
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Определение эволюционного расстояния между нуклеотидными последовательностями
Истинные расстояния в моей модели: число точечных замен на 100 нуклеотидов | ||||||||||||
metj | a | uzel1 | uzel2 | uzel3 | uzel4 | b | c | d | e | f |
| |
metj | 0 | 90 | 50 | 55 | 60 | 65 | 90 | 90 | 90 | 90 | 90 |
|
a | 0 | 140 | 145 | 150 | 155 | 180 | 180 | 180 | 180 | 180 |
| |
uzel1 | 0 | 5 | 10 | 15 | 40 | 40 | 40 | 40 | 40 |
| ||
uzel2 | 0 | 15 | 10 | 35 | 35 | 35 | 45 | 45 |
| |||
uzel3 | 0 | 25 | 50 | 50 | 50 | 30 | 30 |
| ||||
uzel4 | 0 | 25 | 25 | 45 | 55 | 55 |
| |||||
b | 0 | 50 | 70 | 80 | 80 |
| ||||||
c | 0 | 70 | 80 | 80 |
| |||||||
d | 0 | 80 | 80 |
| ||||||||
e | 0 | 60 |
| |||||||||
f | 0 |
| ||||||||||
Матрица попарных расстояний, вычисленных по методу Джукса – Кантора | ||||||||||||
uzel1 | uzel2 | f | uzel4 | e | b | uzel3 | c | d | metj | a |
| |
uzel1 | 0 | 3,22 | 34,67 | 8,33 | 31,24 | 29,35 | 10,48 | 30,29 | 27,97 | 36,19 | 94,15 |
|
uzel2 | 0 | 39,85 | 11,94 | 34,67 | 24,83 | 6,94 | 25,71 | 25,27 | 41,48 | 101,1 |
| |
f | 0 | 23,52 | 45,42 | 65,14 | 43,71 | 64,39 | 62,19 | 69 | 134,2 |
| ||
uzel4 | 0 | 22,24 | 36,7 | 19,32 | 39,85 | 38,79 | 43,14 | 104,8 |
| |||
e | 0 | 58,66 | 42,03 | 59,36 | 59,36 | 68,21 | 125,3 |
| ||||
b | 0 | 17,31 | 37,22 | 50,19 | 61,47 | 138 |
| |||||
uzel3 | 0 | 19,32 | 31,72 | 48,37 | 110,1 |
| ||||||
c | 0 | 50,19 | 73,9 | 136,1 |
| |||||||
d | 0 | 62,19 | 118,8 |
| ||||||||
metj | 0 | 60,05 |
| |||||||||
a | 0 |
| ||||||||||
Матрица попарного сходства: среднее число совпадающих нуклеотидов на 100 позиций | ||||||||||||
uzel1 | uzel2 | f | uzel4 | e | b | uzel3 | c | d | metj | a |
| |
uzel1 | 0 | 3,15 | 27,76 | 7,89 | 25,55 | 24,29 | 9,78 | 24,92 | 23,34 | 28,71 | 53,63 |
|
uzel2 | 0 | 30,91 | 11,04 | 27,76 | 21,14 | 6,62 | 21,77 | 21,45 | 31,86 | 55,52 |
| |
f | 0 | 20,19 | 34,07 | 43,53 | 33,12 | 43,22 | 42,27 | 45,11 | 62,46 |
| ||
uzel4 | 0 | 19,24 | 29,02 | 17,03 | 30,91 | 30,28 | 32,81 | 56,47 |
| |||
e | 0 | 40,69 | 32,18 | 41,01 | 41,01 | 44,79 | 60,88 |
| ||||
b | 0 | 15,46 | 29,34 | 36,59 | 41,96 | 63,09 |
| |||||
uzel3 | 0 | 17,03 | 25,87 | 35,65 | 57,73 |
| ||||||
c | 0 | 36,59 | 47 | 62,78 |
| |||||||
d | 0 | 42,27 | 59,62 |
| ||||||||
metj | 0 | 41,32 |
| |||||||||
a | 0 |
| ||||||||||
Сравнение расстояний, полученных на основе частоты замен и метода Джукса - Кантора с истинным
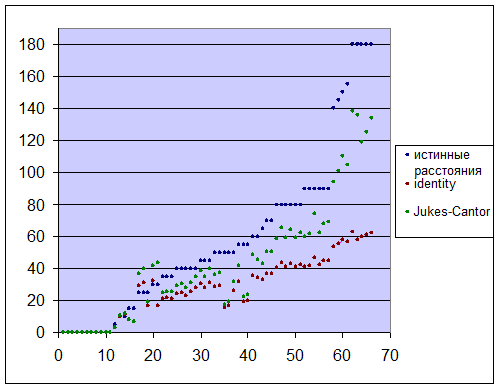
На графике видно, что расстояния, полученные по частоте замен и по методу Джукса-Кантора меньше истинных, что особенно хорошо проявляется для далеких последовательностей. Однако метод Джукса-Кантора лучше отражает реальную картину, чем метод по частоте замен. Это может объясняться следующим:
Истинный ход эволюции отличается от того, что мы можем реконструировать отличается возможностями обратных и повторных (замен на различные нуклеотиды больше 1 раза в одной позиции) замен. На основе частоты замен невозможно учесть оба этих события. Предположим, у нас есть две последовательности, между которыми произошло 20 замен. Значение идентичности не отражает того, что 1) могло произойти сперва 30 замен, а потом 10 обратных или 2) произошло 15 замен, потом еще 15 - но 5 для одних и тех же позиций
Метод Джукса-Кантора учитывает возможность обратных замен, но не отражает возможность замен на одних и тех же позициях.
Поэтому данный метод лучше реконструирует картину эволюции. Но для далеких последовательностей и данный метод не является оптимальным, т. к. (см. график выше) резко возрастает разница между расстояниями, рассчитанными по данному методу и истинными.
График зависимости 2-х оценок расстояния от величины «истинного» расстояния
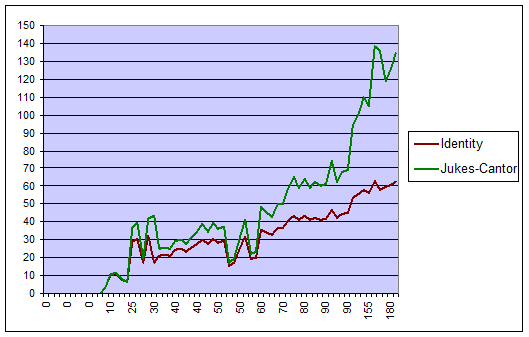
На данном графике четко видно, что метод Джукса-Кантора более точно определяет истинные расстояния, чем метод, основанный на частоте замен.
