

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
К истории развития флоры и растительности Амурской области // Ученые записки Забайкальского государственного гуманитарно-педагогического университета им. Чернышевского. Серия «Естественные науки» .– 2009. – № 1. – С. 100-104.
К ИСТОРИИ РАЗВИТИЯ ФЛОРЫ И РАСТИТЕЛЬНОСТИ АМУРСКОЙ ОБЛАСТИ.
Благовещенск. Амурский филиал Ботанического сада-института ДВО РАН
Starchenkoamur@mail.ru
Сведения по истории развития флоры и растительности отражены в работах ботаников, палеоботаников, палинологов и других специалистов (Аблаев, 1978; Ахметьев, 1973; Вахрамеев, 1988; Вербицкая, 1962; Камаева, 1990; Кезина, 2005, 2007; Красилов, 1972а, 1972б, 1976, 1989; Криштофович, ; Мамонтова, 1975, 1977а, 1977б, 1978; Махова, 1971; Финько, Заклинская., 1958; Хлонова, 1961, 1969, 1974). Несмотря на внимание, уделяемое вопросам происхождения флоры и растительности, по-прежнему очень далеко до создания общей картины. Крайне мало имеется фактических данных по возникновению и развитию растительности в раннепалеозойские эпохи. По сохранившимся находкам растительных остатков более изученными оказались каменноугольный, пермский и последующие периоды.
Общепринятая точка зрения предполагает, что первые растения, которые можно считать высшими, возникли в палеозое от гипотетических предковых форм водорослей. Выход растений на сушу начался в девоне, а относительно богатая флора высших споровых сформировалась в каменноугольный период (Криштофович, 1955, 1957, 1958; Тахтаджян, 1970, 1980; Недолужко, 1997). Последующие периоды, учитывая сложные геологические и климатические процессы, привели к появлению более высокоразвитых групп высших растений, в том числе первых покрытосеменных на границе юрского и мелового периодов (Криштофович, 1957, 1958; Sun Ge, Dilcher, 2002).
Находки растительных остатков на территории Амурской области известны благодаря отработке и изучению буроугольных залежей (Криштофович, 1958; Шульман, 1971; Ахметьев, 1973, 1990; Красилов, 1976, 1985; Камаева, 1990; Кезина, 2000, 2005 и др.). Полученные палеоботанические и палинологические данные очень важны, т. к. меловой комплекс является тем основанием, на котором развивалась все более поздние и современные флоры обоих полушарий (Криштофович, 1958; Ахметьев, 1973).
В палеоцене из основной меловой флоры начинают дифференцироваться различные флоры в Европе, Америке и Азии. Новые флоры продолжали далее развиваться частью автохтонно, частью принимая в себя новые виды с севера, благодаря чему постоянно образовывались смешанные флоры, в которых виды, господствующие прежде, постепенно либо сменялись более молодыми, либо становились реликтами. В эоцене в области Арктики начинает выделяться многочисленная группа сережкоцветных, которая мигрирует на юг, смешивается с автохтонными видами и создает устойчивый комплекс голарктической третичной тургайской флоры. Эта флора охватила в олигоцен всю Азию от впадины Тургайского пролива до Японии и Камчатки, далее через Берингию - Аляску и всю западную Америку до естественных границ умеренной флоры. Позже эта флора с изменениями проникла и распространилась по Европе. В этой флоре смешались поздние виды сережкоцветных (Platanus, Zelcovia, Castanea, Juglans, Fagus, Ulmus, Acer и др.), более ранние виды покрытосеменных и голосеменных, в первую очередь, Ginkgo. С середины олигоцена в результате изменений климата началось постепенное выпадение субтропических флор в Европе, Азии и Северной Америке при частичном сохранении отдельных видов как третичных реликтов. На территории Амурской области следы субтропической флоры найдены в растительных остатках позднего миоцена (бузулинская свита).
Позднее начинается дифференциация областей голарктической флоры и постепенный распад на палеоарктическую и неоарктическую флоры. Локальное развитие травяных равнин (степей) в Азии началось, вероятно, в эоцене и наиболее активно продолжилось в позднем миоцене. Характер остатков споропыльцевых комплексов плиоценовой флоры указывает на такие же процессы в районах внутренней Азии (Ахметьев, Шевырева, 1989; Вахрамеев, 1988). Обширность территории, активные климатические процессы создали в северном полушарии гораздо более сложные взаимоотношения флор, чем в южном полушарии.
В результате совместных работ палеоботаников ГИН РАН и палинологов АмурКНИИ в гг. проведен детальный анализ флоры и растительности позднего мела и кайнозоя на территории Амурской области (Верхнее Приамурье). По разрезам буроугольных месторождений выявлены основные семейства и роды, дана примерная оценка разнообразия жизненных форм третичной флоры, установлено заметное снижение количества древесных форм на рубеже мела и палеогена и увеличение числа покрытосеменных, что указывает на значительные климатические изменения (Красилов, 1976, 1985; Ахметьев и др., 2002; Кезина, 2005). Это доказывает, что рассматриваемая флора имела широкий климато-экологический спектр, что положительно отразилось на ее дальнейших эволюционных изменениях (Кезина, 2005).
Эоценовая флора являлась наиболее теплолюбивой из кайнозойских флор и полидоминантной по составу, включая виды, различные по происхождению (Кезина, 2005). В олигоцене, в основном, завершилось становление флоры тургайского экотипа. Флора олигоцена утрачивает многие термофильные и экзотические (субтропические) элементы, но увеличивается доля Ореховых, Буковых и Березовых. Для флоры отмечено доминирование родов и видов, сближаемых с современными таксонами (рис.).
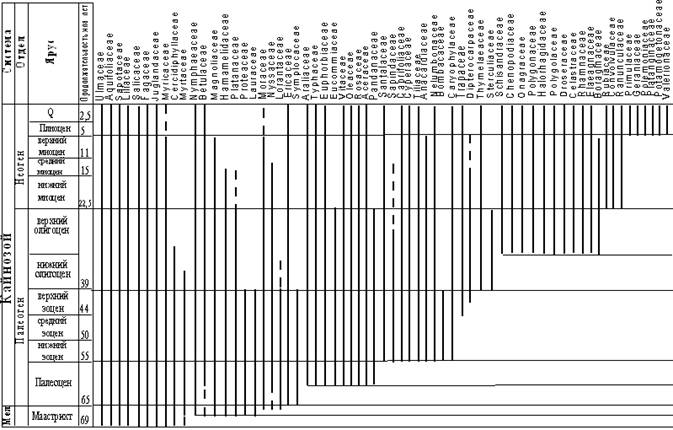
Рис. Распределение семейств покрытосеменных в позднеммаастриских и кайнозойских флорах Верхнего Приамурья (Кезина, 2007).
Климат миоцена характеризовался повышением температур, приведшим к наступлению климатического оптимума на рубеже раннего и среднего миоцена, и последующим ухудшением климатических показателей (Кезина, 2005). Флора миоцена является по составу гетерогенной, мезофильной и характеризуется дальнейшим уменьшением термофильных элементов. Субтропические американо-восточноазиатские роды (Sequoia, Taxodium, Castanea, Magnolia и др.) сменяются современными хвойными и разнообразными Betulaceae, Salicaceae, Ulmaceae, Aceraceae. Существенную роль играют кустарничковые и травянистые покрытосеменные. К концу миоцена происходит замещение многих оставшихся термофильных видов широко распространенными голарктическими (рис.). В этот период флоры юга Дальнего Востока наиболее близки между собой по видовому составу. Постепенное исчезновение термофильных, в основном, вечнозеленых растений, перераспределение доминантов, субдоминантов и сопутствующих видов стали основными факторами, формирующими полидоминантную многоярусную хвойно-широколиственную растительность (Кезина, 2005).
В плиоцене на территории Верхнего Приамурья отмечается усиление тектонической активности. Процессы воздымания в отдельных районах соседствовали с процессами погружения, что способствовало образованию мелководных, но обширных по площади пресноводных водоемов. Растительность к концу плиоцена уже полностью соответствует современной лесной растительности Маньчжурии и юга РДВ, представляя собой широколиственные и хвойно-широколиственные леса с заметным участием третичных реликтов (Кезина, 2005).
К концу плиоцена только крайний юго-восток Сибири еще сохранял обедненную тургайскую флору, дериватом которой являлась амуро-уссурийская флора. Остальная Сибирь была покрыта хвойной тайгой. Климатические условия во время голоцена на территории северного полушария не были постоянными. Отмечены периоды относительного потепления и резкие похолодания вплоть до локальных оледенений. На территории Дальнего Востока климат позднего голоцена был переходным от сурового климата конца плейстоцена к более благоприятному климату раннего и, особенно, среднего голоцена. В позднем голоцене произошло некоторое общее изменение климатических условий в сторону похолодания (Нейштадт, 1957). Именно изменение климатических условий вызывало изменение границ существовавших в то время зон растительности и ареалов многих видов.
В четвертичную эпоху (четвертичная флора) происходит географическое перераспределение отдельных флор, а появление новых форм растительности ограничено возникновением новых рас и разновидностей – редко – новых видов и вымиранием отдельных форм повсеместно или на отдельных территориях.
На территории Амурской области сплошное оледенение отсутствовало, в ледниковые эпохи увеличивали свои площади только горные ледники. На равнинах ледниковые условия не сложились из-за небольшого количества выпадающего зимой снега, но смена растительности происходила в сторону увеличения площади таёжных лесов и марей. Теплолюбивые растения отступали на юг, на их место приходили холодостойкие. Районы, не подвергавшиеся сплошному оледенению, в четвертичную эпоху сохранили связи с южными флорами и, несмотря на суровость климата, разнообразный растительный покров с большим числом реликтов (виды Trapa, Brasenia, Aldrovanda, Phellodendron amurense, Juglans mandshurica и др.).
Современные флора и растительность Амурской области неоднородны. Значительную часть территории занимает светлохвойная тайга с участием лиственницы, сосны и других голосеменных. В случае различных крупных изменениях окружающей среды, в первую очередь, антропогенных, зональные таежные леса замещаются производными березовыми лесами, в которых процент хвойных деревьев зависит от возраста производных лесов и нарушенности территории. Темнохвойные леса с участием ели и пихты занимают незначительный процент территории Амурской области преимущественно на востоке и северо-востоке. (Карта…, 1969). На юге области сохранились различной степени нарушенности различные неморальные леса с участием дуба монгольского, широколиственных пород и отдельных реликтов, являющиеся крайне обедненным дериватом тургайской флоры. На крайнем юго-востоке Амо неморальная растительность представлена кедрово-широколиственной тайги с участием кедра сибирского и значительным участием реликтовых третичных видов.
Специфика флоры и растительности области связана преимущественно с двумя факторами: принадлежностью основной части территории области к бассейну Амура и расположением области на границе Восточноазиатской и Бореальной флористических областей. На южные, центральные и восточные районы области прослеживается влияние Тихого океана.
Библиографический список
1. Аблаев и история флор побережий Японского моря. – М.: Наука, 1978. – 172 с.
2. Ахметьев и эоценовые флоры юга Дальнего Востока, их стратиграфическое значение и климатическая характеристика // Сов. геология– № 7. – С. 14-29.
3. Ахметьев и миоценовые флоры юга Дальнего Востока СССР как показатели климатической обстановки // Изв. АН СССР. Сер. геол– № 4. – С. 134-144.
4. Ахметьев дифференциация позднемеловых и кайнозойских флор Земного шара на фоне геологических событий. - М.: ВИНИТИ, 1990. – 100 с. – (Итоги науки и техники. Сер. Стратиграфия. Палеотология).
5. , , Шелудченко похолодание на востоке СССР по палеоботаническим данным // Бюл. МОИП. Отд. геол– Т. 61, вып. 4. – С. 87-100.
6. , Шевырева цагаянского типа в Призайсанье (Восточный Казахстан) // Изв. АН СССР. Сер. геол– № 6. – С. 80–89.
7. , , КодАД-РУл Т. М., Манчестер и флора пограничных слоев мела и палеогена юго-восточной части Зейско-Буреинского осадочного бассейна: сб. памяти В. А. Вахрамеева. – М.: ГЕОС, 2002. – С. 275-315.
8. , , Манчестер мел-палеогеновых континентальных отложений Зейско-Буреинского бассейна (Приамурья) на основе комплексов растительных микро - и макрофоссилий: Методические аспекты палинологии. – М.: ИГИРГИ. 2002.– С. 10–12.
9. Вахрамеев и меловые флоры и климаты земли. – М.: Наука, 1988. – 214 с.
10. Вербицкая обоснование стратиграфического расчленения меловых отложений Сучанского каменноугольного бассейна – М.; Л.: Изд-во АН СССР, 1962. – 165 с. – (Тр. лаб. геологии угля АН СССР; Вып. 15).
11. Камаева и флора пограничных отложений мела и палеогена Зейско-Буреинской впадины. – Хабаровск: ДВНЦ АН СССР, 1990. – 67 с.
12. Карта растительности бассейна Амура. / , , и др.; Под. ред. . - М 1:2500000 – М.: ГУГК, 1969.
13. Кезина Верхнего Приамурья в позднем мезозое // Тихоокеан. геология№ 6 - С. 24-34.
14. Кезина кайнозоя Верхнего Приамурья. – Владивосток: Дальнаука, 2000. – 51 с. , 13 л. табл.
15. Кезина угленосных отложений позднего мела и кайнозоя Верхнего Приамурья. - Владивосток: Дальнаука, 2005. – 206 с.
16. Кезина угленосных отложений позднего маастрихта и кайнозоя Верхнего Приамурья: Автореф. дис. … докт. геол.-минерал. Наук. – Сыктывкар, 2007. – 32 с.
17. Кезина, 2000,2005, 2007;
18. Красилов флора реки Буреи. – М.: Наука, 1972а. – 151 с.
19. Красилов наземных растений. – Владивосток: ДВНЦ АН СССР, 1972б. – 207 с.
20. Красилов берингийских связей в формировании кайнозойской флоры Восточной Азии и Северной Америки // Берингия в кайнозое. – Владивосток: ДВНЦ АН СССР, 1976а. – С. 129-344.
21. Красилов флора Амурской области. – М.: Наука, 1976. – 92 с.
22. Красилов период эволюции земной коры и биосферы. – М.: Наука, 1985. – 240 с.
23. Красилов Дальнего Востока: геодинамическая схема, опорные разрезы, флорогенез // Кайнозой Дальнего Востока. – Владивосток: ДВО АН СССР, 1989. – С. 23-34.
24. Красилов, 1972а, 1972б, 1976, 1985, 1989;
25. Криштофович ботанико-географических областей Северного полушария с начала третичного периода//Вопросы геологии Азии. – М.; Л.: Изд-во АН СССР, 1955. – Т. 2. – С. 824-844.
26. Криштофович . - 4-е изд. - Л.: Изд-во АН СССР, 19с.
27. Криштофович Ангарской суши//Мат-лы по истории флоры и растительности СССР. - М.; Л.: Изд-во АН СССР, 1958. - Вып. 3. - С. 7-41.
28. Криштофович и развитие мезозойской флоры // Избр. тАД-РУды. - М.; Л.: Изд-во АН СССР, 1959. –С. 179-199.
29. Мамонтова -пыльцевая характеристика и корреляция континентальных отложений Ушумунской впадины // Тр. СахКНИИ ДВНЦ АН СССР. 1975. – Вып. 36. – С. 32-40.
30. Мамонтова переходных слоев верхнего мела и палеогена Амуро-Зейской депрессии // Палеоботаника на Дальнем Востоке. – Владивосток: ДВНЦ АН СССР, 1977 а. – С. 32-37.
31. Мамонтова и корреляция кайнозойских отложений Амуро-Зейской депрессии по палинологическим критериям // Геология Верхнего Приамурья. – Владивосток: Б. и., 1977 б. С. 19-22.
32. Мамонтова флора Амуро-Зейской депрессии // Стратиграфия кайнозойских отложений Дальнего Востока. – Владивосток, 1978. – С. 35-41.
33. Махова черты растительности Амуро-Зейской равнины в неогене и плестоцене по палинологическим данным: Автореф. дис. … канд. геол.-минерал. наук. – М. 1971. – 35 с.
34. Недолужко растения: проблема эволюции жизненных форм. – Владивосток: Дальнаука, 1997. – 120 с.
35. Недолужко арборифлоры российского Дальнего Востока // Комаров. чтения. - Владивосток, 1997. - Вып. 43. - С. 83-127.
36. Нейштадт лесов и палеогеография СССР в голоцене/Отв. ред. . – М.: Изд-во АН СССР, 1957 – 404 с.
37. , К вопросу о стратиграфии рыхлых отложений Зейско-Бурейской депресии // Изв. АН СССР. Сер. геол., 1958. – № 2. – С. 25-43.
38. Хлонова и пыльца верхней половины верхнего мела восточной части Западно-Сибирской низменности. - Новосибирск: СО АН СССР, 1961. – 138 с. – (Тр. Ин-та геологии и геофизики СО АН СССР, вып. 7).
39. Хлонова -пыльцевая характеристика меловых отложений Зея-Буреинской впадины // Спорово-пыльцевые комплексы мезозоя Сибири и Дальнего Востока. – М.: Наука, 1969. – С. 5-66.
40. Хлонова меловых отложений Сибири и Дальнего Востока. – Новосибирск: Наука, 1974. – 166 с.
41. Хлонова характеристика меловых отложений на р. Кие (Западная Сибирь). – М.: Наука, 1976. – 103 с.
42. Шульман изучения природы Амурской области //Вопросы географии Верхнего Приамурья. – Благовещенск: Б. и., 1971. – С. 58-98.
43. Тахтаджян и расселение цветковых растений. – Л.: Наука, 1970. – 145 с.
44. Тахтаджян цветковых растений// Жизнь растений. - М.: Просвещение, 1980. - Т. 5: Цветковые растения, ч. 1. - С. 103-112.
45. Sun Ge, Dilcher D. L. Early Angiosperms from the Lower Cretaceous of Jixi, eastern Heilongjiang, China // Rev. Palaeobot. Palynol– Vol. 121. – P. 91-112.
