
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
МИНИСТЕРСТВО ВНУТРЕННИХ ДЕЛ РОССИЙСКОЙ ФЕДЕРАЦИИ КРАСНОДАРСКИЙ УНИВЕРСИТЕТ МВД РОССИИ
КАФЕДРА ПСИХОЛОГИИ И ПЕДАГОГИКИ
«У Т В Е Р Ж Д А Ю»
Начальник кафедры
подполковник полиции
______________
«___» ___________ 2012 г.
Дисциплина: «Анатомия и физиология центральной нервной системы ».
По направлению подготовки (специальности) 050407.65 Педагогика и психология девиантного поведения
специализация - Социальная педагогика
ЛЕКЦИЯ
ТЕМА 16: «Физиология коры головного мозга»
Обсуждена и одобрена на заседании кафедры Протокол № от «___»_________20 г. | Подготовил: начальник кафедры психологии и педагогики |
Краснодар 2012
Объем времени, отводимый на лекцию: 2 часа.
Место проведения: учебная аудитория
Методика проведения: классическая лекция.
Основное содержание темы:
Кора головного мозга относится к самому молодому отделу головного мозга. Наряду с этим она осуществляет высшую регуляторную функцию в организме и формирует психофизиологическую деятельность, определяющую поведение человека. Кора больших полушарий представляет собой слой серого вещества общей площадью 1см2, покрывающий большие полушария. В состав серого вещества входит 14 миллиардов нейронов, а количество глиальных клеток почти в 10 раз больше. Кора больших полушарий составляет около 40% всей массы головного мозга. Структурной единицей коры полушарий, как и всей нервной системы, является нейрон.
Основные термины и понятия:
Нейрон, нервная ткань, миелинезированные и немиелинизированные тела нейронов, униполярные, биполярные, псевдоуниполярные, полиполярные нейроны.
Цели занятия:
1. Формирование у курсантов, слушателей, прочных знаний о структурно-функциональных характеристиках коры головного мозга.
2. Закрепить знания в областях коры больших полушарий.
3. Содействовать осознанию курсантами, студентами и слушателями важности изучаемого предмета.
План с занятия:
Введение.
1. Структурно-функциональная характеристика коры головного мозга.
2. Чувствительные, двигательные и ассоциативные области коры больших полушарий.
Заключение (выводы).
Литература:
1. , , Фельдман центральной нервной системы. – Ростов-на-Дону: Феникс, 2006.
2. , Цехмистренко нервной системы. – М.: Мир, 2006.
3. Физиология человека: учебник для мед. ин-тов / под ред. . М.: 2009.
4. , Туровский центральной нервной системы для психологов. – СПб.: Питер, 2009.
5. , Кульба центральной нервной системы. Р-н-Д 2008 г.
Дополнительная литература
1. Антонова и физиология центральной нервной системы. – М.: Высшее образование, 2006.
2. , Туровский центральной нервной системы для психологов - СПб.: Питер, 2007
3. Дерягина физиологии центральной нервной системы. – М.: Изд-во МосУ МВД России, 2008
4. , Туровский центральной нервной системы для психологов - СПб.: Питер, 2007.
5. , Яковлев центральной нервной системы. – М.: Academia, 2004.
6. , , Фельдман центральной нервной системы. – Ростов-на-Дону: Феникс, 2006.
Введение
Исключительная роль новой коры головного мозга в организации сложных форм высшей нервной деятельности, ее более позднее эволюционное возникновение и наиболее бурное развитие, высший иерархический уровень вертикально согласованной деятельности центральной нервной системы составляют наиболее существенные особенности этого отдела мозга. Они привлекли и продолжают удерживать внимание многих исследователей к изучению физиологии коры больших полушарий головного мозга.
Новая кора (неокортекс) представляет собой слой серого вещества общей площадью 1500— 2200 см2, покрывающий большие полушария конечного мозга. Она составляет около 72 % всей площади коры и около 40 % массы головного мозга. В коре имеется около 14 млрд. нейронов, количество глиальных клеток примерно в 10 раз больше. Кора головного мозга является в филогенетическом плане наиболее молодой нервной структурой, у человека она осуществляет высшую регуляцию функций организма и психофизиологические процессы, обеспечивающие различные формы поведения.
Вопрос 1.
Структурно-функциональная характеристика коры
Слои коры большого мозга. В направлении с поверхности в глубь коры различают шесть горизонтальных слоев.
Молекулярный слой имеет очень мало клеток, но большое количество ветвящихся дендритов пирамидных клеток, формирующих сплетение, расположенное параллельно поверхности. На этих дендритах образуют синапсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер таламуса.
Наружный зернистый слой составлен в основном звездчатыми клетками и частично малыми пирамидными клетками. Волокна клеток этого слоя расположены преимущественно вдоль поверхности коры, образуя кортикокортикальные связи.
Наружный пирамидный слой состоит преимущественно из пирамидных клеток средней величины. Аксоны этих клеток, как и зернистые клетки II слоя, образуют кортикокортикальные ассоциативные связи.
Внутренний зернистый слой по характеру клеток (звездчатые клетки) и расположению их волокон аналогичен наружному зернистому слою. На нейронах этого слоя образуют синаптиче-ские окончания афферентные волокна, идущие от нейронов специфических ядер таламуса и, следовательно, от рецепторов сенсорных систем.
Внутренний пирамидный слой образован средними и крупными пирамидными клетками, причем гигантские пирамидные клет-
ки Беца расположены в двигательной коре. Аксоны этих клеток образуют эфферентные кортикоспинальные и кортикобульбарный двигательные пути.
Слой полиморфных клеток образован преимущественно веретенообразными клетками, аксоны которых образуют кортико-таламические пути.
Оценивая в целом афферентные и эфферентные связи коры, можно отметить, что в слоях I и IV происходят восприятие и обработка поступающих в кору сигналов. Нейроны II и III слоев осуществляют кортикокортикальные ассоциативные связи. Покидающие кору эфферентные пути формируются преимущественно в V—VI слоях. Более детально деление коры на различные поля проведено на основе цитоархитектонических признаков (формы и расположения нейронов) К. Бродманом (1909), который выделил 52 поля, многие из которых характеризуются функциональными и нейрохимическими особенностями.
Гистологические данные показывают, что элементарные нейронные цепи, участвующие в обработке информации, расположены перпендикулярно поверхности коры. Электрофизиологические исследования (В. Маунткасл, 1957) с погружением микроэлектродов перпендикулярно поверхности соматосенсорной коры показали, что все встречаемые на пути нейроны отвечают на раздражитель только одного качества, например тактильный. Напротив, при погружении электрода под углом на его пути попадались нейроны разной сенсорности. В результате был сделан вывод о наличии в коре мозга функциональных объединений нейронов, расположенных в цилиндрике диаметром 0,5— 1,0 мм, включающем все слои коры и содержащем несколько сотен нейронов. Такие структуры были названы нейронными колонками, обнаруженными в моторной коре и различных зонах сенсорной коры. Соседние нейронные колонки могут частично перекрываться, а также взаимодействовать друг с другом по механизму латерального торможения и осуществлять саморегуляцию по типу возвратного торможения.
Кортикализация функций — возрастание в филогенезе роли коры большого мозга в анализе и регуляции функций организма и подчинение себе нижележащих отделов ЦНС. Например, регуляция таких двигательных функций, как локомоция (прыжки, ходьба, бег) и выпрямительные рефлексы, у низших позвоночных (амфибий) полностью обеспечивается стволом мозга, и удаление больших полушарий практически их не изменяет. У кошек перерезка ствола между средним и промежуточным мозгом существенно не влияет на выпрямительные рефлексы, но нарушает в остром периоде локомоцию, которая в дальнейшем частично восстанавливается. Выключение коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и выпрямительных рефлексов.
Результаты изучения локализации функций позволили ученым в середине XX в. разделить кору головного мозга на различные функциональные зоны — картирование коры. Детальные функциональные карты (К. Клейст, 1959) используются в неврологической практике, однако надо иметь в виду упрощенный характер этих схем. Одним из наиболее крупных вариантов функционального разделения новой коры головного мозга является выделение в ней сенсорной, ассоциативной и двигательной областей.
Вопрос 2.
Сенсорные области коры
Сенсорные области коры — зоны, в которые проецируются сенсорные раздражители (синонимы: проекционная кора, корковые отделы анализаторов). Они расположены преимущественно в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от специфических сенсорных ядер таламуса (вентральных, задних латерального и медиального). Сенсорная кора имеет хорошо выраженные II и IV слои и называется гранулярной.
Зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма, называются первичными сенсорными областями (ядерными частями анализаторов, как считал ). Они состоят преимущественно из мономодальных нейронов и формируют ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное (топографическое) представительство частей тела, их рецепторных полей. Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, полимодальные нейроны которых отвечают на действие нескольких раздражителей.
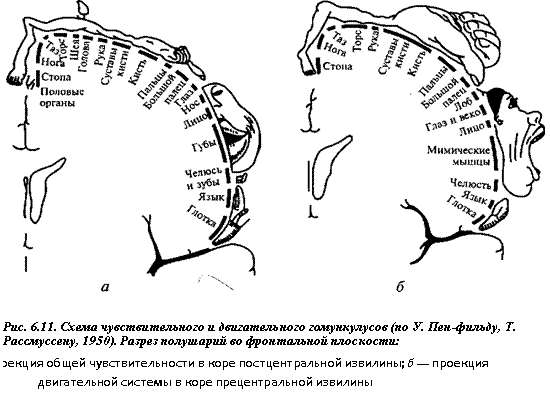
Важнейшей сенсорной областью является теменная кора постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий (поля 1 — 3), которую обозначают как первичная соматосенсорная область (S I). Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорно-двигательного аппарата от мышечных, суставных, сухожильных рецепторов. Проекция участков тела в этой области характеризуется тем, что проекция головы и верхних отделов туловища расположена в нижнелатеральных участках постцентральной извилины, проекция нижней половины туловища и ног — в верхнемедиальных зонах извилины, проекция нижней части голени и стоп — в коре парацентральной дольки на медиальной поверхности полушарий (см. рис. 6.11). При этом проекция наиболее чувствительных участков (язык, губы, гортань, пальцы рук) имеет относительно большие зоны по сравнению с другими частями тела. Предполагается, что в зоне тактильной чувствительности языка расположена проекция и вкусовой чувствительности (первичная сенсорная зона S I).
Кроме S I выделяют меньших размеров вторичную соматосен-сорную область S II, расположенную на верхней стенке боковой борозды, на границе ее пересечения с центральной бороздой. Функции филогенетически более древней S II изучены плохо. Известно, что локализация поверхности тела в ней менее четкая, импульсация сюда поступает как от противоположной, так и от «своей» стороны, предполагают ее участие в сенсорной и моторной координации двух сторон тела.
Другой первичной сенсорной зоной является слуховая кора (поля 41, 42), которая расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). В этой зоне в ответ на раздражение слуховых рецепторов кортиева органа формируются звуковые ощущения, изменяющиеся по громкости, тону и другим качествам. Здесь имеется четкая топическая проекция: в разных участках коры представлены различные участки кортиева органа. К проекционной коре височной доли относится предположительно также центр вестибулярного анализатора в верхней и средней височных извилинах (поля 20 и 21). Обработанная сенсорная информация используется для формирования «схемы тела» и регуляции функций мозжечка (височно-мостомозжечковый путь).
Еще одна первичная проекционная область новой коры расположена в затылочной коре — первичная зрительная область (кора части клиновидной извилины и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки, и каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения. Раздражение коры 17-го поля приводит к возникновению световых ощущений. Около поля 17 расположена кора вторичной зрительной области (поля 18 и 19). Нейроны этих зон полимодальны и отвечают не только на световые, но и на тактильные, слуховые раздражители. В данной зрительной области происходит синтез различных видов чувствительности и возникают более сложные зрительные образы и их опознание. Раздражение этих полей вызывает зрительные галлюцинации, навязчивые ощущения, движение глаз.
 |
Основная часть информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору.
Ассоциативные области коры
Ассоциативные области коры (синонимы: межсенсорная, межанализаторная кора) включают участки новой коры, расположены рядом с сенсорными и двигательными зонами, но не выполняющие непосредственно чувствительных или двигательных функций. Границы этих областей обозначены недостаточно четко, неопределенность преимущественно связана со вторичными проекционными зонами, функциональные свойства которых являются переходными между свойствами первичных проекционных и ассоциативных зон. Ассоциативная кора является филогенетически наиболее молодой частью новой коры, получившей наибольшее развитие у приматов и человека. У последнего она составляет около 50 % всей коры или 70 % неокортекса.
Основной физиологической особенностью нейронов ассоциативной коры, отличающей их от нейронов первичных зон, является полисенсорность (полимодальность): они отвечают с почти одинаковым порогом не на один, а на несколько раздражителей — зрительные, слуховые, кожные и др. Полисенсорность нейронов ассоциативной коры создается как ее кортикокортикальными связями с разными проекционными зонами, так и главным ее афферентным входом от ассоциативных ядер таламуса, в которых уже произошла сложная обработка информации от различных чувствительных путей. В результате этого ассоциативная кора представляет собой мощный аппарат конвергенции различных сенсорных возбуждений, позволяющий произвести сложную обработку информации о внешней и внутренней среде организма и использовать ее для осуществления высших психофизиологических функций.
По таламокортикальным проекциям выделяют три ассоциативные системы мозга: таламотеменную, таламолобную и таламовисочную систему.
Таламотеменная система представлена ассоциативными зонами теменной коры (поля 5, 7, 40), получающими основные афферентные входы от задней группы ассоциативных ядер таламуса (латеральное заднее ядро и подушка). Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, моторную кору и ядра экстрапирамидной системы. Основными функциями таламотеменной системы являются гнозис, формирование «схемы тела» и праксис. Под гнозисом понимают функцию различных видов узнавания: формы, величины, значения предметов, понимание речи, познание процессов, закономерностей и др. К гностическим функциям относится оценка пространственных отношений, например взаимного расположения предметов. В теменной коре выделяют центр стереогнозиса, расположенный сзади от средних отделов постцентральной извилины (поля 7, 40, частично 39) и обеспечивающий способность узнавания предметов на ощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела («схемы тела»), центр которой расположен в поле 7 теменной коры. Под праксисом понимают целенаправленное действие, центр его находится в надкраевой извилине (поля 39 и 40 доминантного полушария) и обеспечивает хранение и реализацию программы двигательных автоматизированных актов (например, рукопожатие, причесывание, зажигание спички и др.).
Таламолобная система представлена ассоциативными зонами лобной коры (поля 9 — 14), имеющими основной афферентный вход от ассоциативного медиодорсального ядра таламуса. Главной функцией лобной ассоциативной коры является формирование программ целенаправленного поведения, особенно в новой для человека обстановке. Реализация этой общей функции основывается на других функциях таламолобной системы: 1) формирование доминирующей мотивации, обеспечивающей направление поведения человека. Эта функция основана на тесных двусторонних связях лобной коры с лимбической системой и ролью последней в регуляции высших эмоций человека, связанных с его социальной деятельностью и творчеством; 2) обеспечение вероятностного прогнозирования, что выражается изменением поведения в ответ на изменения обстановки окружающей среды и доминирующей мотивации; 3) самоконтроль действий путем постоянного сравнения результата действия с исходными намерениями, что связано с созданием аппарата предвидения (акцептор результата действия в функциональной системе согласно теории ).
В результате проведения по медицинским показаниям префронтальной лоботомии, при которой пересекаются связи между лобной долей и таламусом, наблюдается развитие «эмоциональной тупости», отсутствие мотивации, твердых намерений и планов, основанных на прогнозировании. Такие люди становятся грубыми, нетактичными, ненадежными, у них появляется тенденция к повторению каких-либо двигательных актов, хотя обстановка уже изменилась и надо выполнять другие действия.
Таламовисочная система не получила еще достаточной научной проработки. Говоря о роли височной коры, надо отметить, что некоторые ассоциативные центры, например стереогнозиса и праксиса, включают в себя и участки височной коры (поле 39). В височной коре расположен слуховой центр речи Вернике, находящийся в задних отделах верхней височной извилины (поля 22, 37 и 42 левого доминантного полушария). Этот центр обеспечивает речевой гнозис — распознание и хранение устной речи, как собственной, так и чужой. В средней части верхней височной извилины (поле 22) находится центр распознания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей (поле 39) находится центр чтения письменной речи, обеспечивающий распознание и хранение образов письменной речи.
Психофизиологические функции, осуществляемые ассоциативной корой, инициируют поведение организма, обязательным компонентом которого являются произвольные целенаправленные движения, осуществляемые при обязательном участии двигательной коры.
Двигательные области коры
Понятие о двигательной коре больших полушарий начало формироваться с 80-х годов XIX в., когда было показано, что электрическое раздражение у животных некоторых корковых зон вызывает движение конечностей противоположной стороны (Г. Фрич, Е. Гитциг, 1870). В двигательной коре выделяют первичную и вторичную моторные области.
В первичной моторной коре (прецентральная извилина, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела (см. рис. 6.11). При этом проекции мышц нижних конечностей и туловища расположены в верхних участках прецентральной извилины и занимают сравнительно небольшую площадь, а проекции мышц верхних конечностей, лица и языка расположены в нижних участках извилины и занимают большую площадь («двигательный человечек» Пенфильда). Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры. Двигательные реакции на раздражение первичной моторной коры осуществляются с минимальным порогом (высокая возбудимость), они представлены элементарными сокращениями мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральное). При поражении этой корковой зоны утрачивается способность к тонким координированным движениям рук, особенно пальцев.
Вторичная двигательная кора (поле 6) расположена на латеральной поверхности полушарий, впереди прецентральной извилины (премоторная кора). Она осуществляет высшие двигательные функции, связанные с планированием и координацией произвольных движений. Кора поля 6 получает основную часть эфферентной импульсации базальных ядер и мозжечка и участвует в перекодировании информации о программе сложных движений. Раздражение коры поля 6 вызывает более сложные координированные движения поля, например поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения мышц-сгибателей или мышц-разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле 44), обеспечивающие речевой праксис, а также, музыкальный моторный центр (поле 45), определяющий тональность речи, способность петь.
В моторной коре лучше, чем в других зонах коры, выражен слой, содержащий гигантские пирамидные клетки Беца. Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, а также от ба зальных ганглиев и мозжечка. Основной эфферентный выход двигательной коры на стволовые и спинальные моторные центры : формируют пирамидные клетки V слоя. Пирамидные и сопряженные с ними вставочные нейроны расположены вертикально по отношению к поверхности коры. Такие рядом лежащие нейронные комплексы, выполняющие сходные функции, называют функциональными двигательными колонками (рассмотренные выше). Пирамидные нейроны двигательной колонки могут возбуждать или тормозить мотонейроны стволовых и спинальных центров, например: иннервирующих одну мышцу. Соседние колонки в функциональном плане перекрываются, а пирамидные нейроны, регулирующие деятельность одной мышцы, расположены обычно не в одной, а в нескольких колонках.
Основные эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные пути, начинающиеся от гигантских пирамидных клеток Беца и менее крупных пирамидных клеток V слоя коры прецентральной извилины (60 % волокон), премоторной коры (20% волокон) и постцентральной извилины (20% волокон). Крупные пирамидные клетки имеют быстропроводящие аксоны и фоновую импульсную активность около 5 Гц, которая при движении увеличивается до 20 — 30 Гц. Эти «быстрые» пирамидные клетки иннервируют крупные (высокопороговые) &-мотонейроны в двигательных центрах ствола и спинного мозга, регулирующих физические движения. От мелких пирамидных клеток отходят тонкие медленнопроводящие миелиновые аксоны. Эти клетки имеют фоновую активность около 15 Гц, которая во время движения увеличивается или уменьшается. Они иннервируют мелкие (низкопороговые) &-мотонейроны в стволовых и спинальных двигательных центрах, регулирующих тонус мышц.
Пирамидные пути состоят из 1 млн волокон кортикоспинального пути, начинающихся от коры верхней и средней трети прецентральной извилины, и 20 млн волокон кортикобульбарного пути, начинающегося от коры нижней трети прецентральной извилины (проекция лица и головы). Волокна пирамидного пути оканчиваются на &-мотонейронах двигательных ядер III — VII и IX — XII черепных нервов (кортикобульбарный путь) или на спинальных двигательных центрах (кортикоспинальный путь). Через двигательную кору и пирамидные пути осуществляются произвольные простые движения и сложные целенаправленные двигательные программы, например профессиональные навыки, формирование которых начинается в базальных ганглиях и мозжечке и заканчивается во вторичной моторной коре. Большинство волокон пирамидных путей осуществляют перекрест, однако небольшая часть волокон остается неперекрещенными, что способствует компенсации нарушенных функций движения при односторонних поражениях. Через пирамидные пути осуществляет свои функции и премоторная кора: двигательные навыки письма, поворот головы, глаз и туловища в противоположную сторону, а также речь (речедвигательный центр Брока, поле 44). В регуляции письма и особенно устной речи имеется выраженная асимметрия больших полушарий мозга: у 95 % правшей и 70 % левшей устная речь контролируется левым полушарием.
К корковым экстрапирамидным путям относятся кортикорубраль-ные и кортикоретикулярные пути, начинающиеся приблизительно от тех зон, которые дают начало пирамидным путям. Волокна кортикорубрального пути оканчиваются на нейронах красных ядер среднего мозга, от которых далее идут руброспинальные пути. Волокна кортикоретикулярных путей оканчиваются на нейронах медиальных ядер ретикулярной формации моста (от них идут медиальные ретикулоспинальные пути) и на нейронах ретикулярных гигантоклеточных ядер продолговатого мозга, от которых начинаются латеральные ретикулоспинальные пути. Через эти пути осуществляется регуляция тонуса и позы, обеспечивающих точные целенаправленные движения. Корковые экстрапирамидные пути являются компонентом экстрапирамидной системы головного мозга, к которой относятся мозжечок, базальные ядра, моторные центры ствола; она осуществляет регуляцию тонуса, позы равновесия, выполнение заученных двигательных актов, таких как ходьба, бег, речь, письмо и др. Поскольку кортикопирамидные пути отдают многочисленные коллатерали структурам экстрапирамидной системы, обе системы работают в функциональном единстве, поэтому некоторые авторы (Р. Шмидт, 1985) считают нецелесообразным их разделение.
Оценивая в общем роль различных структур головного и спинного мозга в регуляции сложных направленных движений, можно отметить, что побуждение (мотивация) к движению создается в лимбической системе, замысел движения — в ассоциативной коре больших полушарий, программы движений — в базальных ганглидх, мозжечке и премоторной коре, а выполнение сложных движений происходит через двигательную кору, моторные центры ствола и спинного мозга (Р. Шмидт, 1985).
Межполушарные взаимоотношения
Межполушарные взаимоотношения у человека проявляются в двух главных формах — функциональной асимметрии больших полушарий и совместной их деятельности.
Функциональная асимметрия полушарий является важнейшим психофизиологическим свойством головного мозга человека. Исследование функциональной асимметрии полушарий началось в середине XIX в., когда французские медики М. Дакс (1836) и П. Брока (1861) показали, что нарушение речи человека возникает при поражении коры нижней лобной извилины, как правило, левого полушария. Несколько позже немецкий психиатр К. Вернике (1874) обнаружил в коре заднего отдела верхней височной извилины левого полушария слуховой центр речи, поражение которого приводит к нарушению понимания устной речи. Эти данные и наличие моторной асимметрии (праворукости) способствовали формированию концепции, согласно которой для человека характерно левополушарное доминирование, образовавшееся эволюционно в результате трудовой деятельности и являющееся специфическим свойством его мозга. Однако в XX в. в результате применения различных методических подходов, особенно при исследовании больных с расщепленным мозгом (перерезка мозолистого тела), было показано, что по ряду психофизиологических функций доминирует не левое, а правое полушарие. Таким образом возникла концепция частичного доминирования полушарий (Р. Сперри, 1981).
Выделяют психическую, сенсорную и моторную межполушарную функциональную асимметрии мозга. При исследовании психофизиологических функций было показано, что в речи словесный информационный канал контролируется левым полушарием, а несловесный канал (голос, интонация) — правым. Абстрактное мышление и сознание связаны преимущественно с левым полушарием. При выработке условного рефлекса в начальной фазе доминирует правое полушарие, а во время упрочения рефлекса — левое. Правое полушарие осуществляет обработку информации одновременно, синтетически, по принципу дедукции, лучше воспринимаются пространственные и относительные признаки предметов. Левое полушарие производит обработку информации последовательно, аналитически, по принципу индукции, лучше воспринимает абсолютные признаки предметов и временные отношения. В эмоциональной сфере правое полушарие обусловливает преимущественно более древние, отрицательные эмоции, контролирует проявления сильных эмоций, в целом оно более «эмоционально». Левое полушарие обусловливает в основном положительные эмоции, контролирует проявление более слабых эмоций.
В сенсорной сфере роль правого и левого полушарий лучше всего проявляется при зрительном восприятии. Правое полушарие воспринимает зрительный образ целостно, сразу во всех подробностях, легче решает задачу различения предметов и опознания визуальных образов предметов, которые трудно описать словами, создает предпосылки конкретно-чувственного мышления. Левое полушарие оценивает зрительный образ расчлененно, аналитически, при этом каждый признак (форма, величина и др.) анализируется раздельно. Легче опознаются знакомые предметы и решаются задачи сходства предметов, зрительные образы лишены конкретных подробностей и имеют высокую степень абстракции; создаются предпосылки логического мышления.
Моторная асимметрия выражается прежде всего в право-леворукости, которая контролируется моторной корой противоположного полушария. Асимметрия других групп мышц (например, толчковая нога) имеет индивидуальный, а не видовой характер.
Функциональная асимметрия больших полушарий, обеспечивая новый, более высокий уровень регуляции сложных функций мозга, одновременно повышает требования к совмещению деятельности двух полушарий.
Парность в деятельности больших полушарий обеспечивается наличием комиссуральной системы (мозолистого тела, передней и задней, гиппокампальной и хабенулярной комиссур, межталамического сращения), которые анатомически соединяют два полушария головного мозга. Результаты, свидетельствующие о переносе возбуждения из одного полушария в другое, получены преимущественно с использованием электрофизиологических методик и метода условных рефлексов.
Основные факты, полученные с помощью электрофизиологических методик, показали, что возбуждение из участка раздражения одного полушария передается через комиссуральную систему не только в симметричный участок другого полушария, но и в несимметричные участки коры (например, из зрительной коры одного полушария в моторную кору противоположного полушария). Морфологические исследования также подтвердили наличие (кроме поперечных волокон мозолистого тела, связывающих симметричные участки коры) продольных и вертикальных волокон, связывающих несимметричные корковые поля.
Заключение
Использование метода условных рефлексов показало, что если рефлекс вырабатывается, например, на тактильное раздражение кожи одной стороны тела, то он легко воспроизводится при таком же раздражении симметричных участков кожи другой стороны тела, т. е. в процессе выработки рефлекса происходит «перенос» временной связи в другое полушарие. Предварительная перерезка мозолистого тела прекращала (или сильно затрудняла) «перенос» условного рефлекса. Вместе с тем перерезка всех комиссур больших полушарий полностью не исключает возможность межполушарного взаимодействия при выработке временной связи. Эти результаты говорят о том, что элементарные формы взаимодействия двух полушарий могут осуществляться через четверохолмие и ретикулярную формацию ствола.
