. Цель настоящей работы - исследование молекулярных механизмов эволюции на разных стадиях географического видообразования на примере широкоареальных видов мышевидных грызунов. Для этого были поставлены следующие задачи:
1. Выяснить систематическое положение трех хромосомных форм и двух географических рас малой лесной мыши Sylvaemus uralensis, а также двух географических изолятов полевой мыши Apodemus agrarius по данным маркеров ядерной и митохондриальной ДНК.
2. Провести анализ филогенетических связей малой лесной и полевой мышей с родственными видами по данным маркеров яДНК и мтДНК с таксономической интерпретацией данных.
3. С помощью маркеров яДНК и мтДНК дать оценку генетического разнообразия и характеристику популяционно-генетической структуры S. uralensis и A. agrarius.
4. Дать сравнительный анализ генетико-географической структурированности и эволюционных историй S. uralensis и A. agrarius.
Научная новизна. Впервые выполнено сравнительное исследование генетической изменчивости двух видов группы лесных и полевых мышей: Sylvaemus uralensis и Apodemus agrarius на обширной части их видовых ареалов с использованием молекулярных маркеров ядерного и митохондриального геномов. Практически все полученные в работе результаты являются новыми и приоритетными. Впервые секвенированы последовательности гена цитохрома b мтДНК трех хромосомных форм малой лесной мыши и двух изолированных популяционных группировок полевой мыши. Впервые на базе молекулярных данных проведен демографический анализ популяций и разработаны биогеографические и филогеографические концепции для малой лесной и полевой мышей. Сравнительный популяционно-генетический анализ двух видов позволил выявить особенности их разнообразия, реконструировать филогенетические связи с родственными видами, уточнить систематическое положение ряда исследованных форм. Результаты не поддерживают видовую самостоятельность S. ciscaucasicus, S. mosquensis, S. pallipes и S. kastschenkoi, предполагают конспецифичность видов S. fulvipectus и S. hermonensis, и позволяют считать географические расы S. uralensis видами на стадии становления. Малая лесная мышь из Горно-Алтайска, морфологически мышь Кащенко, имеет гибридное происхождение: по ядерным маркерам она занимает промежуточное положение между географическими расами S. uralensis, а по митохондриальным - принадлежит азиатской расе. Дифференциация двух географически изолированных группировок полевой мыши, дальневосточно-китайской и европейско-сибирско-казахстанской, укладывается в рамки внутривидовых популяционных отличий. Молекулярно-генетическими данными поддерживается выделение двух подвидовых форм: A. a. agrarius и A. a. mantchuricus.
Теоретическая и практическая значимость работы. Полученные результаты генетической изменчивости двух широкоареальных видов важны для понимания эволюционных процессов, происходящих на разных этапах видообразования. Уточнение систематического положения ряда форм западно-палеарктической группы лесных мышей является вкладом в разработку систематики и филогении группы лесных и полевых мышей. Поскольку лесные и полевые мыши широко распространены, являются основным компонентом многих биоценозов, вредителями сельского хозяйства, природными носителями инфекционных заболеваний человека и животных, исследование их внутривидовой генетической изменчивости и систематики важны для решения задач практической эпидемиологии и паразитологии.
Апробация работы. Результаты были представлены на международной научной конференции "Современные проблемы генетики" (Минск, 2005), на международной научной конференции "Эволюция жизни на Земле" (Томск, 2005) на съезде Всесоюзного Териологического Общества (Москва, 2006), на конференции-конкурсе молодых ученых Биолого-почвенного института ДВО РАН (Владивосток, 2006), на международной научной конференции "Современные проблемы биологической эволюции" (Москва, 2007), на международном симпозиуме Modern Achievements in Population, Evolutionary and Ecological Genetics (MAPEEG) (Владивосток, 2007) и на совместном заседании отделов зоологии и эволюционной биологии и генетики БПИ ДВО РАН в 2007 году.
Публикации. По теме диссертации опубликовано 6 работ.
Структура и объем работы. Диссертация состоит из введения, 3 глав, заключения, выводов, списка литературы и приложения. Список литературы включает 188 наименований, из них 130 на иностранных языках. Работа изложена на 258 страницах и содержит 44 таблицы и 74 рисунка.
Благодарности. Автор выражает глубокую признательность научному руководителю, д. б.н. за обучение и помощь в освоении молекулярно-генетических методов, методические указания и всестороннюю поддержку. Также автор искренне благодарен к. б.н. (ИБР им. РАН, Москва), к. б.н. (ИПЭЭ им. РАН) и д. б.н. (БПИ ДВО РАН) за любезно предоставленный материал, к. б.н. , к. б.н. , д. б.н. , д. б.н. , д. б.н. за обсуждение и критические замечания.
ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ
Глава 1. Обзор литературы
1.1. Видовое разнообразие, распространение и таксономическая классификация лесных и полевых мышей. В данном разделе рассмотрен вклад зоологических исследований в решение таксономических проблем лесных и полевых мышей. Кратко описаны основные таксономические системы, разработанные разными авторами для для данной группы видов, а также общая информация о биологии и географическом распространении.
1.2. Молекулярная филогенетика и систематика лесных и полевых мышей. Проанализированы основные работы по молекулярно-генетическим исследованиям межвидовой дифференциации и филогении видов в составе двух родов: Sylvaemus и Apodemus.
1.3. Внутривидовая изменчивость лесных и полевых мышей по данным молекулярных маркеров ядерной и митохондриальной ДНК. В данном разделе рассмотрены основные работы по исследованию внутривидовой генетической изменчивости лесных и полевых мышей с помощью мультилокусных маркеров ядерной ДНК и индивидуальных последовательностей участков митохондриальной ДНК.
1.4. Краткая характеристика основных молекулярно-генетических методов исследования. В разделе дана краткая характеристика таких молекулярно-генетических методов, как полимеразная цепная реакция, RAPD анализ и секвенирование последовательностей ДНК (метод химической деградации по Маскаму-Гилберту, ферментативный метод по Сангеру и ПЦР-секвенирование).
Глава 2. Материалы и методы
2.1. Сведения об использованном материале. Материалом для молекулярно-генеических исследований послужили 78 образцов пяти видов Sylvaemus: S. uralensis (n=63), S. ponticus (n=6), S. fulvipectus (n=6), S. flavicollis (n=1), S. sylvaticus (n=2) и 49 образцов полевой мыши A. agraius, собранных в 33 разных областях России и стран ближнего зарубежья. В общей сложности, включая литературные данные из базы GenBank, в анализ было включено 15 видов лесных и полевых мышей, что составило 224 образца. Материал по видам Sylvaemus любезно предоставлен к. б.н. (ИБР им. РАН). Материал по полевой мыши из европейско-сибирско-казахстанского изолята любезно предоставлен к. б.н. (ИБР им. Кольцова РАН) и (ИПЭЭ им. РАН), а из дальневосточного изолята - д. б.н. (БПИ ДВО РАН). Почти все образцы кариологически и биохимически типированы.
2.2. Выделение ДНК. ДНК лесных и полевых мышей выделяли из тканей печени и сердца, фиксированных в 80% этаноле с использованием протеиназы К и дальнейшей экстракции смесью фенол-хлороформ.
2.3. RAPD анализ ядерного генома. RAPD анализ видов лесных мышей проводился с использованием 12 олигомерных праймеров, для RAPD анализа полевой мыши использовано 10 праймеров. Статистическая обработка данных, включая филогенетические реконструкции, выполнялась с использованием ряда программ: RFLPScan 3.12; PopGen 3.2 (Yeh et al., 1999); TFPGA 1.3 (Miller, 1997); NTSYS 2.0 (Rholf, 1987); Treecon 1.3b (Van de Peer & De Wachter, 1994); Statistica 6.0.
2.4. Анализ последовательностей гена цитохрома b митохондриальной ДНК. Амплификацию и секвенирование гена цитохрома b мтДНК проводили с помощью пяти универсальных специфических праймеров (Paabo et al., 1988; Kocher et al., 1989; Irwin et al, 1991) и флуоресцентных красителей BigDye Terminator Sequencing Kit. Чтение последовательностей выполняли на капиллярном лазерном секвенаторе ABI 310 на базе БПИ ДВО РАН. Выравнивание последовательностей и дальнейшую статистическую обработку данных проводили в программах: Mega 3.1 (Kumar et al., 2005), Arlequin 3.11 (Excoffier, 2006), Modeltest 3.06, Paup 4. b.10 (Swofford, 2000).
Глава 3. Результаты и обсуждение.
3.1. Молекулярная филогенетика и систематика восточных популяций малой лесной мыши Sylvaemus uralensis. Малая лесная мышь впервые как самостоятельный вид была описана в 1952 году в Словакии (Kratochvil, Rosicky, 1952), позже была идентифицирована в Средней Азии, Восточной Европе и на Кавказе (Filipucci et al., 1992; Межжерин, Зыков, 1991; Воронцов и др. 1992; Челомина и др., 1998). Высокая гетерогенность этого вида вскоре была обнаружена по кариологическим и генетико-биохимическим данным, но таксономическая интерпретация данных у разных авторов отличалась. Одни авторы считали европейские формы "хорошими" видами (Орлов и др., 1996), другие признавали такую точку зрения необоснованной и, на основе биохимических данных, выделяли внутри вида две географические расы: азиатскую и европейскую (Богданов, 2004); в состав последней входили две хромосомные формы: восточно-европейская и южно-европейская (Богданов, 2001). Мы попытались разрешить эту проблему с помощью молекулярных маркеров ДНК.
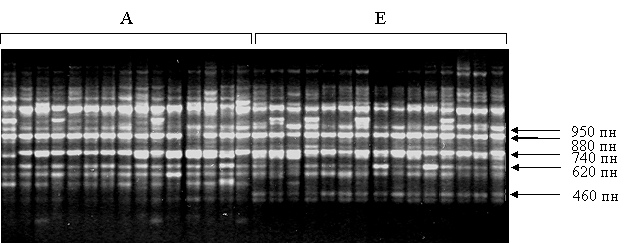 3.1.1. Генетическая изменчивость и дифференциация малой лесной мыши по данным анализа мультилокусных RAPD-маркеров. RAPD маркеры видов Sylvaemus продемонстрировали высокие видоспецифичность и внутривидовую изменчивость. У малой лесной мыши выявлено 255 локусов, 228 из которых оказались полиморфными. Между географическими расами наблюдались отличия в содержании высокочастотных локусов, а также был выявлен диагностический признак - фрагмент длиной 460 пар нуклеотидов (пн), характеризующий европейскую расу (рис. 1).
3.1.1. Генетическая изменчивость и дифференциация малой лесной мыши по данным анализа мультилокусных RAPD-маркеров. RAPD маркеры видов Sylvaemus продемонстрировали высокие видоспецифичность и внутривидовую изменчивость. У малой лесной мыши выявлено 255 локусов, 228 из которых оказались полиморфными. Между географическими расами наблюдались отличия в содержании высокочастотных локусов, а также был выявлен диагностический признак - фрагмент длиной 460 пар нуклеотидов (пн), характеризующий европейскую расу (рис. 1).
Рис. 1. RAPD-PCR спектры малой лесной мыши S. uralensis, полученные с помощью праймера OPC-05 (A - азиатская раса, Е - европейская раса)
Параметры генетической изменчивости (полиморфизм P, гетерозиготность Heun, наблюдаемое na и эффективное ne число аллелей на локус, индекс Шеннона-Вивера I) у географических рас были примерно одинаковыми, а среди хромосомных форм наиболее изменчивой оказалась южно-европейская форма. Бóльшая часть генетического разнообразия приходилась на внутрипопуляционную компоненту (HS>DST), а значение индекса генетической дифференциации (GST) между азиатской и европейской географическими расами было существенно выше (GST=0.13), чем между европейскими хромосомными формами (GST=0.07).
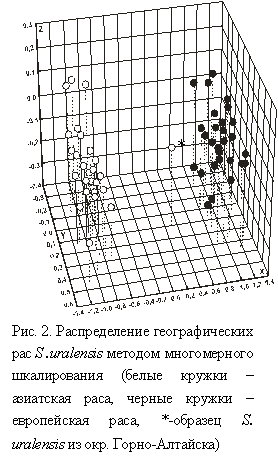 Уровень генных потоков (Nm) между европейскими хромосомными формами превышал таковые между мышами азиатской и европейской рас в 2-2.5 раза. Средние значения генетических дистанций Нея внутри географических рас и между ними оказались примерно равными (D=0.26 и 0.33, соответственно), но ниже межвидовых значений в 3-4 раза. Высокая дифференцированность малой лесной мыши на географические расы также была выявлена с помощью точного теста Фишера (p=0) и метода многомерного шкалирования (рис. 2). Интересно, что образец из Горно-Алтайска, морфологически мышь Кащенко, локализовался между расами S. uralensis, что, предполагает его гибридное происхождение, хотя данные о существовании зоны гибридизации между географическими расами S. uralensis отсутствуют. Результаты корреляционного анализа генетических параметров S. uralensis и экологических факторов среды свидетельствуют о влиянии географической широты и рельефа на генетическую структуру S. uralensis. Корреляции географических и генетических расстояний внутри азиатской расы были положительными (r=0.87), а внутри европейской расы корреляции отсутствовали (r=0.06), что предполагает разный характер расселения географических рас S. uralensis.
Уровень генных потоков (Nm) между европейскими хромосомными формами превышал таковые между мышами азиатской и европейской рас в 2-2.5 раза. Средние значения генетических дистанций Нея внутри географических рас и между ними оказались примерно равными (D=0.26 и 0.33, соответственно), но ниже межвидовых значений в 3-4 раза. Высокая дифференцированность малой лесной мыши на географические расы также была выявлена с помощью точного теста Фишера (p=0) и метода многомерного шкалирования (рис. 2). Интересно, что образец из Горно-Алтайска, морфологически мышь Кащенко, локализовался между расами S. uralensis, что, предполагает его гибридное происхождение, хотя данные о существовании зоны гибридизации между географическими расами S. uralensis отсутствуют. Результаты корреляционного анализа генетических параметров S. uralensis и экологических факторов среды свидетельствуют о влиянии географической широты и рельефа на генетическую структуру S. uralensis. Корреляции географических и генетических расстояний внутри азиатской расы были положительными (r=0.87), а внутри европейской расы корреляции отсутствовали (r=0.06), что предполагает разный характер расселения географических рас S. uralensis.
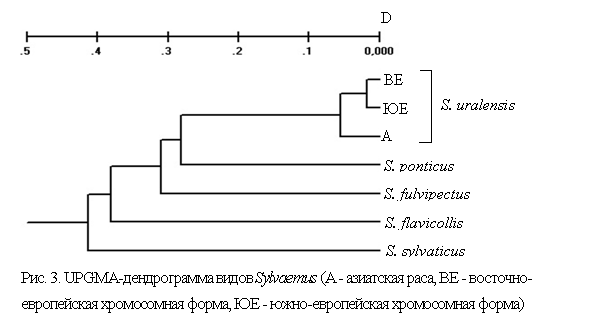
Филогенетические реконструкции дифференцируют малую лесную мышь на два основных кластера, соответствующих географическим расам. На популяционной дендрограмме UPGMA (рис. 3) идентифицируется восточно - и южно-европейская хромосомные формы S. uralensis, а азиатская раса по отношению к ним является базальной, т. е. предположительно более древней. Дифференциация малой лесной мыши на расы была также поддержана топологией минимального остовного дерева MST. Все филогенетические реконструкции указывали на высокий уровень дивергенции сравниваемых видов Sylvaemus и более высокую дифференциации географических рас S. uralensis по сравнению с дифференциацией европейских хромосомных форм (рис. 3).
3.1.2. Филогенетические связи видов и внутривидовых форм мышей рода Sylvaemus были реконструированы по данным 408 пн участка гена цитохрома b мтДНК на основе генетических дистанций, рассчитанных с помощью двупараметрической модели Кимуры (Kimura, 1980). Выбор оптимальной математической модели для расчета генетических дистанций проводили с помощью информационного критерия Акейки (AIC) (Akaike, 1974) в программе Modeltest 3.06. Расчет дистанций проводился с учетом всех типов замен во всех позициях кодонов. Величины генетических дистанций (d) между видами Sylvaemus, варьировали в широких пределах (d=5.2%-12%); значение d между европейской и азиатской расами малой лесной мыши (d=5.7%) соответствовало нижнему уровню видовой дивергенции, а различия хромосомных форм европейской расы не превысили пределов внутривидовой дифференциации других видов Sylvaemus (d=1.4%), поэтому их состоятельность в качестве видов S. mosquensis и S. ciscaucasicus, выделяемых некоторыми авторами (Орлов и др., 1996), нашими данными не поддерживается. Экземпляры форм S. pallipes и S. kastschenkoi, распространённых соответственно на Памиро-Алтае и Северном Алтае отнесены при кластерном анализе к особям азиатской расы малой лесной мыши, что противоречит существующему мнению о видовом ранге указанных форм (Межжерин, 1997). По данным RAPD анализа мы предположили, что образец S. kastschenkoi может иметь гибридное происхождение (см. раздел 3.1.1). Локализация этого образца внутри азиатской расы S. uralensis по данным мтДНК подтверждает это предположение. Возраст видов Sylvaemus оценен в 2-5 млн. лет, время формирования географичесих рас S. uralensis составило более 2 млн. лет назад.
Филогенетические реконструкции демонстрируют высокую генетическую дифференциацию видов лесных мышей рода Sylvaemus с выраженной филогеографической

Рис. 4. UPGMA-дендрограмма видов Sylvaemus (римскими цифрами обозначены основные кластеры, арабскими-значения bootstrap, %)
структурой широкоареальных таксонов. На дендрограмме UPGMA (рис. 4) выявляются 4 основных последовательно ответвляющихся кластера: I - S. sylvaticus, II - S. fulvipectus c S.hermonensis, III - S. alpicola, S. flavicollis и S.ponticus, IV - географические расы S. uralensis. Видовые кластеры, за исключением S. fulvipectus, поддерживаются высокими бутстрепп значениями (90-100% в рамках 1000 репликаций); монофилия разных видов статистически поддерживается только в двух случаях: для S. ponicus с S. flavicollis (90%) и для S. fulvipectus с S. hermonensis (100%). Уровень дифференциации между последними видами оказался очень низким (d=0.8%), что, в совокупности с вышеизложенными результатами, свидетельствует о целесообразности их сведения в синонимы с наименованием S. fulvipectus, имеющим приоритет.
3.1.3. Генетическое разнообразие, систематика и филогеография малой лесной мыши были исследованы по данным частичного (на полной выборке из 63 образцов) и полного секвенирования (на 33 образцах, представляющих все локалитеты) гена цитохрома b мтДНК. В целом популяционные группы S. uralensis отличались как по вариабельности нуклеотидных сайтов гена цитохрома b, так и по основным параметрам генетического разнообразия (генное разнообразие h, нуклеотидное разнообразие π, число попарных различий k). Генетическое разнообразие географических рас было примерно одинаковым, а среди хромосомных форм наиболее гетерогенной (как и в RAPD анализе) оказалась южно-европейская форма.
 Высокий уровень дифференциации географических рас был выявлен с помощью различных тестов, включая статистику FST и оценку генных потоков (Nm), тест на распределение разнообразия AMOVA и многомерное шкалирование. Наименее дифференцированными друг с другом оказались европейские хромосомные формы (FST=0.05-0.12), что примерно в 7 раз ниже дифференциации азиатских и европейских популяционных групп (FST=0.818-0.868). Соответственно, генные потоки между азиатской и европейской расами были ниже (Nm=0.08-0.11), чем между европейскими хромосомными формами (Nm=3.57-8.95), причём около 80% генетического разнообразия приходилось на межпопуляционную компоненту. Тест на демографическую структуру популяций предполагал быстрое расселение азиатской расы за короткий промежуток времени
Высокий уровень дифференциации географических рас был выявлен с помощью различных тестов, включая статистику FST и оценку генных потоков (Nm), тест на распределение разнообразия AMOVA и многомерное шкалирование. Наименее дифференцированными друг с другом оказались европейские хромосомные формы (FST=0.05-0.12), что примерно в 7 раз ниже дифференциации азиатских и европейских популяционных групп (FST=0.818-0.868). Соответственно, генные потоки между азиатской и европейской расами были ниже (Nm=0.08-0.11), чем между европейскими хромосомными формами (Nm=3.57-8.95), причём около 80% генетического разнообразия приходилось на межпопуляционную компоненту. Тест на демографическую структуру популяций предполагал быстрое расселение азиатской расы за короткий промежуток времени
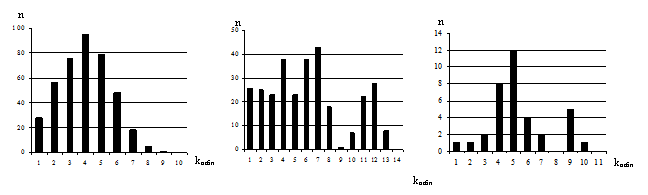
Азиатская раса Восточно-европейская хромосомная форма Южно-европейская хромосомная форма
Рис. 5. Распределение наблюдаемых попарных различий (kнабл) в хромосомных формах и географических расах S. uralensis
(унимодальное распределение попарных различий) с одной стороны, и относительную демографическую стабильность во времени европейских хромосомных форм – с другой (мультимодальное распределение попарных различий) (рис. 5). Результаты предполагали
также прохождение восточно-европейской хромосомной формой стадии «бутылочного горлышка» (выраженный пик в области 0-1 оси абсцисс). Последние демографические изменения в хромосомных формах S. uralensis , согласно полученым данным, датированы 80-100 тыс. лет назад. Эти результаты поддерживались филогенетическими реконструкциями MST. На минимальном остовном дереве азиатская раса образовала многолучевую структуру с одним центром, тогда как европейская раса была представлена более гетерогенной группой (рис. 6). Другие филогенетические реконструкции также предполагали высокий уровень дифференциации географических рас S. uralensis с высокой бутстреп-поддержкой.
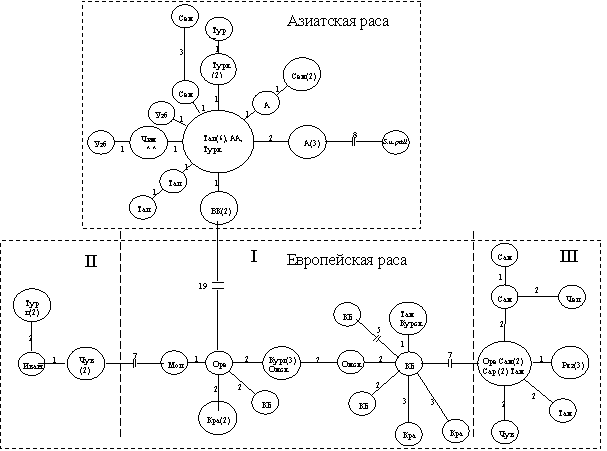 |
Рис. 6. MST для S. uralensis (данные частичного секвенирования участка гена цитохрома b мтДНК; цифрами обозначено число мутационных шагов между гаплотипами; в скобках - число особей)
Суммируя полученные данные, была разработана гипотеза становления современного ареала малой лесной мыши, включающая биогеографическую (по данным RAPD анализа) и филогеографическую (по данным секвенирование гена цитохрома b мтДНК) концепции, которые вполне согласовывались друг с другом. Гипотеза предполагала два альтернативных пути миграции S. uralensis в направлении с Юга на Север: 1. По южному побережью Каспийского моря, через восточный Иран и Туркменистан на Кавказ и далее в Европу. 2. Через южные территории Западной Сибири и Северо-Восточный Казахстан в Европейскую часть России и далее в Европу. Первый, "южный" путь представляется нам более древним. В пользу этого предположения выступают, например, повышенный уровень генетического разнообразия у южно-европейской хромосомной формы, а также данные палеонтологических исследований, доказывающих существование вполне сформированных видов Sylvaemus на территории кавказского перешейка в среднем Плейстоцене и предполагающих их плиоценовую древность (Верещагин, 1959). Поэтому есть вероятность того, что вид проникал на южные территории еще в Плиоцене. Тем не менее, принимая во внимание низкий уровень генных потоков и отсутствие «южных» генетических связей между европейской и азиатской расами S. uralensis можно заключить, что если "южный" путь когда-либо и существовал, он был менее интенсивным при распространении S. uralensis в период Плейстоцена и позднее, для поддержания контакта между расами.
Дифференциация географических рас S. uralensis, согласно нашим данным по секвенированию гена цитохрома b мтДНК, произошла более 2 млн. лет назад, возможно, в результате возникновения между ними экологического барьера в виде зоны сухого климата с сильной остепненностью местности. Эта датировка соответствует результатам палеонтологических и палеоклиматических исследований (Верещагин, 1959; Борисов, 1975; Зубаков, 1990). Экологические условия в Средней Азии и Европейской части России позволили географическим расам малой лесной мыши эволюционировать отдельно друг от друга достаточно долго в разных климатических поясах, в результате чего они достигли такой дифференциации на уровне мтДНК, которая позволяет считать их видами на стадии становления. Кроме того, в роли рефугиума для европейских малых лесных мышей в периоды похолодания мог выступать кавказский перешеек. Контакт между расами видимо не возобновлялся, несмотря на последние демографические изменения внутри каждой из них, поэтому следует ожидать формирования на исследованных территориях двух самостоятельных видов из двух географических рас малой лесной мыши.
3.2. Молекулярная филогенетика и систематика полевой мыши Apodemus agrarius из дальневосточно-китайского и европейско-сибирско-казахстанского изолятов. Полевая мышь A. agrarius распространена от Центральной Европы до окраин Дальнего Востока. Ареал этого вида состоит из двух изолированных массивов – европейско-сибирско-казахстанского (ЕСК) и дальневосточно-китайского (ДВК), разделенных евроазиатской дизъюнкцией в Забайкалье (Карасёва и др., 1992). По морфологическим признакам внутри вида на территории СНГ выделяют не более 5 подвидов (Громов, Ербаева, 1995), на хромосомном уровне показана относительная кариологическая однородность вида (Боескоров и др., 1992; Картавцева, 2002), а по биохимическим данным установлено, что ДВК более гетерогенен, чем ЕСК (Павленко, 1997). По результатам иммунологического анализа выделено 4 расы полевой мыши (Чернуха и др., 1996). Молекулярно-генетические исследования популяций этого вида ограничиваются работами на территории Корейского полуострова (Han et al., 1996; Koh et al., 1993; 2000).
 3.2.1. Генетическая изменчивость и дифференциация полевой мыши по данным анализа мультилокусных RAPD-маркеров. RAPD анализ полевой мыши выявил 258 локусов, 230 из них оказались полиморфными. Фиксированных отличий между географическими изолятами не выявлено (рис. 7), в целом у вида отмечается высокое содержание низкочастотных полиморфных локусов.
3.2.1. Генетическая изменчивость и дифференциация полевой мыши по данным анализа мультилокусных RAPD-маркеров. RAPD анализ полевой мыши выявил 258 локусов, 230 из них оказались полиморфными. Фиксированных отличий между географическими изолятами не выявлено (рис. 7), в целом у вида отмечается высокое содержание низкочастотных полиморфных локусов.
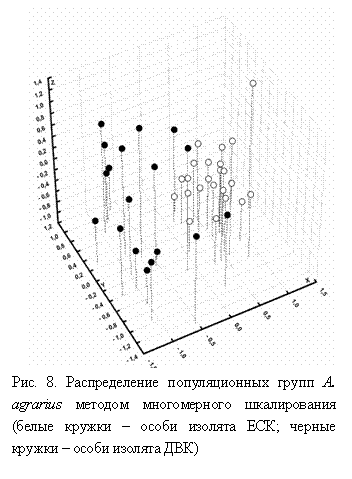 Рис. 7. RAPD-PCR спектры полевой мыши A. agrarius, полученные с помощью праймера OPC-12. ЕСК – европейско-сибирско-казахстанский изолят A. agrarius; ДВК – дальневосточный изолят A. agrarius, в. г. – внешняя группа (дорожки слева направо: A. peninsulae, S. uralensis, M. musculus, M. oeconomus, M. psilurus epsilanus)
Рис. 7. RAPD-PCR спектры полевой мыши A. agrarius, полученные с помощью праймера OPC-12. ЕСК – европейско-сибирско-казахстанский изолят A. agrarius; ДВК – дальневосточный изолят A. agrarius, в. г. – внешняя группа (дорожки слева направо: A. peninsulae, S. uralensis, M. musculus, M. oeconomus, M. psilurus epsilanus)
В пределах географических массивов полевой мыши выявлен достаточно высокий (но ниже, чем у S. uralensis) уровень изменчивости, изолят ДВК оказался более полиморфным по сравнению с ЕСК по всем генетическим параметрам. Уровень внутрипопуляционной дифференциации был выше у ДВК, а коэффициент GST - у ЕСК. Генетические дистанции как внутри популяционных групп, так и между ними отличались незначительно: Dun=0.15, 0.12 и 0.17, соответственно). Географические массивы были достоверно дифференцированы по аллельным частотам тестом Фишера (p=0); метод многомерного шкалирования на основе генетических дистанций показал их неполную дифференциацию (рис. 8).
Корреляционный анализ генетических и географических расстояний также выявил отличия между географическими изолятами A. agrarius. Результаты корреляционного анализа параметров генетической изменчивости и экологических факторов исследованных территорий предполагают высокую зависимость генетической структуры A. agrarius от лесистости и географической долготы.
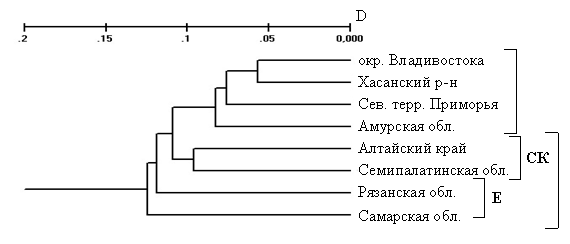
Минимальное остовное дерево предполагает более высокую гетерогенность ДВК по сравнению с ЕСК. UPGMA-дерево, построенное на основе генетических дистанций между локальными выборками полевой мыши, включающими не менее трех особей каждая, располагает их в соответствии с принадлежностью к изолятам ЕСК и ДВК (рис. 9).
 |
![]()
![]()
Рис. 9. UPGMA-дендрограмма локальных популяций A. agrarius (ЕСК - европейско-сибирско-казахстаникий изолят; ДВК - дальневосточно-китайский изолят)
3.2.2. Филогенетические связи видов и внутривидовых форм мышей рода Apodemus реконструированы по данным секвенирования 408 пн участка гена цитохрома b мтДНК на основе генетических дистанций, рассчитанных с использованием двупараметрической модели Кимуры с учетом всех типов замен во всех позициях кодонов (выбор оптимальной модели осуществлен с помощью программы Modeltest 3.06). Межвидовые генетические дистанции, варьируют в более широких пределах (d=7.7%-20%), по сравнению с видами рода Sylvaemus. Топология видов Apodemus на разных филогенетических реконструкциях сохраняется с высокой статистической поддержкой на уровне видовых кластеров. Виды группируются в две большие группы (без статистической поддержки) (рис. 10). Первая группа включает виды A. agrarius, A. chevrieri и A. speciosus, вторая - A. peninsulae, A. argenteus, A. draco и A. semotus. Единственный экземпляр A. gurkha из Непала на большинстве реконструкций образует отдельную, базальную ветвь. Время дивергенции группы видов Apodemus, согласно значениям генетических дистанций по данным мтДНК, составляет 3-8 млн. лет, а время дивергенции географических изолятов A. agrarius - около 300 тыс. лет назад. Полевая мышь A. agrarius обладает низким уровнем внутривидовой дифференциации (d=0.7%), наиболее близкой по этому критерию (среди

Рис. 10. UPGMA-дендрограмма видов Apodemus (римскими цифрами обозначены основные группы кластеров, арабскими - значения bootstrap, %)
изученных видов Apodemus) к A. peninsulae (d=0.4%). Вместе с тем, высокий уровень дифференциации выявлен между гаплотипами A. agrarius с территорий бывшего СНГ и Юго-Восточного Китая (d=2.3%). Изолят ЕСК, в отличие от ДВК, на филогенетических реконструкциях представлял собой единый кластер с низким уровнем дифференциации составляющих его гаплотипов (всего 0.3%). Гаплотипы из Южного Китая и некоторые гаплотипы Дальнего Востока России на всех филогенетических реконструкциях формировали хорошо дифференцированные базальные ветви. Эти данные подтверждают самостоятельность подвидовой формы A. a. mantchuricus, выделяемого зоологами в дальневосточной части ареала полевой мыши. Кроме того, они предполагают генетическую и таксономическую обособленность полевой мыши Южного Китая.
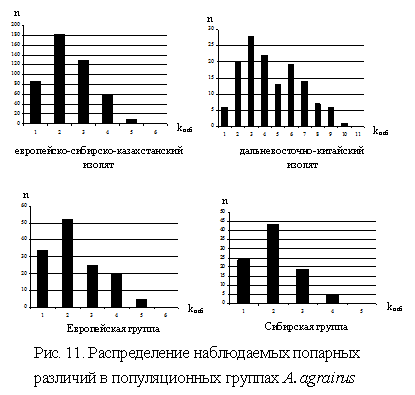
![]() 3.2.3. Генетическое разнообразие, систематика и филогеография полевой мыши были исследованы по данным частичного (на полной выборке из 48 особей) и полного секвенирования (на выборке из 18 особей, представляющих все локалитеты) последовательностей гена цитохрома b мтДНК. Значения основных параметров генетического разнообразия были выше у ДВК, по сравнению с ЕСК. Уровень дифференциации двух географических изолятов друг c другом невысокий. Значение коэффициента FST между европейской и сибирско-казахстанской популяционными группами в 2.5 раза ниже, чем между европейскими и дальневосточными популяциями (FST=0.13 и 0.31, соответственно), а уровень генных потоков – в 3 раза выше (Nm=3.24 и 1.13 соответственно). Результаты теста AMOVA предполагают, что около 83% разнообразия сосредоточено внутри географических изолятов A. agrarius. Таким образом, дифференциация между географическими изолятами A. agrarius на территории бывшего СНГ выражена слабо, а гетерогенность ДВК выше, по сравнению ЕСК, что соответствует собственным результатам RAPD анализа и литературным данным по генетической изменчивости изоферментов A. agrarius (Павленко, 1997). Методом многомерного шкалирования, с привлечением данных GenBank, выявляются три гаплогруппы мышей: европейско-сибирско-казахстанская, северная дальневосточная и южная дальневосточная (Китай, данные GenBank). Тест на демографическую структуру указывает на быструю экспансию изолята ЕСК полевой мыши за короткий промежуток времени (унимодальное распределение попарных различий) и относительно стабильное состояние ДВК (бимодальное распределение попарных различий). Кроме того, данные предполагают прохождение европейскими популяциями стадии «бутылочного горлышка» примерно 30 тыс. лет назад (рис. 11).
3.2.3. Генетическое разнообразие, систематика и филогеография полевой мыши были исследованы по данным частичного (на полной выборке из 48 особей) и полного секвенирования (на выборке из 18 особей, представляющих все локалитеты) последовательностей гена цитохрома b мтДНК. Значения основных параметров генетического разнообразия были выше у ДВК, по сравнению с ЕСК. Уровень дифференциации двух географических изолятов друг c другом невысокий. Значение коэффициента FST между европейской и сибирско-казахстанской популяционными группами в 2.5 раза ниже, чем между европейскими и дальневосточными популяциями (FST=0.13 и 0.31, соответственно), а уровень генных потоков – в 3 раза выше (Nm=3.24 и 1.13 соответственно). Результаты теста AMOVA предполагают, что около 83% разнообразия сосредоточено внутри географических изолятов A. agrarius. Таким образом, дифференциация между географическими изолятами A. agrarius на территории бывшего СНГ выражена слабо, а гетерогенность ДВК выше, по сравнению ЕСК, что соответствует собственным результатам RAPD анализа и литературным данным по генетической изменчивости изоферментов A. agrarius (Павленко, 1997). Методом многомерного шкалирования, с привлечением данных GenBank, выявляются три гаплогруппы мышей: европейско-сибирско-казахстанская, северная дальневосточная и южная дальневосточная (Китай, данные GenBank). Тест на демографическую структуру указывает на быструю экспансию изолята ЕСК полевой мыши за короткий промежуток времени (унимодальное распределение попарных различий) и относительно стабильное состояние ДВК (бимодальное распределение попарных различий). Кроме того, данные предполагают прохождение европейскими популяциями стадии «бутылочного горлышка» примерно 30 тыс. лет назад (рис. 11).
Результаты тестов на дифференциацию, включая многомерное шкалирование, в целом поддерживаются топологией минимального остовного дерева MST, причем филогеографическая структура наиболее выражена на дереве, построенном по данным полных последовательностей гена цитохрома b мтДНК (рис. 12). Выявляются три гаплогруппы, одна в составе ЕСК и две (северная и южная) в ДВК.
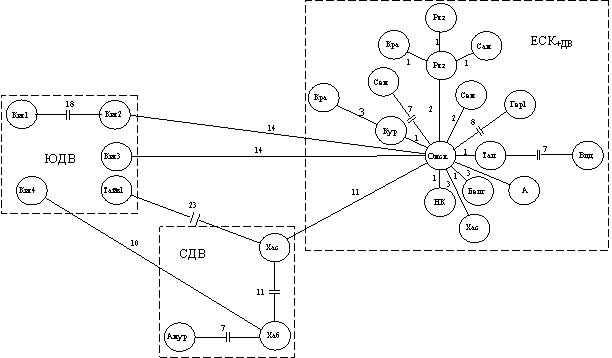 |
Рис. 12. MST для A. agrarius (данные секвенирования 1140 пн гена цитохрома b мтДНК; цифрами обозначено число мутационных шагов между гаплоипами)
Другие филогенетические реконструкции поддерживают единство ЕСК и гетерогенность ДВК, причем самыми древними признаны гаплотипы полевой мыши из Юго-Восточного Китая и Тайваня (данные GenBank). Возможно, расширение выборки (как по числу особей, так и по их географической локализации) позволит подтвердить самостоятельность таких подвидовых форм, как A. a. agrarius, A. a. karelicus, A. a. volgensis и A. a. tianschanicus. Однако на данный момент мы можем подтвердить лишь валидность A. a. mantchuricus и A. a. agrarius.
Согласно разработанным нами биогеографической (по данным RAPD анализа) и филогеографической (по данным гена цитохрома b мтДНК) концепциям, гипотетическим центром экспансии A. agrarius являются территории Юго-Восточного Китая, что соответствует литературным данным. Заселение полевой мышью евразийского континента осуществлялось в два этапа. Вначале шло заселение по двум основным направлениям: в Западную Европу и на Дальний Восток России, причем основная миграция в Западную Европу осуществлялась через южные территории Западной Сибири и Северо-Восточный Казахстан, где некоторые представители вида могли сформировать реликтовые популяции. Заселение территорий Приморья и Приамурья происходило напрямую из Китая (с Юга на Север). По нашим оценкам, распространение полевой мыши из Южного Китая началось примерно 1 млн. лет назад, когда произошла дивергенция основных линий гаплотипов. На втором, более позднем этапе, вид расселился по Северной Палеарктике, затем произошла локальная радиация полевой мыши из заселенных территорий. Между европейской и дальневосточной частями континента мог сформироваться "мост", который, однако, был неустойчив и не обеспечивал достаточных генных потоков между географическими массивами. Эти события, по нашим данным, происходили примерно 300-400 тыс. лет назад. Наиболее мощное распространение шло именно в период радиации из южных территории Западной Сибири в Северо-Восточный Казахстан в европейскую часть континента, что вполне соответствует предположениям, основанным на данных изменчивости изоферментов (Павленко, 1997). Гипотетическая схема расселения A. agrarius напоминает таковую A. peninsulae, разработанную ранее по данным секенирования 402 пн участка гена цитохрома b мтДНК (Serizawa et al., 2002), хотя датировка основных событий расселения этих двух видов имеет расхождения.
ЗАКЛЮЧЕНИЕ
В настоящей работе дана оценка генетического разнообразия малой лесной мыши S. uralensis и полевой мыши A. agrarius на обширной части их ареалов с помощью мультилокусных RAPD маркеров яДНК, а также последовательностей гена цитохрома b мтДНК. Проанализировано 476 локусов яДНК пяти видов Sylvaemus и 258 локусов полевой мыши A. agrarius. Для малой лесной мыши выявлены видо - и расоспецифические RAPD локусы. RAPD маркеров, дискриминирующих два географически изолированных массива полевой мыши, не обнаружено.
Определена полная последовательность (1140 пн) гена цитохрома b мтДНК для 51 и 408 пн фрагмента этого гена - для 126 особей пяти видов лесных мышей и полевой мыши; полученные данные зарегистрированы в базе GenBank.
Анализ межвидовых филогенетических связей позволил установить положение малой лесной и полевой мышей в системе филогенетических связей родственных видов (родов Sylvaemus и Apodemus). Малая лесная мышь монофилетична, с самой выраженной среди родственных видов филогеографической структурой. Филогеографическая структура полевой мыши среди изученных видов рода Apodemus является наименее выраженной после A. peninsulae. Несмотря на высокий эволюционный возраст A. agrarius, дифференциация его отдельных популяций, особенно в пределах европейско-сибирско-казахстанского изолята низкая, что говорит о позднем проникновении вида на эти территории.
Генетическая дифференциация популяционных комплексов (географических рас) S. uralensis соответствует географической подразделённости; дивергенция географических рас на уровне мтДНК достигла межвидовых значений (5.7%, что соответствует более 2 млн. лет раздельной эволюции) с полным завершением сортировки различных линий гаплотипов. На уровне ядерной ДНК географические расы дифференцированы на подвидовом уровне с фиксацией межрасовых отличий - RAPD маркеров. Генетическая дифференциация восточно - и южно-европейской хромосомных форм статистически недостоверна; они представляют собой общую эволюционную единицу. Для популяций A. agrairus генетическая дифференциация лишь частично соответствует географической подразделённости: изолят ЕСК изолят состоит из одной, а изолят ДВК - из двух основных дивергентных групп. Видовое единство популяционных группировок полевой мыши сомнений не вызывает; ЕСК и ДВК изоляты не имеют фиксированных отличий по локусам яДНК (RAPD маркеры) и разделяют между собой общие гаплотипы мтДНК.
Использование полноразмерных последовательностей мтДНК, по нашим данным, предпочтительно для исследования популяционных группировок с низким уровнем дифференциации, тогда как для высокодифференцированных групп достаточно использовать участки индивидуальных генных последовательностей.
Микроэволюционные процессы на уровне ядерного и митохондриального геномов имеют согласованный характер в популяциях обоих исследованных видов, что в популяционно-генетических исследованиях встречается далеко не всегда. Для сравниваемых видов разработаны хорошо согласующиеся между собой биогеографические (по данным RAPD анализа) и филогеографические (по данным гена цитохрома b мтДНК) гипотезы. Виды заселяли исследованные территории в разные геологические эпохи и под влиянием разных экологических факторов. Основной экологический фактор, влияющий на формирование характера генетической изменчивости малой лесной мыши - географическая широта, а полевой мыши - лесистость и географическая долгота. Причем корреляция этих факторов с параметрами генетической изменчивости у обоих видов прослеживается по данным как ядерного, так и митохондриального геномов.
Эволюционные истории популяций обоих видов согласовываются с теорией географического видообразования, отражая его начальный этап у A. agrairus и завершающий - у S. uralensis.
ВЫВОДЫ
1. Географические расы Sylvaemus uralensis Pallas, 1811 - виды на стадии становления; их дивергенция на уровне мтДНК (5.7%, что соответствует более 2 млн. лет раздельной эволюции) достигла межвидовых значений (с полным завершением сортировки различных линий гаплотипов), а на уровне ядерной - подвидовых (с фиксацией межрасовых отличий - RAPD маркеров).
2. Популяционные группировки Apodemus agrarius Pallas, 1711 сохранили видовое единство; европейско-сибирско-казахстанский и дальневосточно-китайский изоляты не имеют фиксированных отличий по локусам яДНК (RAPD маркеры) и разделяют между собой общие гаплотипы мтДНК.
3. Лесная мышь Алтая, морфологически мышь Кащенко, имеет гибридное происхождение: по ядерным маркерам она занимает промежуточное положение между географическими расами S. uralensis, а по митохондриальным - принадлежит азиатской расе.
4. Европейские формы S. ciscaucasicus Ognev, 1924 и S. mosquensis Ognev, 1913, и азиатские формы S. pallipes Barret-Hamilton, 1900 и S. kastschenkoi Kuznetzov, 1932 конспецифичны с S. uralensis; S. fulvipectus Ognev, 1924 и S. hermonensis Filippucci, Simson, Nevo, 1989 являются синонимами; S. ponticus Sviridenko, 1936 и S. flavicollis Melchior, 1834 - самостоятельные виды.
5. Генетическое разнообразие S. uralensis достаточно высокое, равномерно распределено между популяциями азиатской и восточно-европейской хромосомных форм; южно-европейская хромосомная форма наиболее разнообразна, что ассоциируется с ее близостью к центру внутривидовой радиации; её дифференциация с восточно-европейской формой статистически недостоверна.
6. A. agrarius генетически менее разнообразна, чем S. uralensis, изменчивость дальневосточных популяций выше, чем популяций европейско-сибирско-казахстанского изолята; принадлежность наиболее древних гаплотипов мтДНК к популяциям Южного Китая предполагает происхождение полевой мыши в Юго-Восточной Азии.
7. Заселение исследованных территорий Северо-Западной Палеарктики S. uralensis , начавшееся по максимальной оценке около 2 млн. лет назад, могло осуществляться двумя альтернативными путями ("Южным", вдоль побережья Каспийского моря и "Северным", через Северо-Восточный Казахстан); последние демографические изменения S. uralensis произошли в Плейстоцене (80-100 тыс лет назад), когда восточно-европейские популяции прошли стадию бутылочного горлышка.
8. Становление современного ареала A. agrarius началось около 1 млн лет назад и сопровождалось последующими расселением в Северной Палеарктике и локальными радиациями; в Неоплейстоцене (30-40 тыс лет назад) европейские популяции пережили событие последнего бутылочного горлышка.
9. Виды Sylvaemus (S. sylvaticus, S. flavicollis, S. ponticus, S. fulvipectus, S. alpicola и S. uralensis) высоко обособлены и монофилитичны, их эволюционный возраст составляет 2-5 млн лет. За исключением узкоареальных S. ponticus и S. alpicola, имеют выраженную филогеографическую структуру; наиболее высоко дифференцированы генетико-географические группировки S. uralensis.
10. Виды Apodemus (A. agrarius, A. peninsulae, A. argenteus, A. speciosus, A. chevrieri, A. draco и A. gurkha) высоко дивергированы и монофилетичны, их эволюционный возраст составляет 3-8 млн лет. Филогеографическая структура видов выражена в разной мере; генетически наименее структурированы среди изученных видов - A. peninsulae и A. agrarius.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
Статья, опубликованная в ведущем рецензируемом научном журнале:
1. , , Челомина изменчивость и дифференциация полевой мыши Apodemus agrarius: результаты RAPD-PCR анализа // Генетика. 2007. Т. 43. № 6. С. 804-817.
Работы, опубликованные в материалах международных конференций и симпозиумов:
2. , , Богданов разнообразие и филогенетические связи трех хромосомных форм малой лесной мыши Sylvaemus uralensis Pallas, 1811 (Rodentia, Muridae): данные RAPD-PCR анализа // Материалы III Международного симпозиума "Эволюция жизни на Земле". - Томск: ГГУ, 2005. - С. 49–51.
3. , , Богданов изменчивость полевой мыши Apodemus agrarius Pallas, 1771 на территории бывшего СССР: данные RAPD-PCR анализа // Материалы международной научной конференции "Современые проблемы генетики". - Минск: ИГЦ НАН Беларуси, 2005. - С. 69.
4. , , Богданов филогенетика и филогеография лесных мышей рода Sylvaemus по данным частичного секвенирования гена цитохрома b митохондриальной ДНК // Материалы международного совещания "Териофауна России и сопредельных территорий (VIII съезд Териологического общества)". - Москва: Товарищество научных изданий КМК, 2007. - С. 26.
5. , , Богданов процессы в популяциях малой лесной (Sylvaemus uralensis) и полевой (Apodemus agrarius) мышей: данные секвенирования гена цитохрома b мтДНК // Материалы международной конференции "Современные проблемы биологической эволюции". - Москва: ГДМ, 2007. - С. 159-161
6. Atopkin D. M., Chelomina G. N. and Bogdanov A. S. Molecular phylogeography of two widespread rodent species Sylvaemus uralensis and Apodemus agrarius based on cytochrome b gene variation // Abstr. of the Intern. Symp. MAPEEG. - Vladivostok, 2007. - P. 8.
