
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
АДАПТИВНЫЙ ОТВЕТ ПРОРОСТКОВ ЛЬНА-ДОЛГУНЦА
НА ДЕЙСТВИЕ ОСТРОГО ГАММА - И УФ-С-ОБЛУЧЕНИЯ
Автори: , ,
Аннотация: Представлены результаты комплексного исследования радиоадаптивного ответа (РА), начиная от получения дозовых зависимостей действия гамма - и УФ-С-облучения на проростки льна-долгунца с целью определения адаптирующих и тестирующих доз и завершая изучением гистологических особенностей РА. Показан специфический и неспецифический РА и его связь с гормезисным действием адаптирующих факторов. Полученные результаты позволили обосновывать гипотезу о возможной роли надклеточных процессов в обеспечении радиоадаптации.
Анотація: Здійснено повний алгоритм дослідження радіоадаптивної відповіді, починаючи від отримання дозових залежностей дії гострого гамма-опромінення і УФ-С-опромінення на проростки льону-довгунця з метою визначення адаптуючих і тестуючих доз і завершуючи вивченням власне радіоадаптивної відповіді. На прикладі специфічної і неспецифічної радіоадаптивної відповіді показано її зв’язок із гормезисною дією адаптуючих чинників і вивчено гістологічні аспекти механізму радіоадаптації. На основі отриманих результатів обґрунтовується гіпотеза про можливу роль надклітинних (зокрема, проліферативних) процесів у забезпеченні радіоадаптації.
Abstract : Complete algorithm of research of radioadaptive answer is in-process carried out, starting from the receipt of dose dependences of action of acute gamma-irradiation and UV-C-irradiation on acrospires of long-stalk flax with the purpose of determination of adapting and testing doses, and completing the study actually of radioadaptive answer. On the example of specific and heterospecific radioadaptive response the conection with the hormetic action of adapting factors and the histological aspects of mechanism of radioadaptation were studied. On the basis of the received results a hypothesis is grounded about the possible role of supracellular (in particular, proliferative) processes in providing the radioadaptation.
Ключевые слова: лен-долгунец, УФ-С-облучение, гамма-облучение, радиогормезис, радиоадаптация.
Ключові слова: льон-довгунець, УФ-С-опромінення, гамма-опромінення, радіогормезис, радіоадаптація.
Keywords: long-stalk flax, UV-C-irradiation, gamma-irradiation, radiohormeis, radioadaptation.
Ключові слова: льон-довгунець, УФ-С-опромінення, гамма-опромінення, радіогормезіс, радіоадаптація.
Keywords: flax, the UV-C irradiation, gamma irradiation, radiogormezis, radioadaptation.
Введение
Биологическую адаптацию можно разделить на эпигенетическую, когда приспособление к условиям среды происходит в течение онтогенеза и которая обеспечивается эпигенетическими механизмами регуляции, и на генетическую, обеспечиваемую механизмами генетической изменчивости, наследственности и отбора [1]. Незначительные кратковременные изменения факторов внешней среды не приводят к заметным изменениям физиологических функций растений, что обусловлено их способностью сохранять относительно стабильное состояние при изменениях в окружающей среде, т. е. поддерживать гомеостаз. Однако резкое и долговременное влияние неблагоприятных факторов приводит к нарушению многих функций растений и даже к их гибели. При воздействии на растение неблагоприятных факторов (стрессоров) в нем возникает напряженное состояние, отклонение от нормы, т. е. стресс [2, 3].
После воздействия стрессора организм определенное время может находиться в одном из следующих состояний: а) оставаться на исходном уровне адаптированности, когда сохраняется его конститутивный уровень приспособленности - ординарная адаптация; б) на уровне повышенной резистентности – гиперадаптация (состояние эустресса по Г. Селье); в) на уровне пониженной резистентности – гипоадаптация (состояние дисстресса по Г. Селье).
Каков механизм изменения приспособленности организмов? Предполагается, что в организме под влиянием стрессора происходит переходный процесс [4], который имеет характер перерегулирования недовосстановления или сверхвосстановления) параметров биосистемы (например, скорость роста и развития, скорость деления клеток и т. п.). Восстановление исходных значений параметров является следствием работы биологических систем восстановления, которые функционируют на всех уровнях их интеграции. При этом возможно положительное перерегулирование (гиперкомпенсация), когда имеет место стимуляция параметров биосистемы, которые характеризуют ее жизнеспособность. В случае действия ионизирующей радиации говорят о радиоостимуляции или радиогормезисе.
Одним из возможных механизмов повышения исходного уровня устойчивости (гиперадаптации) является изменение количественных параметров функционирования объекта в рамках его качественной специфичности. Адаптирующие факторы, фактически, выявляют потенциал толерантности (адаптивный потенциал) биологических объектов, что может быть достигнуто, в частности, путем применения ионизирующего и УФ-излучения.
Ранее нами была показана прямая связь состояния гиперадаптированости растений к действию ионизирующего излучения с состоянием радиогормезиса по ростовым параметрам, что дало основание изучать механизмы адаптации путем изучения самого по себе радиогормезиса [5]. Одним из возможных механизмов гормезиса (а следовательно, и адаптации) является стимуляция гистологических параметров апикальной меристемы корня (пролиферативной активности меристематических клеток, объема меристемы, размера клеток), на что мы в первую очередь и обратили внимание в работе.
Методы и объекты
Определяли дозовую зависимость действия гамма-облучения на семена льна-долгунца Linumusitatissimum L., f. elongata по ростовым характеристикам проростков, полученных из облученных семян, гистологические (митотический индекс, объем меристемы, размер клеток) характеристики апикальной меристемы корней. Семена облучали на кобальтовой гамма-установке «Рокус» в дозах от 1 до 10 Гр при мощности 1,42 сГр/с. Облученные семена замачивали в течение 6 ч в отстоянной водопроводной воде в чашках Петри, раскладывали на стеклах, покрытых влажной фильтровальной бумагой и размещали в специальной влажной камере в термостате при 22 °С. Каждые сутки проростки фотографировали. Измерение длины корешков осуществляли с помощью компьютерной программы tps2. Ростовую активность проростков определяли путем расчета индекса роста (ИР) – отношение текущей средней длины корней проростков к их исходной средней длине. УФ-С-облучение проростков осуществляли с помощью ламп типа ОБМ-150м при мощности 0,5 - 3,4 Вт/м2.
Митотический индекс (МИ) определяли по общепринятой методике [6].
Адаптивный ответ изучали по традиционной схеме – тестирующей дозе (ТД) предшествовала (за 24 ч) адаптирующая доза (АД). Применение ТД позволяло выявить направление (знак) и величину модифицирующего резистентность влияния АД. Стимулирующую ростовые показатели дозу (радиогормезисную) принимали в качестве адаптирующей. В опытах были использованы следующие варианты: 1. Абсолютный контроль. 2. Вариант, на который действовала только ТД. 3. Вариант, на который действовала лишь АД. 4. Вариант, который сначала подвергали действию АД, а затем (после 24 ч) ТД (АД + ТД). Возможны несколько способов представления адаптационных эффектов: 1. Нормирование значений параметров всех вариантов (№ на соответствующие параметры контрольного варианта («абсолютный» контроль » - №/1, 2/1, 3/1, 4/1 - 1-й способ. 2. Нормирование значений параметров варианта «АД» на «контроль» (3/1) и варианта «АД + ТД» на «свой» контроль ТД (4/2). Этот способ позволяет определить, как АД влияет на контроль и как АД влияет на последствия применения ТД, - 2-й способ. 3. Нормирование варианта «ТД» на контроль (2/1) и варианта «АД + ТД» на вариант «АД» (4/3), что позволяет определить способ влияния АД на параметры дозовой зависимости (если она, конечно, получена), - 3-й способ. В работе использовали только первые два способа. Статистическая обработка результатов проводилась стандартными методами [7]. Далее обсуждаются только статистически достоверные результаты (р ≥ 0,05).
Результаты и обсуждение
Поскольку в работе использовали УФ-С-облучение, которое имеет относительно низкую проникающую способность в биологических тканях, т. е. невозможно было рассчитывать на достаточно полную реализацию адаптационного потенциала, как на это можно было бы рассчитывать в случае использования гамма-облучения, то необходимо было в максимальной степени оптимизировать условия проращивания семян и выращивания проростков. Кроме этого, экспериментальным воздействиям подвергали субпопуляции растений (если считать, что имеющиеся растительные материалы в виде семян или полученных из них проростков представляли собой искусственно созданную популяцию), так как не все семена прорастали, а из того, что прорастало, не всегда вырастали нормальные в морфологическом отношении проростки. Выбраковке подлежали проростки с механически поврежденными зародышевыми органами, с зонами бактериальной и/или грибной инфекции и т. д. С этой целью, во-первых, в опытах использовали более-менее здоровые проростки и, во-вторых, обращали внимание на плотность выращивания растений, что согласно нашим предварительным данным могло существенно повлиять на физиологические характеристики растений.
Известно, что внутривидовая конкуренция является жесткой, поскольку организмы одного вида потребляют одинаковые ресурсы из окружающей среды. Также они имеют идентичные болезни и паразитов, что при высокой плотности монокультуры приводит к самоизреживанию популяции для поддержания экологического равновесия. Эти механизмы необходимы в условиях избытка потомков в каждом поколении; под их влиянием выживают самые приспособленные организмы [8]. Результаты, представленные на рис. 1 указывают, что при большой плотности проростков возможно взаимное ростовое ингибирование соседних проростков (плотность посадки 15 шт. на стекло и более). Наибольшую стимуляцию проростки льна испытывают при плотности посадки 10 шт. на стекло, т. е. при расстоянии 8 - 9 мм между соседними проростками. Поэтому было решено в дальнейшем работать с этой плотностью.
Поскольку исследования адаптационного эффекта проводили по традиционной схеме, т. е. применению ТД предшествовала АД, то, во-первых, нужно было получить дозовые зависимости в интервале гормезисных и ингибирующих доз, во-вторых, проверить тождество гормезисных и адаптирующих доз и, в-третьих, получить собственно адаптирующие (гиперадаптирующие) эффекты. Причем последнее предполагалось сделать двумя способами - когда в роли АД выступала определенная доза гамма-облучения или определенная доза УФ-С-облучения.

Рис. 1. Зависимость ИР проростков льна от полученной дозы УФ-С-облучения.
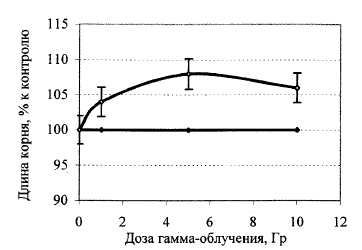
Рис. 2. Дозовая зависимость влияния острого гамма-облучения семян льна на длину корней 24-часовых проростков. Средние данные 9 экспериментов.
На рис. 2 представлены результаты изучения дозовой зависимости влияния острого гамма-облучения семян льна на длину корней 24-часовых проростков льна. Мы намеренно не использовали высоких доз, которые бы ингибировали ростовые параметры проростков, полученных из облученных семян, потому что гамма-излучение планировали использовать лишь как способ получения стимулирующих (гормезисных) эффектов. Представленные на рис. 2 оценки воздействий определенных доз являются обобщенными данными 10 идентичных по схеме и условиям проведения экспериментов. В дальнейшем дозу острого облучения 5 Гр как такую, которая давала самый большой и самый стабильный гормезисный (положительно стимулирующий) эффект, использовали как потенциально адаптирующую.
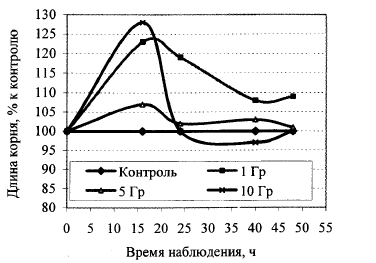
Рис. 3. Ростовая активность корней проростков льна-долгунца, полученных из гамма-облученных семян льна.
На рис. 3 можно видеть динамику ростовой активности корней проростков льна-долгунца, которые получены в одном из экспериментов, где использовали различные дозы острого гамма-облучения семян льна. Такой способ представления результатов позволяет прийти к выводу о временном (транзитивном) характере гормезисного действия облучения, который наблюдается только в определенном временном интервале. Если учитывать тот факт, что на первичном (физико-химическом) уровне гамма-облучение действует абсолютно деструктивно, в частности ионизируя атомы и молекулы, то стимуляцию ростовой активности мы должны рассматривать как компенсаторный, а точнее гиперкомпенсаторный процесс [4], который благодаря работе систем гомеостатической регуляции приводит, в конце концов, к возврату значений ростовых параметров до уровня соответствующих значений контрольных растений. Именно это является причиной транзитивности гормезисного эффекта. Конечно, его можно наблюдать только в определенном дозовом диапазоне и при определенных значениях мощности модифицирующего воздействия [9]. Вероятно, при более раннем наблюдении за ростовой активностью корней можно было бы наблюдать ее ингибирование. Именно потому, что основные гормезисные «события» происходят при данных условиях проведения экспериментов в течение 24 ч, мы в дальнейшем при изучении адаптивных реакций ориентировались именно на это время.
Подобные исследования были проведены при УФ-С-облучении с той лишь разницей, что облучали двухсуточные проростки льна, поскольку проникающая способность УФ-С-облучения недостаточна, чтобы зародыш в сухом семени испытал непосредственное влияние облучения. На рис. 4 видно, что примененный дозовый интервал позволил выявить как гормезисные эффекты, так и эффекты подавления ростовой активности. Следует заметить (см. также рис. 5) еще то, что временной интервал, в котором наблюдается гормезисный эффект короче (менее 20 ч), чем при наблюдении гормезисных эффектов у проростков, полученных при гамма-облучении семян. Объяснить это можно имеющейся разницей в физиологическом состоянии растений на момент облучения – покоящееся состояние зародыша при применении гамма-облучения и вполне физиологически активное состояние суточных проростков при применении УФ-С-облучения.
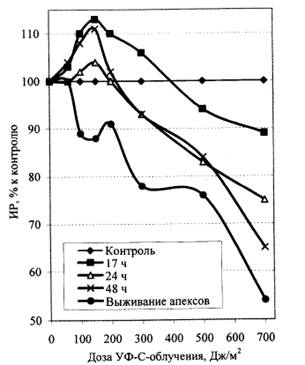
Рис. 4. ИР корней проростков льна най часы после УФ-С-облучения. 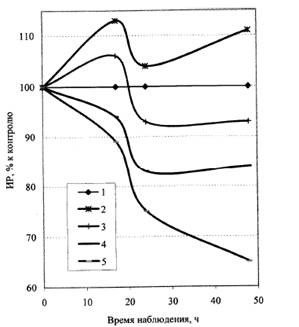
Рис. 5. Динамика влияния УФ-С-облучения на ИР корней проростков льна: 1 - контроль; 2 - 150 Дж/м2; 3 - 300 Дж/м2; 4 – 500 Дж/м2; 5 - 700 Дж/м2.
На рис. 4 представлена также дозовая зависимость выживаемости апикальной части зародышевых корней облученных проростков, определенная по необратимой остановке роста корня и побурению его апикальной части. Видно, что дозы в пределахДж/м2 является пороговыми с точки зрения возможности необратимого повреждения апикальной меристемы. Учитывая этот факт, при дальнейшем исследовании адаптивных реакций использовали в качестве адаптирующих дозы меньше 100 Дж/м2, хотя они и не были максимально гормезисными в отношении ростовой активности корней. Полученные в этих экспериментах данные позволили определить диапазон ТД – Дж/м2. По степени угнетения ростовой активности их можно считать оптимальными, так как меньшие дозы уменьшают модификационный интервал (разница величины эффекта между контрольным и опытным вариантами, выраженная, например, в процентах, изменение которой под влиянием модифицирующего фактора может быть принята за меру его влияния), а большие могут перевести систему в состояние необратимо ингибирования и эффект адаптирующего влияния не проявится.
Факт несовпадения дозовых зависимостей по ростовому параметру корня и по выживаемости апикальной части корня указывает на существенный аспект проблемы изучения механизмов адаптации (гормезиса). Дело в том, что теоретически и практически невозможно подвергнуть организм такому воздействию, чтобы получить гормезисные эффекты по всем возможным и существенным параметрам. Как считал [10], за адаптацию надо «платить». Иными словами, адаптация всегда фиксируется только по определенному параметру (или комплексу параметров), а по другим параметрам можно наблюдать даже дезадаптивные (гипоадаптивные) состояния или процессы, т. е. у адаптации есть своя «цена», что мы, собственно, и видели, сравнивая ростовые параметры и параметры выживаемости в одном и том же диапазоне УФ-С-облучения.
Динамика реагирования проростков льна на действие УФ-С-облучения (рис. 5) подобна динамике реагирования на действие острого гамма-облучения семян. Наблюдается постепенное возвращение ростовой активности стимулированных вариантов к контрольному уровню. Ингибирующие дозы действуют либо обратимо (дозы до 500 Дж/м2), когда угнетенная ростовая активность со временем восстанавливается, либо необратимо (за время наблюдения), когда степень угнетения постоянно растет (доза 700 Дж/м2).
После определения адаптирующих (фактически, потенциально адаптирующих) доз острого гамма-облучения, УФ-С-облучения и тестирующих доз УФ-С-облучения была осуществлена серия экспериментов по изучению радиоадаптации (гиперрадиоадаптации) проростков льна при действии гамма - и УФ-С-облучения. На рис. 6 представлены результаты одного из типичных по схеме проведения и полученным данным опыта по изучению неспецифической (перекрестной) радиоадаптации, т. е. такой, когда адаптирующий и тестирующий фактор имеют разную природу (в нашем случае – гамма-излучение и УФ-С-излучение соответственно). Как видно, тест-облучение применяли в тот момент, когда гормезисный эффект по ростовому параметру достигал максимального значения. Следует также отметить транзитивный характер самого адаптивного ответа. Об этом свидетельствует факт схождения кривых, описывающих динамику реагирования только на тест-дозу и на тестирующую дозу, которой предшествовало адаптирующее влияние. Результаты расчетов, представленные на рис. 6, сделаны первым из вышеуказанных способов и они позволяют определить абсолютное влияние различных схем облучения на ростовые параметры.

Рис. 6. Динамика ростовой активности облученных в разных режимах проростков льна (1-й способ расчета – все варианты в процентах к абсолютному контролю).
На рис. 7 представлены результаты четырех независимых опытов по изучению неспецифической (перекрестной) радиоадаптации, полученные при применении второго способа расчета радиоадаптационного эффекта. Видно, что резистентность корней проростков льна, полученных из предварительно облученных семян, возрастает на 10 %. Динамика адаптивного ответа напоминает динамику гормезисного действия облучения – ростовые параметры адаптированных вариантов постепенно сближаются с соответствующими параметрами неадаптированных, т. е. наблюдается транзитивность адаптивного ответа. Такой способ представления данных позволяет, фактически, при изучении радиоадаптивных реакций обходиться без абсолютного контроля. Особенно это важно тогда, когда экспериментатор имеет ограниченные ресурсы (время, материалы, объекты и др.). Однако если нас все же интересует механизм радиоадаптивного ответа, то эксперимент должен проводиться по полной, четырехвариантной схеме.
Результаты изучения специфического адаптивного ответа, т. е. такого, когда определенный фактор (в нашем случае УФ-С-облучение) вызывает повышение уровня резистентности к действию того же по своей природе фактора (в нашем случае опять же УФ-С-облучения) представлены на рис. 8. Можно констатировать факт качественного сходства этих данных с теми, что получены при изучении неспецифической адаптации. В частности, наблюдается та же транзитивность гормезисного действия облучения и относительно небольшое модифицирующее (в сторону повышения резистентности) действие предшествующего (адаптирующего) облучения.
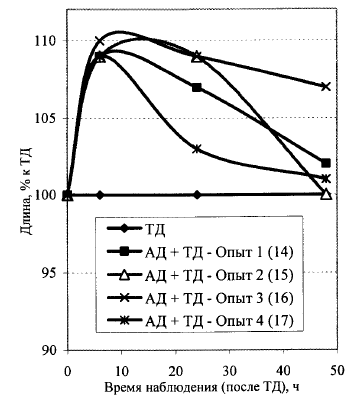
Рис. 7. Динамика ростовой активности облученных в разных режимах проростков льна (2-й способ расчета, процент к варианту ТД - 1000 Дж/м2), которые получены из предварительно облученных в дозе 5 Гр семян льна-долгунца.
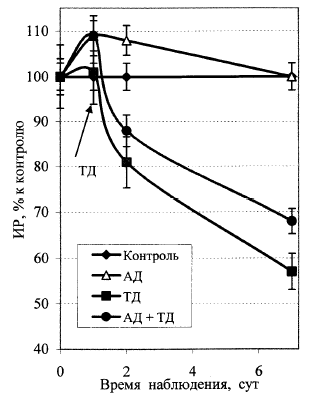
Рис. 8. Динамика ростовой активности корней проростков льна после применения разных схем УФ-С-облучения: контроль; АД – 19,8 Дж/м2; ТД –752,4 Дж/м2; АД + ТД.
Полученные результаты по изучению радиоадаптации ставят, как минимум, четыре вопроса.
Во-первых, какова возможная причина неспецифического действия определенных факторов, которые повышают резистентность к факторам другой природы? Объяснить это можно тем, что гамма - и УФ-С-излучение, несмотря на существенную разницу в энергии их квантов, которая делает первые (гамма-кванты) способными ионизировать атомы и/или молекулы облучаемого объекта, а вторые (кванты УФ-излучения) лишь возбуждать атомы, тем не менее имеют в качестве основной мишени своего действия молекулы ДНК. Кроме того, имеет место схожесть механизмов репарации повреждений ДНК, индуцированных этими видами излучения. Все это, вероятно, обусловливает сходство механизмов реагирования на гормезисные дозы каждого фактора, следствием чего является переход в состояние повышенной резистентности.
Во-вторых, почему радиомодифицирующее действие ограничивается десятью процентами и можно ли считать эту величину лишь частью адаптационного потенциала? Что касается реализации адаптационного потенциала, который выявляется с помощью УФ-С-облучения, то, действительно, учитывая относительно малую проникающую способность этого типа излучения, можно считать, что гормезисный, а следовательно, адаптационный потенциал полностью не реализуется. Увеличение дозы облучения в этом случае тоже проблему не решит, поскольку при этом будет увеличиваться доля необратимо поврежденных клеток.
В-третьих, можно ли продлить период пребывания в состоянии повышенной резистентности или, что то же самое, гормезисном состоянии? В наших опытах растения, подвергшиеся радиомодифицирующему влиянию и были затем подвергнуты «испытанию» тест-дозами, фактически в постфакторном периоде аходились в одинаковых условиях и, что главное, эти условия не были селективными, т. е. были близки к оптимальным как для адаптированных, так и для неадаптированных растений. Но если бы тест-дозы были близки к необратимому ингибированию, то даже небольшая разница в радиорезистентности, которая была приобретена благодаря гормезисному влиянию предварительного (перед тест-влиянием) облучения, могла способствовать большей выживаемости модифицированных растений, т. е. радиоадаптирующий эффект мог наблюдаться практически на протяжении всего онтогенеза. В противном случае зафиксировать состояние повышенной радиорезистентности можно было, создав такие условия в пострадиационном периоде (после адаптирующего воздействия), когда бы адаптированные растения смогли бы выжить лишь благодаря стимуляции ростовой активности. Например, это могло осуществитьсятогда, когда в реальных условиях открытого грунта корни стимулированных каким-то образом растений смогли достичь водоносного горизонта, а нестимулированные нет.
В-четвертых, почему наблюдается транзитивность радиоадаптивного (и, фактически, гормезисного) эффекта? Этот вопрос связан с предыдущим. Действительно, применение сублетальных тестирующих доз позволяет как модифицированным, так и немодифицированным растениям восстанавливать свои физиологические параметры до одинакового уровня, что и есть, фактически, проявлением транзитивности. Следует также учитывать то обстоятельство, что мы в эксперименте имеем дело с такой относительно мелкосемянной культурой, как лен, в этилированный проростках которой очень быстро исчерпывается запас питательных веществ. То обстоятельство, что мы зафиксировали гормезисный эффект по ростовым параметрам корневой системы проростков, вынудило нас сделать предположение относительно одного из промежуточных звеньев между облучением и конечным гормезисным эффектом. Таким звеном могла быть стимуляция деления клеток апикальной меристемы корня и/или стимуляция (ускорение) удлинения клеток зоны растяжения корня.
Таким образом, важно выяснить, была ли какая-то гистологическая предпосылка стимуляции ростовой активности корневой системы. Предполагалось, что в случае радиогормезисного эффекта по ростовым параметрам будет наблюдаться стимуляция митотической активности клеток корневой меристемы. При этом она должна была предшествовать повышению соответствующего параметра контрольного варианта.
Исследования с использованием цитологических параметров подтвердили наше предположение. В частности, установлено повышение митотической активности клеток апикальной меристемы первичных корешков 24-часовых проростков льна-долгунца, которые получены из острооблученных семян (таблица). Это сопровождалось также увеличением объема апикальной меристемы.
Гистологические параметры клеток апикальной меристемы первичных корней
24-часовых проростков льна-долгунца, полученных из остро-облученных семян
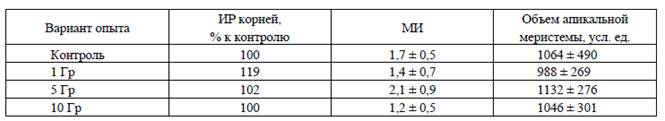
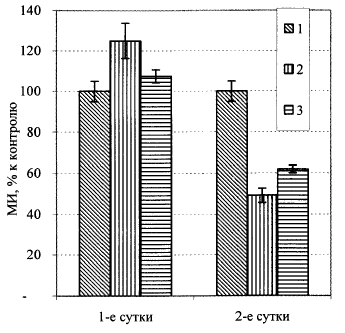
Рис. 9. Изменение МИ меристемы корня проростков льна в разное время после УФ-С-облучения стимулирующими дозами: 1 - контроль; 2 - 50 Дж/м2; 3 - 70 Дж/м2.

Рис. 10. Изменение объема апикальной меристемы корня проростков льна-долгунца в разное время после УФ-С-облучения стимулирующими дозами: 1 - контроль; 2 - 50 Дж/м2; 3 - 70 Дж/м2.
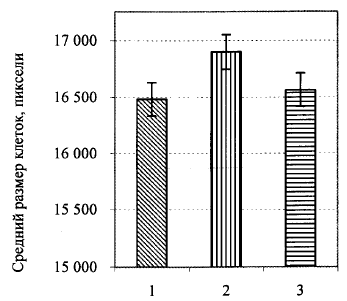
Рис. 11. Изменение размера клеток меристемы корня льна-долгунца на 1-е сутки после облучения АД: 1 - контроль; 2 - 50 Дж/м2; 3 - 70 Дж/м2.
Подобные данные были получены при действии гормезисных доз УФ-С-облучения (рисСледует обратить внимание на то, что увеличение МИ и объема меристемы происходит на фоне практически неизменного размера меристематических клеток, что указывает на реальное увеличение численности клеток в меристеме и что, в свою очередь, может сделать меристему (как критическую структуру) более резистентной (надежной) к действию стрессоров различной природы [11]. Конечно, такие рассуждения имеют смысл лишь с учетом того, что временной интервал между адаптирующим и тестирующим влиянием был достаточно большим (24 ч), чтобы задействовать именно указанные механизмы. При интервалах в несколько часов, скорее всего, будут работать модифицированные репарационные механизмы, которым в основном уделялось внимание при изучении радиоадаптивного ответа [12]. Таким образом, полученные результаты позволяют дополнительно обосновать гипотезу о существовании надклеточных механизмов радиоадаптации, в основе которых лежит стимуляция пролиферативной активности клеток, образующих критические ткани растения.
СПИСОК ЛИТЕРАТУРЫ
1. , Проблема фенотипического стресса и адаптации у растений // Физиология и биохимия культурных растений№ 2. - С.
2. На уровне целого организма. - М.: Наука, 19с.
3. , , Стресс у растений (биофизический подход). - М.: Изд-во Моск. ун-та, 19с.
4. Действие ионизирующего излучения на целостный растительный организм. - М.: Энергоатомиздат, 19с.
5. Міхєєв О. М., , І. Роль регенераційних механізмів в радіоадаптації проростків Pisum sativum L. // Вісник Харківського національного університету. Сер. БіологіяВипС.
6. Практикум по цитологии растений. - М.: Агропромиздат, 19с.
7. Программа STATISTICA для студентов и инженеров. - 2-е изд. - М.: Компьютер Пресс, 20с.
8. Элементы общей теории адаптации. - Вильнюс: Мокслас, 19с.
9. Концепция биологического действия малых доз ионизирующей радиации // Радиационная биология. РадиоэкологияТ. 35, № 5. - С.
10. Адаптация, стресс и профилактика. - М.: Наука, 19с.
11. Надежность растительных систем. - К.: Наук. думка, 19с.
12. Ikushima T., Aritomi H., Morista J. Radioadaptive response: Efficient repair of radiation induced DNA damage in adapted cells // Mut. ResVol. 358. - P.
