
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
НОСОВА
Марина Николаевна
ОНТОГЕНЕТИЧЕСКОЕ СТАНОВЛЕНИЕ КРОВЕНОСНОГО
РУСЛА В ГОЛОВНОМ МОЗГЕ И СКЕЛЕТНЫХ МЫШЦАХ У
КРЫС ЛИНИЙ ВИСТАР И НИСАГ
03.00.13 – физиология
А В Т О Р Е Ф Е Р А Т
диссертации на соискание ученой степени
кандидата биологических наук
Новосибирск 2001
Работа выполнена в Институте физиологии СО РАМН
и Институте цитологии и генетики СО РАН
Научные руководители :
д. м.н. профессор
д. б.н. профессор
Официальные оппоненты:
д. м.н. профессор
д. б.н.
Ведущая организация – Московский государственный университет
им.
Защита диссертации состоится “ ” 2001 года в часов
на заседании диссертационного совета Д 001.014.01 при Институте физиологии СО РАМН
С диссертацией можно ознакомиться в библиотеке Института физиологии СО РАМН
Автореферат разослан “ ” 2001 г
Ученый секретарь
диссертационного совета
Актуальность темы. Особенности кровоснабжения различных органов и тканей обусловлены структурным своеобразием их кровеносной системы: разными формами артериального и венозного отделов, капиллярного русла и особенностями сосудистой стенки, включая ее рецепторный аппарат, воспринимающий регуляторные сигналы. Для зрелых животных выявлены важные структурные компоненты кровеносного русла, которые определяют регионарную гемодинамику [Шошенко и др., 1982]. Однако онтогенетическое становление их остается неисследованным, хотя знание его важно, в частности, для понимания механизмов развития генетически обусловленных заболеваний сердечно-сосудистой системы и такого распространенного из них как эссенциальная артериальная гипертензия. Хорошо известно, что такая гипертензия выявляется в молодом возрасте [Якобсон и др., 1996]. У объекта наших исследований – крыс линии НИСАГ (наследственная индуцированная стрессом артериальная гипертензия) признаки ее выявляются в первый месяц постнатальной жизни [Маркель, 1985, 1991].
Цель исследования. Изучить закономерности онтогенетического формирования внутриорганной кровеносной системы у крыс Вистар и НИСАГ и выявить те характеристики ее, которые определяют особенности регионарной гемодинамики.
Для исследования ангиоархитектоники выбраны два органа – скелетная мышца и полушария головного мозга. Выбор скелетной мышцы был продиктован двумя обстоятельствами. Во-первых, скелетная мускулатура – самый большой орган у позвоночных, в том числе и у объекта наших исследований – крысы. Он составляет у нее 45% массы тела, а мышечная доля сердечного выброса – 40% в покое и до 60% при мышечной работе [Шошенко, 1975]. Онтогенетические изменения формы и гемодинамики в мышечном русле, в значительной мере, характеризуют изменения во всей кровеносной системе животного. Второе обстоятельство – это большой диапазон изменений кровотока в мышце в связи с ее функциональной потребностью в кислороде. И хотя крыса – маленькое теплокровное, у которого этот диапазон существенно меньше, чем у крупного, но по сравнению с другими органами крысы он более выражен [Шошенко, 1975; Скардс, 1984; Турганбаева и др., 1996].
Головной мозг, наоборот, сохраняет уровень объемного кровотока при функциональных нагрузках относительно постоянным [Москаленко, 1984], хотя внутри мозговых структур он изменяется, и пиальные артерии играют определяющую роль в этом перераспределении кровотока [Мчедлишвили, Барамидзе, 1994]. Головной мозг у млекопитающих интересен еще и тем, что его постнатальный рост, особенно, рост его полушарий, значительно опережает таковой скелетной мускулатуры. У крыс он практически завершается к концу первого месяца жизни, в то время как увеличение массы тела (и массы мышц) продолжается еще два и более месяцев [Дмитриева, 1971; Tamaki, 1985]. Поэтому интересно сопоставить закономерности и скорости формирования кровеносной архитектоники в двух столь разных органах и выявить влияние на них генетически обусловленной гипертензии.
Задачи исследования. 1. Провести посмертное морфометрическое исследование мозгового и мышечного кровеносных русел у крыс Вистар в возрасте 3-5 мес., 1 мес. и 2 нед.
2. Провести численное моделирование гемодинамики в мозговом и мышечном кровеносных руслах у крыс Вистар в возрасте 3-5 мес., 1 мес. и 2 нед.
3. Выявить особенности сосудистой архитектоники в мозговом и мышечном руслах у крыс линии НИСАГ в возрасте 3-5 мес., 1 мес. и 2 нед.
4. Оценить у зрелых крыс Вистар и НИСАГ чувствительность артерий мышечного типа к норадреналину (НА) и участие в этом a-адреноре-цепторов.
Научная новизна. Описано постнатальное становление мозгового русла (на примере бассейна средней мозговой артерии) у крыс Вистар и показано, что формирование типичной формы этого русла с характерной гемодинамикой происходит во второй половине первого месяца жизни. Показано становление этого русла у крыс НИСАГ и выявлены изменения, происходящие в нем в первый месяц жизни и в зрелом возрасте. Показана возрастная динамика кровеносной ангиоархитектоники в скелетной мышце (на примере двуглавой мышцы бедра) у крыс Вистар и НИСАГ. Выявлены ранние изменения ее у крыс НИСАГ, приводящие к увеличению сосудистого сопротивления, и поздние изменения, обусловленные повышенной разветвленностью русла. Выявлена повышенная спастическая реакция сосудов мышечного типа у крыс НИСАГ на иммобилизационный стресс и воздействие норадреналина. Показано, что одним из механизмов этого явления может быть повышенная чувствительность артериальных a-адре-норецепторов при стимуляции их норадреналином.
Теоретическое и практическое значение. Полученные результаты (на крысах Вистар) способствуют пониманию динамики постнатального становления внутриорганного кровеносного русла и механизмов влияния своеобразия формы этого русла на гемодинамику. Материалы, полученные на крысах НИСАГ, отчасти раскрывают структурно-функциональные механизмы повышения сосудистого сопротивления в процессе развития генетически обусловленной стресс-чувствительной артериальной гипертензии.
Положения, выносимые на защиту. 1. У зрелых крыс Вистар различие гемодинамических показателей в мозге и скелетной мышце определяется разной формой их кровеносных русел, прежде всего, степенью и локализацией расширения в направлении от магистрали к капиллярам.
2. У крыс Вистар постнатальное становление кровеносного русла в мозговых полушариях заканчивается к концу первого месяца жизни, а в скелетной мышце продолжается по мере ее онтогенетического роста.
3. У крыс НИСАГ нарушено постнатальное развитие внутриорганного кровеносного русла: в раннем возрасте это выражается, в частности, в повышенном сопротивлении мышечного русла, а в зрелом возрасте – в высокой разветвленности прекапиллярных отделов мозгового и мышечного русел и в повышенной чувствительности артериального отдела сосудистой системы к вазоконстрикторным стимулам, реализуемым через a-адрено-рецепторы
Апробация работы. По материалам диссертации сделаны доклады на II и III съездах физиологов Сибири и Дальнего Востока, Новосибирск, 1995 и 1997; международной конференции по микроциркуляции, Ярославль, 1997; XVII съезде физиологов России, Ростов-на-Дону, 1998, и на симпозиуме по регуляции мозгового кровообращения в норме и патологии, Ростов-на-Дону, 1999.
Структура и объем диссертации. Диссертация включает введение, четыре главы – обзор литературы, материалы и методы, результаты исследований и обсуждение, после которых следуют выводы, список цитируемой литературы (источников) и приложение. Работа изложена на 175 страницах машинописного текста, содержит 25 таблиц и 31 рисунок.
Материалы и методы
В работе использовали самцов нормотензивных крыс Вистар и гипертензивной линии НИСАГ в возрасте 2 нед, 1 и 3-5 мес. Линия НИСАГ получена в ИЦиГ СО РАН путем селекции на основе аутбредной линии Вистар [Маркель, 1985].
Сосудистую архитектонику изучали методом, разработанным в лаборатории микроциркуляции Института физиологии СО РАМН [Шошенко и др., 1982]. Он включает два этапа: 1. Измерение на посмертных препаратах параметров артериальных и венозных разветвлений, начиная от крупных внутриорганных магистралей к капиллярам, и построение зависимости этих параметров от диаметра стволов; морфометрические измерения вели под микроскопом МБИ-3 с помощью окулярной линейки-транспортира и окулярной сетки. 2. На основании морфометрических данных проводили численное построение кровеносного русла от артерий к венам и моделирование в нем кровотока (с использованием уравнения Пуазейля) для определения гемодинамических параметров; в расчетах принимали скорость кровотока в капилляре равной 1 мм/с, диаметр капилляра – 7 мкм и вязкость крови – 2,35 сП. Статистическую обработку и гемодинамическое моделирование проводили на ЭВМ по программам и и с помощью .
Систолическое АД измеряли у крыс под эфирным рауш-наркозом на хвосте сфигмографическим методом. Влияние иммобилозационного стресса на АД и ЧСС изучали у бодрствующих животных после ежедневного помещения их в тесную клетку на 1 ч. За сутки до опыта животным под нембуталовым наркозом (40 мкг/кг) имплантировали катетер в брюшную аорту. Изменения АД проводили электроманометром СР-1 (США) в первые (стресс-1) и третьи (стресс-3) сутки. Действие НА на АД и ЧСС исследовали у наркотизированных крыс (нембутал 50 мг/кг), используя для этого канюлю в вентральной хвостовой артерии. Измерение среднего АД (электроманометр Statkam P32) и ЧСС (ритмокардиометр) проводили в течении 10 мин с интервалом 30 сек после внутриартериального введения НА в дозе 2 мкг/кг.
Чувствительность сосудов мышечного типа к НА изучали in vitro на сегменте изолированной хвостовой артерии по изменению потока через него до и после введения НА в перфузат в дозе 1 мкг/дл (методика описана ранее, Осадчук, Маркель, 1976).
Синтез фосфатидилинозитолфосфатов (ФИФ) исследовали in vitro в изолированном сегменте брюшной аорты по интенсивности включения 32Р. Гомогенезацию ткани и экстракцию фосфолипидов проводили по методике Downes, Michell (1985) и Schacht (1981). Фосфатидилинозитолы разделяли методом тонкослойной хроматографии.
Статистическую обработку материала проводили с использованием t-критерия Стъюдента. Для сравнения достоверности различий между группами и между опытами “стресс-1”и “стресс-3” использовали тест Mann-Whitney, а внутри каждой группы – критерий Вилкоксона.
Исследование онтогенетического становления мозгового и мышечного русел крыс Вистар и НИСАГ проводилось лично автором, который пользовался советами к. б.н. . Показатели АД и ЧСС в условиях иммобилизационного стресса и внутриартериального введения НА измерялись совместно с дипломницей НГУ . Чувствительность артерий мышечного типа исследовалась совместно с аспиранткой ИЦиГ СО РАН . Синтез фосфатидилинозитолфосфатов изучала совместно с , и .
Результаты и их обсуждение
1. В мозговом русле разновозрастных крыс Вистар изменения большинства параметров сосудистых разветвлений от магистральных артерии и вены к капиллярам однотипны: увеличиваются длины сосудов L (падает их относительная длина L/D0, D0 – диаметр ствола), растет угол разветвления j0, а сами разветвления становятся более симметричными по доли сечения ветвей, H1,2=D1,22/D12+D22, где D1 и D2 диаметры ветвей) и углам их поворота (j1 и j2, j0=j1 + j2). В венозном отделе эти изменения менее выражены. С возрастом несколько увеличиваются L и L/D0, особенно в венозном русле. Артериальный коэффициент ветвления k, показывающий изменение просвета русла при переходе от ствола к ветвям и равный (D12+D22)/ D02, у 2-недельных крыс почти независим от D0 и имеет небольшую величину (около 110%). К месячному возрасту он приобретает куполообразную форму с наибольшей величиной около 130% в бифуркациях с D0 30-10 мкм.
Средняя длина капилляра в мозговой коре у 2-недельных и зрелых крыс равняется 170-160 мкм, у месячных крыс она в 1,4 раза больше. Общая протяженность капилляров в 1 мм3 коры увеличивается к месячному возрасту в 1,5 раза, после чего изменяется мало.

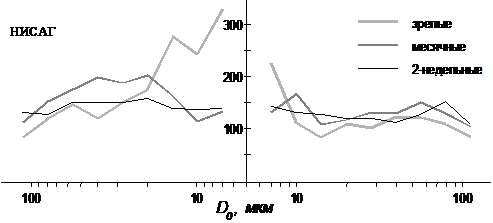
Рис. 1. Прирост числа сосудов в бассейне средней мозговой
артерии при двукратном уменьшении площади сечения сосуда
у разновозрастных крыс разных линий (расчетные данные)
Моделирование показывает (рис. 1), что к месячному возрасту мозговое артериальное русло в области стволов с D0 30-10 мкм резко разветвляется (расширяется), что приводит к возрастанию числа конечных сосудов (артериальных концов капилляров) в бассейне, что должно сопровождаться
увеличением числа венозных путей оттока (табл. 1). При этом растет (рис. 2) перепад от артериальной магистрали к капилляру линейной скорости кровотока V и потери гидравлического давления DP, особенно в этой части русла. С месячного возраста в мозговом русле формируется куполообразная зависимость пристеночного напряжения сдвига (t=8hV/D) с максимальной величиной t в области артерий D 35-20 мкм. Потери P на пути от артериальной магистрали к венозной у месячных и зрелых крыс выше, чем у 2-недельных (при условии одинаковой V в капиллярах), хотя длина пути у всех крыс примерно равна. Сосудистое сопротивление мозгового русла с возрастом падает.

Рис. 2. Параметры гемодинамики в бассейне средней мозговой артерии
А – артерии, В – вены
2. В мозговом русле крыс НИСАГ разного возраста изменения таких параметров сосудистых разветвлений как L, L/D0, H и j схожи с изменением их у одновозрастных крыс Вистар. Похожа у них и конструкция капиллярного русла: средняя длина капилляра у 2-недельных и зрелых крыс около 160 мкм, у месячных она выше на 20%; общая протяженность капилляров в 1 мм3 коры с возрастом растет до тех же величин, что у крыс Вистар, особенно к первому месяцу жизни. Основное различие касается коэффициента ветвления k. Если у крыс Вистар к первому месяцу постнатальной жизни в артериальной русле формируется куполообразная зависимость k(D0), и она сохраняется у них в зрелом возрасте, то у месячных крыс НИСАГ наблюдается похожая зависимость, но очень невыраженная (k – более низкий) и сдвинутая к более крупным пиальным артериям, однако у зрелых животных ее уже нет. У этих животных наиболее высокие k локализуются в прекапиллярных разветвлениях. Кроме того, у всех крыс НИСАГ в отличии от крыс Вистар посткапиллярные разветвления характеризуются более высокими k.
Моделирование показывает, что у крыс НИСАГ иная зависимость k от D0 отражает отличную от крыс Вистар форму кровеносного дерева (рис. 1). Это, в свою очередь, сказывается на характере артерио-венозного профиля гемодинамических показателей (табл. 1, рис. 2). У месячных крыс НИСАГ перепад V меньше в артериальном русле и больше в венозном. Поэтому соотношение числа вен к числу артерий одного D у них меньше. У этих крыс существенно ниже потери P в артериальном русле (при условии одинаковой V в капиллярах у тех и других крыс), что должно приводить к более высокому внутрикапиллярному гидростатическому давлению. У них отсутствует куполообразная зависимость t(D0), а наибольшие величины t локализованы в капилляре и около него.
У зрелых крыс НИСАГ повышена разветвленность околокапиллярных отделов, особенно с артериальной стороны. Если вновь образованные ветви (или часть их) дают начало конечным капиллярным концам, а не обра-
Рис. 3. Схема образования в русле | зуют сети (рис. 3), то артерио-венозный профиль гемодинамических показателей становится похож на таковой у крыс Вистар (рис. 2). Сосудистое сопротивление мозгового русла у крыс НИСАГ падает с возрастом, причем по сравнению с одновозрастными крысами Вистар, в 2-недельном возрасте оно несколько выше, а в зрелом – ниже (табл. 1). |
3. В двуглавой мышце бедра изменения параметров сосудистых разветвлений в направлении от магистралей к капиллярам у разновозрастных крыс Вистар однотипны: увеличиваются (больше в артериальном русле) k, L/D0 (L падает), H2 (H1 растет), j0, и разветвления становятся более симметричными по H и j. По мере роста крысы и увеличения диаметра мышечного волокна (в среднем, с 13 мкм до 49 мкм) падает в 5,3 раза плотность капилляров (с 3282/мм2 до 621/мм2), растет в 3 раза число капилляров, окружающих одно волокно (с 0,41 до 1,2) и увеличивается средняя длина одного капилляра с 191 мкм до 378 мкм.
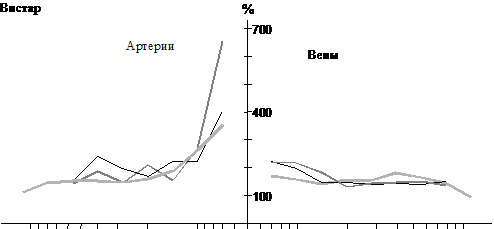
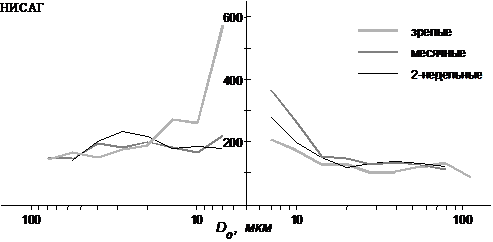
Рис. 4. Прирост числа сосудов в бассейне артерии двуглавой мышцы бедра
при двукратном уменьшении площади сечения сосуда у разновозрастных крыс
разных линий (расчетные данные)
Моделирование показывает, что для мышечного русла характерна высокая разветвленность артериальных стволов с D<20 мкм, больше у молодых крыс (рис. 4). В артериальном отделе, особенно в прекапиллярной части наблюдается значительное снижение V и P, более выраженное у молодых крыс (рис. 5). Нарастание V от капилляров к венам не столь значительно, поэтому число вен превышает число артерий того же диаметра; с возрастом это различие становится меньше (табл. 1). Зависимость t(D0) у месячных и зрелых крыс имеет куполообразную форму с максимальным t в артериальных сосудах диаметром около 20 мкм. У 2-недельных крысят высокая величина t сохраняется во всех артериальных сосудах с D>10 мкм. Сопротивление мышечного бассейна увеличивается с возрастом.

Рис. 5. Параметры гемодинамики в бассейне артерии двуглавой мышцы бедра крыс разных линий, А – артерии, В – вены
4. Характер изменений L, L/D0, H и j от артериальных и венозных магистралей к капиллярам у одновозрастных крыс НИСАГ и Вистар в двуглавой мышце бедра схожий. Существенно не различаются у них и параметры капиллярного русла. Однако зависимость k(D0) у этих двух линий крыс различна, что, согласно расчетам, отражает различие в разветвленности кровеносного дерева. У молодых крыс НИСАГ она в прекапиллярных участках значительно ниже, а в посткапиллярных – выше (рис. 4). Поэтому у этих крыс по сравнению с одновозрастными крысами Вистар меньше артериальный перепад V и P, выше сосудистое сопротивление, а наиболее высокая величина t локализована в венозном отделе (рис. 5, табл. 1). У зрелых крыс НИСАГ разветвленность артериальных стволов с D<30 мкм резко возрастает и становится выше, чем у крыс Вистар, снижается разветвленность посткапиллярных стволов. В связи с этим перепад V и P в артериях значительно увеличивается, а в венах несколько падает, t(D0) приобретает похожую на крыс Вистар форму с наибольшей величиной в артериях с D около 20 мкм; сопротивление русла снижается (при условии, что вновь образованные ветви или часть их не замыкается в сети, рис. 3).
5. Различие форм кровеносного русла в мозге и мышце у крыс Вистар, которое иллюстрируют данные разделов 1 и 3, создает предпосылки к особенностям регуляции мышечного и мозгового кровотока. Моделирование показывает, что одинаковое снижение среднегеометрического k в артериальном русле за счет снижения его в участках с наибольшей величиной (в мышце – прекапиллярные сосуды, а в мозге – мелкие пиальные артерии) повышение сопротивления и падение числа функционирующих капилляров в мышце выражено существенно больше, чем в мозге (табл. 2).
Таблица 2
Параметры длинного пути от артерии D 150 мкм до вены
в бассейне средней мозговой артерии и артерии двуглавой мышцы бедра
у зрелых крыс Вистар до и после (®) сужения артериальных сосудов
(результаты численного моделирования)
Параметр | Мозг | Мышца | ||
артерии | вены | артерии | вены | |
Число генерации: | 39®36 | 28®28 | 34®34 | 42®42 |
Средний k в русле,%: | 116®%) | 106®106 | 120®%) | 114®114 |
Средний H2 в русле,% | 26®23 (88%) | 26®24 | 30®27 (90%) | 24®24 |
Число капиллярных концов | 4354®1%) | 901®901 | 8817®3%) | 3931®3931 |
Соотношение одного диаметра | 4,5®2,0 | 2,2®0,8 | ||
Длина пути, мм | 10,5®11,0 | 11,7®11,7 | ||
Сопротивление, | 14®22 (157%) | 9®19 (211%) |
6. Приведенный в разделе 2 и 4 материал показывает, что у 2-недельных и месячных крыс НИСАГ по сравнению с одновозрастными крысами Вистар кровеносная система в головном мозге и, особенно, в скелетной мышце обладает повышенным сопротивлением. Однако у зрелых крыс НИСАГ наоборот, мы наблюдаем в тех же органах весьма разветвленное артериальное русло, которое, согласно расчетам, должно обладать низким сопротивлением. Возможно, новообразованные ветви или часть их сливается, не меняя тем самым сосудистое сопротивление (рис. 3). В тоже время, наряду
| с этим явлением у зрелых крыс НИСАГ изменяется чувствительность и реактивность сердечно-сосуди-стой системы, что обуславливает их гипертензивный статус. Специально проведенные нами опыты показали, что у зрелых крыс НИСАГ во время иммобилизационного стресса системное АД повышалось на 20-30 мм рт. ст., а ЧСС дос- |
товерно увеличивалась на 150 и более ударов в минуту. Зрелые крысы Вистар на такую стимуляцию реагировали существенно меньшим и быстро проходящим учащением пульса и, как правило, недостоверным повышением АД.
7. У зрелых крыс НИСАГ при введении НА – одного из ведущих медиаторов стресса, прирост среднего АД составляет 30-40 мм рт. ст., как и у крыс Вистар, хотя исходный уровень его существенно отличался (на 32 мм рт. ст.) (рис. 6). В опытах in vitro показано, что поток перфузата через хвостовую артерию при добавлении в него норадреналина уменьшается у крыс НИСАГ в 3,3 раза, а у крыс Вистар всего в 1,3 раза.
Таблица 3
Влияние норадреналина (НА) на синтез фосфоинозитолов в стенке
изолированного in vitro сегмента брюшной аорты крыс
Вистар и НИСАГ
Вторичные посредники(усл. ед.) | Вистар (n=5) | НИСАГ (n=5) | ||
контроль | НА | контроль | НА | |
Фосфатидилинозитол (ФИ) | 45±7 | 50±7 | 90±15 | 120±20 |
Фосфатидилинозитолмонофосфат (ФИФ) | 5±1 | 6±1 | 4±0.5 | 50±5* |
Фосфатидилинозитолдифосфат (ФИФ2) | 10±2 | 15±3 | 7±1 | 60±7* |
Показано различие с контрольной группой при p<0,001*.
В основе повышения сосудистой реактивности могут лежать разные механизмы. В симпатической регуляции тонуса сосудов принимают участие a1-, a2- и b-адренорецепторы гладкомышечных клеток. Взаимодействие a-адренорецепторов со специфическим лигандом приводит к вазоконстрикции, обусловленной активацией системы вторичных посредников (табл. 3): фосфатидилинозитолдифосфат (ФИФ2) – специфичной фосфолипазы С, высвобождением диацилглицерола и инозитолтрифосфата и увеличением внутриклеточной концентрации Ca2+ [Voigt, 1990]. Это, вероятно, является одним из механизмов, обеспечивающих повышение АД в условиях эмоционального стресса, сопровождающегося усиленным выбросом катехоламинов.
Выводы
1. Кровеносное русло зрелых крыс Вистар в мозговых полушариях (бассейн средней мозговой артерии) и в скелетных мышцах (двуглавая мышца бедра) имеет структурные и функциональные органоспецифические черты. В условиях атонии русло скелетной мышцы – его артериальная, и в меньшей степени, венозная части – более разветвлено, особенно в околокапиллярной области; в то время как в мозге повышенная разветвленность, не столь выраженная, характерна для мелких пиальных артерий.
2. Численное моделирование показывает, что у зрелых крыс Вистар а) в мышечном русле по сравнению с мозговым больше перепад линейной скорости кровотока и напорного давления от магистрали к капиллярам, ближе к прекапиллярам располагается максимум пристеночного напряжения сдвига, меньше соотношение числа вен к артериям одного калибра и ниже сопротивление кровотоку; б) сужение сосудов в области наибольшей разветвленности артериального дерева оказывает меньшее влияние на мозговой бассейн, чем на мышечный: в мозге менее выражено снижение числа функционирующих капилляров и меньше повышается сосудистое сопротивление.
3. У крыс Вистар своеобразие мозгового русла, обусловленное в основном, повышенной разветвленностью мелких пиальных артерий, формируется к месячному возрасту. Своеобразие мышечного русла с его наибольшей разветвленностью в околокапиллярных отделах сохраняется в постнатальном онтогенезе, однако в месячном возрасте разветвленность в прекапиллярном отделе наиболее высокая. Другие возрастные изменения русел – удлинение стволов и симметризация разветвлений по сечению ветвей и углам их поворота – схожи в мозге и мышце.
4. Развитие гипертензии у крыс НИСАГ связано со структурными и функциональными изменениями русла, не однотипными в молодом и зрелом возрасте. На первом месяце их постнатальной жизни в мозге и, более выражено в мышце, наблюдается пониженная разветвленность артериального дерева и, как следствие этого, пониженная емкость русла, нарушенный продольный профиль гемодинамических показателей (напорного давления, линейной скорости кровотока и пристеночного напряжения сдвига) и увеличенное сосудистое сопротивление. Пониженная разветвленность русла может свидетельствовать о замедленном росте кровеносного дерева в этом возрасте.
5. К зрелому возрасту у крыс НИСАГ в исследованных органах появляется выраженная разветвленность прекапиллярных стволов, заметно превышающая таковую у зрелых крыс Вистар. Согласно модельным расчетам, эта разветвленность, если она не сопровождается образованием околокапиллярных сетей, должна нормализовать продольный профиль гемодинамических показателей и привести к снижению сосудистого сопротивления, особенно в мышце. Однако, если судить по величине артериального давления у крыс НИСАГ, этого не происходит.
6. У зрелых крыс НИСАГ повышена чувствительность артерий и сердца к стимулирующему действию катехоламинов, опосредованная a-адрено-рецепторами. Ее можно рассматривать как один из механизмов формирования системной гипертензии, обусловленной повышенной артериальной вазоконстрикцией и увеличенным сердечным выбросом.
Список публикаций по теме диссертации
1. , Носова кровеносного русла полушарий головного мозга у двухнедельных и взрослых крыс Вистар // II съезд физиологов Сибири и Дальнего Востока. Тез. докл. Часть 1. Новосибирск. – 1995. – С. 222.
2. Носова мозговых полушарий и двуглавой мышцы у крыс Вистар и НИСАГ // II съезд физиологов Сибири и Дальнего Востока. Тез. докл. Часть 2. – 1995. – С. 319.
3. , , Шошенко изменения кровеносного дерева и их гемодинамические последствия у крыс с гипертензией и сахарным диабетом // Микруциркуляция. Мат. междунар. конф. Ярославль. – 1997. – С. 56.
4. , , Носова структуры органного кровеносного дерева в онтогенезе и регионарные особенности кровотока // 3 съезд физиологов Сибири и Дальнего Востока. Тез. докл. Новосибирск. – 1997. – С. 268.
5. , , Якобсон сосудистой реактивности гипертензивных крыс линий НИСАГ // Бюлл. экспер. биол. и мед. 1998. – Т. 126. – №8. – С. 136-138.
6. , , Шошенко и гемодинамические особенности кровеносного русла мозга у гипертензивных крыс НИСАГ в постнатальном онтогенезе // Бюлл. СО РАМН. – 1998. – №4. – С. 35-42.
7. , , Носова кровеносного дерева и органоспецифичность кровоснабжения // Вестник РАМН. 1998. – №9. – С. 40-44.
8. , , Антипова сосудистых разветвлений и органная гемодинамика // XVII съезд физиологов России. Тез. докл. Ростов-на-Дону. – 1998. – С. 146.
9. , , Антипова кровеносной системы полушарий головного мозга у растущих крыс // Физиол. журн. им. . – 1998. –Т. 84. – №4. – С. 353-362.
10. , , Шошенко ангиоархитектоники головного мозга и скелетных мышц в постнатальном онтогенезе у крыс // Регуляция мозгового кровообращения в норме и патологии. Мат. симп. Ростов-на Дону. – 1999. – С. 32-34.
Таблица 1
Параметры гемодинамики в бассейне, образованном артерией и веной D 70 мкм,
у разновозрастных крыс Вистар (вверху) и НИСАГ (внизу)
(результат численного моделирования)
Параметры | 2-недельные | Месячные | Зрелые | |||
Мозг | мышца | мозг | мышца | мозг | мышца | |
Число генераций артериальных - венозных | 28 – 30 23 – 32 | 43 – 41 37 – 41 | 32 – 29 28 – 23 | 30 – 37 27 – 45 | 30 – 19 24 – 20 | 25 – 29 28 – 23 |
Число капиллярных концов артериальных - венозных | 333 – 162 | 2224 – 587 | 1308 – 167 | 2603 – 702 | 981 – 200 | 1357 – 594 |
356 – 204 | 988 – 550 | 660 – 254 | 1081 – 995 | 1434 – 229 | 3431 – 327 | |
Длина пути, мм (% пути по артериям, капиллярам и венам) | 5,8 (64+3+33) | 6,8 (61+3+36) | 5,8 (63+4+33) | 7,0 (52+3+45) | 5,7 (72+3+25) | 6,5 (43+6+51) |
5,4 (62+3+35) | 6,5 (66+3+31) | 6,2 (67+4+29) | 5,5 (55+4+42) | 5,0 (65+3+32) | 5,7 (60+7+33) | |
Соотношение вены / артерии одного диаметра | 2,1 | 4,5 | 7,9 | 3,7 | 4,9 | 2,3 |
1,7 | 2,1 | 2,6 | 1,2 | 6,9 | 10,4 | |
ΔP, μм рт. ст. (% ΔP β артериях, капиллярах и венах) | 9,3 (54+20+26) | 33,2 (76+7+17) | 22,8 (80+11+9) | 41,5 (74+6+20) | 21,7 (80+8+12) | 27,1 (63+16+21) |
12,2 (44+15+41) | 21,4 (58+10+32) | 11,9 (53+19+28) | 23,3 (44+9+47) | 24,7 (80+7+13) | 38,5 (77+12+11) | |
Сопротивление, 104 мм рт. ст. х с/мл | 73 91 | 39 56 | 50 46 | 41 57 | 57 45 | 52 29 |
