
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
На правах рукописи
СИНТЕЗ И ИЗУЧЕНИЕ СВОЙСТВ ОКСИЛИПИНОВ
ГРИБНОЙ И РАСТИТЕЛЬНОЙ ПРИРОДЫ
02.00.10 – Биоорганическая химия
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата химических наук
Москва-2007
Работа выполнена на кафедре Химии и технологии биологически активных соединений им. Московской государственной академии тонкой химической технологии им. и в лаборатории по исследованию эйкозаноидов отделения гинекологии университетского медицинского центра им. Бенджамина Франклина Свободного Университета (Берлин, Германия)
Научный руководитель доктор химических наук,
профессор
Официальные оппоненты доктор химических наук,
ведущий научный сотрудник
доктор биологических наук,
старший научный сотрудник
Ведущая организация НИИ Физико-химической биологии им.
Московского
государственного университета им.
Защита состоится «_24_» _декабря__2007 г. в _15_ часов на заседании Диссертационного Совета Д 212.120.01 при Московской государственной академии тонкой химической технологии им. Москва, пр. Вернадского, д. 86.
С диссертацией можно ознакомиться в библиотеке МИТХТ им. .
С авторефератом диссертации можно ознакомиться на сайте www. mitht. ru
Автореферат разослан «___» ноября 2007 г.
Ученый секретарь Диссертационного Совета,
кандидат химических наук,
старший научный сотрудник
Общая характеристика работы*
Актуальность проблемы. Микозы сопровождают человеческую цивилизацию не одно тысячелетие. Возбудителями микозов являются микроскопические грибы, одноклеточные или многоклеточные эукариоты – хемоорганогетеротрофы по способу питания. В настоящее время наблюдается значительный рост заболеваемости микозами из-за миграции населения и изменения образа жизни в индустриальных странах. Особенно ощутим вред микозов в трансплантологии, онкогематологии, неонаталогии, а наиболее распространенные возбудители микозов, такие как дрожжи Candida spp., уверенно вытесняют с лидирующих позиций внутрибольничных бактериальных возбудителей. Микозы лидируют в структуре СПИД-ассоциированных патологий, поражая организм, лишенный иммунной защиты. Для разработки эффективных подходов к терапии и диагностике микологических инфекций актуальным является фундаментальное изучение метаболических и сигнальных процессов, определяющих вирулентные свойства, механизмы развития и патогенеза болезнетворных грибов.
В последнее время интенсивно исследуется липидный метаболизм грибов с целью выявления потенциальных клеточных мишеней, знания о которых могут открыть перспективы создания новых фунгицидных или фунгистатических препаратов. Особенное внимание привлекают оксилипины, представляющие широкое семейство окисленных производных полиненасыщенных жирных кислот и выполняющие регуляторную и сигнальную функции в организме животных, растений и грибов. Среди них гидроксилипины – гидроксипроизводные жирных кислот – наиболее распространены в царстве грибов и играют важную роль в процессах роста и развития этих микроорганизмов (Tsitsigiannis et al., 2007). 3- и 18-Гидроксиполиеновые кислоты были обнаружены как продукты трансформации эндогенных и экзогенных жирных кислот в некоторых видах микроскопических грибов и дрожжей (Mucor genevensis, Dipodascopsis uninucleata, Lipomyces yarrowi, Candida albicans, Gaeumannomyces graminis), показано их участие в росте и морфогенезе грибов, а также в модуляции защитных реакций клеток млекопитающих (Deva et al., 2000). Для изучения процессов окисления липидных компонентов клеточных мембран растений и грибов, а также с целью выяснения механизмов действия биологически активных оксилипинов необходимы препаративные количества образцов этих соединений, что делает актуальным поиск эффективных методов их получения.
Оксилипины, являющиеся производными длинноцепных полиненасыщенных
_______________________
* В руководстве работой принимал участие старший научный сотрудник, кандидат химических наук
жирных кислот, достаточно трудно выделить из природных источников в количествах, необходимых для проведения биохимических исследований. Поэтому для изучения механизмов биосинтеза липидных метаболитов в грибах и трансдукции сигналов в животных клетках при микозном поражении актуальной задачей является разработка новых подходов к химическому (в том числе препаративному), ферментативному, микробиологическому синтезу 3-гидрокси - и 18-гидроксипроизводных жирных кислот, их предшественников и изотопно-меченных аналогов.
Настоящая работа является частью научных исследований, проводимых на кафедре Химии и технологии биологически активных соединений им. МИТХТ им. в рамках бюджетной темы № 1Б-4-355 «Синтез супрамолекулярных структур на основе порфиринов, липидов и углеводов с целью изучения процессов, протекающих в клетке и создания препаратов для онкологии, генной терапии и других областей медицины», по грантам РФФИ , , президента РФ по поддержке ведущих научных школ № НШ-2013.2003.3, а также частью работ, выполняемых в плане проекта по исследованию 3-гидроксиполиеновых жирных кислот в лаборатории эйкозаноидов кафедры гинекологии медицинского университетского центра Свободного Университета Берлина (Германия) при поддержке VW-фонда (грант Ni-1/77643).
Цель работы. Цель настоящей работы состояла в синтезе, наработке и изучении биохимических свойств новых и известных 3- и 18-гидроксилипинов природной структуры.
Основными задачами исследования явились:
1) разработка новых методических подходов к химическому и ферментативному синтезу, в том числе препаративному, 3- и 18-гидроксилипинов, их аналогов и меченных тритием производных;
2) изучение метаболизма полученных 3- и 18-гидроксиполиеновых кислот в дрожжах, характеристика профиля образующихся метаболитов и кинетики окисления гидроксикислот;
3) исследование ферментативных превращений синтезированных 3-гидроксилипинов и их аналогов с использованием оксигеназ млекопитающих, определение влияния 3-гидроксикислот на передачу сигналов в животных клетках на моделях хозяин-патоген.
Научная новизна.
- Разработаны новые схемы получения и осуществлен препаративный химический синтез:
а) оптически активных 3-гидроксилипинов: 3(R)-гидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (3(R)-НЕТЕ) и ее аналогов, 3(R)-гидрокси-(5Z,8Z)-тетрадекадиеновой кислоты (3(R)-HTDE) на основе полиацетиленовых предшественников с использованием оптически активных исходных синтонов;
б) 18-гидроксипроизводных арахидоновой кислоты: 18-гидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (18-НЕТЕ), 3(R),18(R/S)-дигидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (3,18-DiHETE) и их аналогов;
в) w-гидроксипроизводных линолевой и a-линоленовой кислот – 18-гидрокси-(9Z,12Z)-октадекадиеновой кислоты (18-HODE) и 18-гидрокси-(9Z,12Z,15Z)-октадекатриеновой кислоты (18-HOTrE).
- Впервые получены кратно-меченные тритием аналоги 3-НЕТЕ и 3,18-DiHETE.
- Разработан способ ферментативного получения дигидроксипроизводных ПНЖК на примере окисления 3-НЕТЕ соевой 15-LOX. Впервые получена 3(R),15(S)-дигидрокси-(5Z,8Z,11Z,13E)-эйкозатетраеновая кислота (3,15-DiHETE), подобраны условия для ее выделения с применением ВЭЖХ на хиральной фазе.
- Впервые установлено, что дрожжи Dipodascopsis uninucleata и условно-патогенный штамм Candida albicans способны синтезировать ди - и тригидроксиполиеновые продукты из экзогенных моногидроксиполиеновых жирных кислот. Определены кинетические параметры взаимодействия синтезированных жирнокислотных субстратов с клетками дрожжей C. albicans, показано, что 3-НЕТЕ является хорошим субстратом для C. albicans.
- Впервые показано, что 3-НЕТЕ и ее производные подвергаются ферментативной трансформации под действием липоксигеназ и циклооксигеназы млекопитающих с образованием метаболитов, аналогичных продуктам окисления арахидоновой кислоты.
- Установлена принципиальная возможность участия 3-гидроксикислот в трансдукции сигналов при взаимодействии хозяин-патоген, и впервые показано, что 3-НЕТЕ является модулятором клеточных сигналов и миметиком арахидоновой кислоты при взаимодействии с клетками млекопитающих.
Практическая значимость. Практическое значение представляют разработанные универсальные подходы к химическому синтезу ациклических производных природных полиненасыщенных кислот, основанные на ацетиленовой стратегии и позволяющие получать аналоги арахидоновой, линолевой, линоленовой кислот, в том числе радиоактивно-меченные и модифицированные заместителями и функциональными группировками с различной степенью гидрофильности, предназначенные для исследования механизмов биологического действия оксилипинов. Способы химического и ферментативного синтеза оксилипинов, а также методы очистки целевых продуктов, разработанные в ходе выполнения исследования, позволяют нарабатывать индивидуальные образцы этих соединений в количествах, достаточных для проведения биохимических исследований, важных для медицины. Полученные данные по изучению метаболизма и ферментативной трансформации оксилипинов природной структуры могут быть использованы при поиске новых малотоксичных соединений с потенциальной фармакологической активностью.
Основные положения, выносимые на защиту.
1. Разработка новых путей препаративного химического синтеза природных 3-и 18-гидроксилипинов, а также их аналогов и кратно-меченных тритием производных.
2. Разработка способа ферментативного получения 3,15-дигидроксиэйкозатетраеновой кислоты на примере окисления 3-НЕТЕ при помощи соевой 15-LOX, оптимизация условий выделения DiHETE с применением ВЭЖХ.
3. Изучение метаболизма синтезированных 3- и 18-гидроксилипинов в клетках дрожжей, выделение продуктов и исследование кинетики окисления гидроксикислот.
4. Исследование ферментативной трансформации синтезированных оксилипинов с использованием оксигеназ млекопитающих, выделение и идентификация продуктов оксигенации.
5. Изучение влияния 3-гидроксипроизводных жирных кислот на трансдукцию сигналов в животных клетках с применением известных биологических моделей и репортерных систем.
Публикации. Результаты работы отражены в 12 публикациях, в том числе в 3 статьях в зарубежных журналах, 1 статье в российском журнале и 8 тезисах докладов на международных и российских научных конференциях.
Апробация работы. Основные результаты работы представлены на Международном симпозиуме «Recent BioMedical Advances in Eicosanoid Research» (Берлин, 2002), на VIII Международной научно-технической конференции «Наукоемкие химические технологии 2002» (Уфа, 2002), на I Всероссийском конгрессе по медицинской микологии (Москва, 2003), на XVII Менделеевском съезде по общей и прикладной химии (Казань, 2003), на Международной конференции по физико-химической биологии, посвященной 70-летию со дня рождения академика (Москва, 2004).
Объем и структура работы. Диссертационная работа изложена на ____ стр. машинописного текста, содержит ____ рисунков, ____ таблиц, ____ схем, состоит из введения, литературного обзора, обсуждения результатов, экспериментальной части, выводов и списка литературы, включающего ____ ссылок.
Список сокращений. COX – циклооксигеназа;
DiHETE – дигидроксиэйкозатетраеновая кислота;
DMF – диметилформамид;
EET – эпоксиэйкозатетраеновая кислота;
GC-MS – хромато-масс-спектрометрия;
HETE – гидроксиэйкозатетраеновая кислота;
HTDE – гидрокситетрадекадиеновая кислота;
HODE – гидроксиоктадекадиеновая кислота;
HOTrE – гидроксиоктадекатриеновая кислота;
HMPA – гексаметилтриамид фосфорной кислоты;
LOX – липоксигеназа;
PGB2 – простагландин В2 (9-оксо-(15S)-гидрокси-проста-(5Z,8(12),13E)-триен-1-овая кислота);
PGE2 – простагландин E2 (9-оксо-(11a,15S)-дигидрокси-проста-(5Z,13E)-диен-1-овая кислота);
THF – тетрагидрофуран;
TMS – триметилсилильный;
TriHETE – тригидроксиэйкозатетраеновая кислота;
prСОХ-2-pLUC - репортерный вектор, содержащий промоторную конструкцию гена cox-2, лигированную с геном люциферазы;
PPARg D/N – плазмида, содержащая точечную мутацию в кодирующей области гена белка PPARg (изоформы рецептора PPAR);
IKK D/N – плазмида, содержащая точечную мутацию в IkB киназном домене IKK-белкового комплекса, ответственного за фосфорилирование процессингового протеина, обеспечивающего действие транскрипционного фактора NF-kB;
prNF-kB-pLUC – репортерный вектор, содержащий промоторный участок гена nf-kB транскрипционного фактора NF-kB, лигированный с геном люциферазы;
АК (АА) – арахидоновая кислота ((5Z,8Z,11Z,14Z)-эйкозатетраеновая);
ПНЖК – природные полиненасыщенные жирные кислоты.
Результаты работы и их обсуждение
Перспективность изучения влияния жирнокислотных метаболитов на трансдукцию сигналов и формирование ответа в клетке, низкое содержание их в природных источниках, отсутствие высокочистых стандартов, трудности биотехнологического получения, малоэффективность известных способов синтеза оксилипинов и изотопно-меченых аналогов стимулировали проведение исследований в области их химического и ферментативного синтеза. Для исследования механизмов ферментативной трансформации и биологических эффектов оксилипинов в клетках млекопитающих и грибов в качестве субстратов были выбраны жирнокислотные производные грибного происхождения и их аналоги: оптически активные R-энантиомеры – 3(R)-HTDE; 3(R)-НЕТЕ, 3(R),18(R/S)-DiHETE и их кратно-меченные тритием производные, а также 18-НЕТЕ – в качестве инструмента для изучения биосинтеза DiHETE. С целью исследования механизмов защиты высших растений от грибковых и бактериальных патогенов были синтезированы w-гидроксильные производные эндогенных полиненасыщенных кислот растительного происхождения: 18-HODE и 18-HOTrE.
1. Синтез 3- и 18-гидроксилипинов, их аналогов и меченных тритием производных
Особенности строения минорных природных полиеновых кислот и оксилипинов потребовали разработки новых универсальных схем их синтеза. Основные задачи химического синтеза ациклических полиненасыщенных соединений заданной структуры, в том числе изотопно-меченых, представлены на рис. 1.
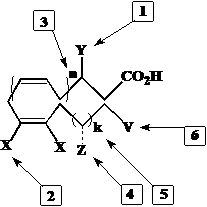
Рис. 1. 1 – стереоселективное введение (R)-гидрокси- или метокси-группы в положение 3; 2 – создание системы метиленразделенных цис-двойных связей с использованием водорода или трития; 3 – изменение количества двойных связей n = 1, 2, 3; 4 – введение гидроксигруппы в положение w-3; 5 – получение жирных кислот и их аналогов, отвечающих w3 или w6 природной структуре; 6 – введение гидроксильной группы в w-положение.
К настоящему времени имеется достаточное количество публикаций, посвященных синтезу гидроксилированных метаболитов арахидоновой кислоты. Так как в природных объектах найдены производные жирных кислот с различной конфигурацией центров асимметрии, подходы к их синтезу должны являться регио - и стереоспецифичными. Основной структурной особенностью природных эйкозаноидов этого класса является наличие системы метиленразделенных Z,Z- (ЕЕТ, НЕТЕ, 18/20-гидрокси-АК) или сопряженных E, Z-двойных связей (DiНЕТЕ). Существует три основных подхода к химическому синтезу ациклических производных ПНЖК. Первый способ («биомиметический») основывается на модификации природных эйкозаноидов, что требует выделения труднодоступных исходных соединений (Manna et al., 1983). Другим более общим и широко используемым способом получения является конденсация по Виттигу, с помощью которой были синтезированы 16-, 17- и 18-гидроксиэйкозатетраеновые кислоты (Heckman et al., 1996). Однако этот метод не дает возможности получения кратно-меченных изотопами аналогов оксилипинов, а также достаточной энантиомерной чистоты синтезируемых продуктов. В нашей лаборатории предложен третий метод создания цис-двойных связей при синтезе эйкозаноидов и родственных соединений, основанный на стереонаправленном восстановлении ацетиленовых предшественников ПНЖК (Иванов и др., 1995), который получил развитие в настоящей работе благодаря применению новых хиральных и бифункциональных ключевых синтонов и позволил получить кратно-меченные тритием оксилипины (схема 1). По данной методологии ключевая реакция кросс-сочетания сопровождается образованием высокореакционноспособных медных комплексов [RCºCCuX]-M+ терминальных ацетиленов без их предварительного металлирования и пропаргильных иодидов in situ, что позволяет проводить конденсации с высокими выходами (75-89%) в достаточно мягких условиях, а также применять в качестве пропаргильных компонентов тозилаты и галогениды. Также синтетическое исследование решало задачу расширения границ метода для создания функционализированных полиненасыщенных структур.
Схема 1

Для получения ациклических дигидроксиполиеновых кислот из монофункционализированных ПНЖК в данной работе были применены методы ферментативного синтеза, широко используемые в настоящее время для получения различных метаболитов длинноцепных полиненасыщенных жирных кислот.
1.1. Полный химический синтез 3-гидроксиполиеновых кислот и их аналогов
Ранее был осуществлен химический синтез 3(R)-HETE, основанный на трансформации 5,6-эпокси-АК (Bhatt et al., 1998). Однако этот метод не позволяет получать 3-гидроксипроизводные меченных или короткоцепных жирных кислот. Нами была разработана новая схема химического синтеза 3-гидроксиполиеновых соединений, основанная на полиацетиленовой стратегии, и получены оптически активные 3(R)-гидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновая (3(R)-HETE, 9а) (схема 2А) и 3(R)-гидрокси-(5Z,8Z)-тетрадекадиеновая (3(R)-HTDE, 14) кислоты (схема 2Б) природной структуры. В качестве исходного материала для получения ключевого хирального синтона 5 был выбран (R)-(-)-эпихлоргидрин (1), оксирановый цикл которого был раскрыт реакцией с этинилтриметилсилилбораном, образующимся из этинилтриметилсилана 2 под действием BuLi в присутствии эфирата трехфтористого бора (Yamaguchi et al., 1983). В результате удаления TMS защиты с ацетиленовой группы был получен хлоралкинол 3.
Схема 2
А

Б

Замещение хлора на цианогруппу давало цианид 4, омыление и последующее метилирование которого приводило к соединению 5. В соответствии с разработанной нами стратегией получение полиацетиленовых предшественников 3(R)-гидроксикислот 7 и 12 проводили с использованием кросс-сочетания оптически активного гидроксилсодержащего ацетиленового соединения – метил-3(R)-гидрокси-5-гексиноата (5) и пропаргильных бромидов: 1-бром-тетрадека-2,5,8-триина (6) или 1-бром-2-октина (11) с последующим гидрированием на катализаторе Линдлара и омылением метиловых эфиров 8 и 13. Синтезированные полиеновые соединения 8 и 13 были дополнительно очищены при помощи препаративной ВЭЖХ на обращенной фазе. Метоксильный аналог 3(R)-НЕТЕ (9б) был получен действием иодистого метила на полиен 8 в присутствии гидрида натрия с последующим щелочным омылением. Кратномеченный тритием аналог 3(R)-НЕТЕ – [5,6,8,9,11,12,14,15-3H]-3(R)-гидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновая кислота (10) – со специфической молярной радиоактивностью 1.65 Ки/ммоль была получена при использовании в процессе гидрирования полиинового предшественника 7 газовой смеси тритий/водород, содержащей 1% трития*, с последующим омылением (схема 2А). В ходе синтеза не наблюдалось рацемизации по С-3 положению, что было подтверждено анализом продуктов 9а, 9б, 14 при помощи ВЭЖХ на хиральной фазе. С целью оптимизации условий проведения ключевых реакций предварительно нами был получен рацемат 3(R/S)-HETE (9с) на основе rac-эпихлоргидрина.
* Получение меченных тритием 3-гидроксиполиеновых кислот проводили под руководством ведущего научного сотрудника лаборатории изотопно-модифицированных физиологически активных веществ Института молекулярной генетики РАН д. х.н.
1.2. Способ ферментативного получения 3(R),15(S)-дигидрокси-(5Z,8Z,11Z,13E)-эйкозатетраеновой кислоты (15а)
Для получения дигидроксипроизводных жирных кислот нами был разработан метод ферментативного синтеза 3,15-DiHETE (15а) с применением индивидуального препарата соевой 15-LOX-1 и 3(R)-HETE (9а) в качестве субстрата (схема 3). Концентрация субстрата (9а) была увеличена в 1.5 раза, а концентрация фермента – в 2 раза по сравнению с проводимыми ранее экспериментами по липоксигеназному окислению арахидоновой кислоты (Ivanov et al., 1998), так как 3-гидроксиарахидоновая кислота является худшим субстратом для липоксигеназы. Продукт ферментативной реакции 15а был выделен после очистки с применением ВЭЖХ на обращенной фазе с выходом 34% (образование сопряженного диена фиксировали при l 234 нм). Для дополнительного подтверждения энантиомерной чистоты 3,15-DiHETE был получен метиловый эфир (15б), подобраны условия проведения анализа этого соединения с применением ВЭЖХ на хиральной фазе: элюирование раствором гексан/этанол (10% об. этанола) давало одиночный пик (Rt =31.36 мин) с оптической чистотой более 96%. Структура соединения была подтверждена GC-MS анализом TMS-производного продукта 15б, согласно которому были идентифицированы ионные фрагменты (m/z): 175 [+CH(OTMS)CH2COOCH3], 173 [+CH(OTMS)(CH2)4CH3], а также молекулярный ион с m/z 479 [M+-15].
Схема 3
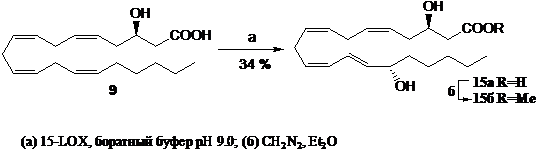
1.3. Химический синтез 18(R/S)-гидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (26), 3(R),18(R/S)-дигидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (29) и ее меченного тритием аналога (31)
Были разработаны новые универсальные подходы к химическому синтезу 18-гидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (18-НЕТЕ, 26) (схема 4), 3(R),18(R/S)-дигидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (3,18-DiHETE, 29) и ее кратно меченного тритием аналога 31 (схема 5) с использованием ацетиленовой стратегии. Получение рацематов 26 и 29 является приемлемым, так как разделение оптических изомеров возможно с помощью ВЭЖХ на хиральной фазе. В качестве исходного компонента при синтезе 18-НЕТЕ был выбран rac-1-хлор-3-пентанол (16), который превращали в иодид 17 по стандартной методике. Алкилирование этинилтриметилсилана 2 с использованием иодида 17 и BuLi в THF/HMPA приводило к образованию триметилсилильного производного гептинола 18. Кросс-сочетание бифункционального пропаргильного фрагмента 20 с бензоилированным производным 19 давало метиленразделенный трииновый спирт 21 с выходом 74%. Замена ОН-группы в соединении 21 на Br и кросс-сочетание бромида 22 с терминальным ацетиленовым производным 23 позволили получить тетраацетиленовый предшественник 24, последующее гидрирование которого на катализаторе Линдлара, а затем одновременное удаление бензоильной и метильной защитных групп в условиях щелочного гидролиза привели к 18(R/S)-гидроксиэйкозатетраеновой кислоте 26 (схема 4).
Схема 4

Синтез 3(R),18(R/S)-дигидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновой кислоты (29) и ее меченного тритием аналога 31 осуществлен исходя из одного тетраацетиленового предшественника 27, полученного реакцией кросс-сочетания метил-3(R)-гидрокси-гексиноата (5) и rac-3-бензоилокси-14-бромтетрадека-6,9,12-триина (22) в присутствии иодида меди (I). Стереоспецифическое гидрирование тройных связей соединения 27 на катализаторе Линдлара при соотношении долей катализатора и субстрата 2:1 по массе с последующей очисткой восстановленного продукта при помощи ВЭЖХ на обращенной фазе приводило к бензоил-замещенному тетраеноату 28 с выходом 64%. [5,6,8,9,11,12,14,15-3H]-3(R),18(R/S)-Дигидрокси-(5Z,8Z,11Z,14Z)-эйкозатетраеновая кислота (31) со специфической молярной радиоактивностью 1.7 Ки/ммоль была получена гидрированием соединения 27 в смеси водород/тритий (99:1), аналогично получению меченой 3(R)-НЕТЕ (10), с последующим удалением защитных групп.
Схема 5

Рацемат 29 был разделен на оптические изомеры с применением ВЭЖХ на хиральной фазе, индивидуальные R,R- и R,S-изомеры выделены с оптической чистотой более 97%. Как видно из рис. 2, разница во временах удерживания двух диастереомеров невелика. Однако данный метод позволяет получать количества оптически чистой DiHETE, достаточные для проведения биохимических исследований.
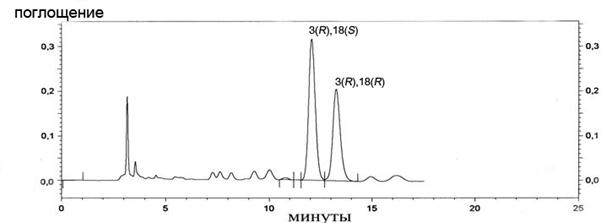
Рис. 2. ВЭЖХ метилового эфира 3(R),18(R/S)-DiHETE на хиральной фазе Chiral Pack AD (DAICEL, Франция), подвижная фаза: гексан/этанол (10% об. этанола), l 210 нм.
1.4. Синтез w-гидроксильных производных линолевой и a-линоленовой кислот
Нa основе полиацетиленовых предшественников была разработана универсальная схема получения 18-гидроксипроизводных природных полиненасыщенных жирных кислот: 18-гидрокси-(9Z,12Z,15Z)-октадекатриеновой кислоты (18-HOTrE, 39) и 18-гидрокси-(9Z,12Z)-октадекадиеновой кислоты (18-HODE, 40) (схема 6).
Схема 6

Полиацетиленовые предшественники 37 и 38 были получены с использованием реакции кросс-сочетания единого для двух полиненасыщенных кислот пропаргильного фрагмента 35 с 3,6-гептадиин-1-олом (34) и 6-гептин-1-олом (36), соответственно. 3,6-Гептадиин-1-ол (34), в свою очередь, получали с применением реакции кросс-сочетания 3-бутин-1-ола (32) и 1-тозилокси-3-триметилсилил-2-пропина (33) с последующим удалением триметилсилильной защиты. Каталитическое гидрирование метиловых эфиров полиацетиленовых кислот 37 и 38 на катализаторе Линдлара в присутствии хинолина и их последующее омыление приводило к образованию полиненасыщенных жирных кислот 39 и 40, соответственно (схема 6).
Структура всех полученных соединений была охарактеризована и подтверждена данными 1Н-, 13С-ЯМР-спектров, масс-спектров и элементного анализа.
Таким образом, была разработана синтетическая стратегия получения природных 3- и 18-гидроксилипинов, дигидроксиполиеновых кислот и их кратно-меченных тритием аналогов. Схемы синтеза позволяют получать ациклические производные жирных кислот с различной длиной цепи, варьировать функциональные заместители, количество и положение двойных связей путем подбора синтетических блоков с заданными структурными характеристиками. Оптимизация условий выделения оксилипинов, в том числе оптически активных, с помощью ВЭЖХ позволила получить эти соединения в препаративных количествах для использования в биологических исследованиях.
2. Изучение ферментативной трансформации, метаболизма 3- и 18-гидроксилипинов и их влияния на передачу сигналов в клетках млекопитающих*
3-Гидроксипроизводные жирных кислот играют важную роль в процессах адгезии, роста и репродукции клеток дрожжей. Вместе с тем остаются невыясненными как механизмы взаимодействия патогенных грибов с животными тканями, так и возможность трансформации грибных оксилипинов ферментами окисления липидов млекопитающих. Актуальным в этой связи является изучение метаболизма оксилипинов в грибах, продуктов взаимодействия оксилипинов с оксигеназами млекопитающих, оксилипин-ассоциированной трансдукции сигналов в животных клетках с целью выявления механизмов патогенеза болезнетворных грибов.
____________________________________________________________________________
* Эксперименты были выполнены в лаборатории по исследованию эйкозаноидов (рук. Ph. D. С. Нигам) отделения гинекологии университетского медицинского центра им. Бенджамина Франклина Свободного Университета Берлина. Культуры штаммов микроорганизмов дикого типа и мутантных были любезно предоставлены д-ром С. Нигамом.
2.1. Исследование ферментативной трансформации 3-гидроксиэйкозатетраеновой кислоты (9с) и ее аналогов с использованием липоксигеназ и циклооксигеназы млекопитающих
С целью выяснения возможности участия 3-гидроксикислот в ферментативном каскаде окисления липидов млекопитающих были проведены исследования по окислению 3(R,S)-HETE (9с) и ее метилового эфира очищенными ферментными препаратами липоксигеназ млекопитающих. Были использованы липоксигеназа лейкоцитов свиньи (l(12S)-LOX) и эпидермальная липоксигеназа ((12R)-LOX), полученная генноинженерным методом. Продукты окисления были выделены с помощью экстракции и последующей ВЭЖХ на обращенной фазе. Наличие 3-гидроксигруппы было установлено с применением хромато-масс-спектрометрии. Было обнаружено образование конъюгированного диена, 3,12-DiHETE (41) (рис. 3). Анализ дигидроксикислот с применением ВЭЖХ на хиральной фазе не показал предпочтительного окисления R- или S-энантиомера по С-3 положению жирной кислоты какой-либо из липоксигеназ (рис. 4).
Также были проведены исследования по взаимодействию 3(R)-метокси-ЕТЕ (9б) с овечьей циклооксигеназой COX-2. Продукты окисления были выделены при помощи известного метода (Hamberg et al., 1998) и затем разделены с применением ВЭЖХ на обращенной фазе. Структура полученных продуктов установлена с использованием GC-MS анализа метиловых эфиров их TMS-производных. Соединения были идентифицированы как простагландин 3(R)-метокcи-PGE2 (42) и 3(R)-метокси-11(S)-гидроксиэйкозатетраеновая кислота (43).
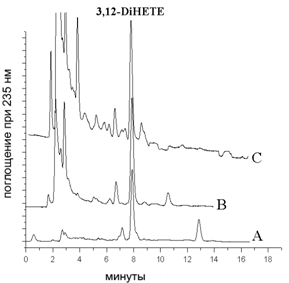
Рис. 3. Аналитическая ВЭЖХ на обращенной фазе продуктов окисления 3(R/S)-HETE под действием l(12S)-LOX (A) (продукт заметилирован перед проведением ВЭЖХ); метилового эфира 3(R/S)-HETE - под действием (12R)-LOX (B) и l(12S)-LOX (C), подвижная фаза MeOH/H2O/AcOH (80:20:0.1% об.)
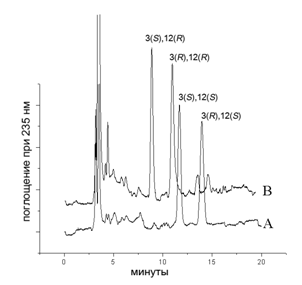
Рис. 4. ВЭЖХ на хиральной фазе заметилированных образцов 3,12-DiHETE, образованной действием l(12S)-LOX (A), (12R)-LOX (B), подвижная фаза гексан/этанол (10% об. этанола)
Щелочной гидролиз 3(R)-метокcи-PGE2 (42) приводил к продукту его дегидратации/изомеризации – циклопентеноновому простагландину 3(R)-метокcи-PGB2 (44), который легко детектировался при 278 нм (рис.5). Структура PGB2 подтверждена данными масс-спектра его TMS-производного.
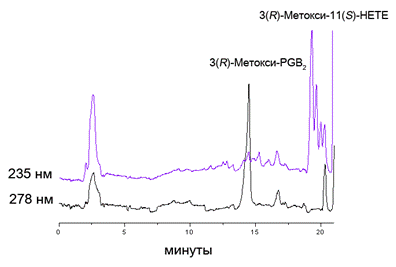
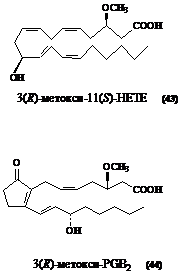
Рис. 5. Аналитическая ВЭЖХ на обращенной фазе продуктов окисления 3(R)-метокси-ЕТЕ (9б) (100 мкМ) под действием СОХ-2 млекопитающих (20 мкг), подвижная фаза CH3CN/H2O/AcOH (85:15:0.05 % об.)
Проведенные исследования показали, что 3-гидроксиполиеновые кислоты и их аналоги могут служить субстратами для ферментов окисления липидов млекопитающих – липоксигеназ и циклооксигеназ. При окислении 3-гидроксипроизводных жирных кислот образуются метаболиты, аналогичные оксигенированной АК, за исключением наличия 3-гидроксигруппы, что, вероятно, делает их узнаваемыми для клеток грибов. Таким образом, можно заключить, что 3-НЕТЕ и ее аналоги являются миметиками арахидоновой кислоты в процессах вторичного окисления. Вследствие доступности генноинженерных ферментов энзиматический синтез 3-гидроксиэйкозаноидов открывает перспективы для изучения потенциальной биологической активности данных соединений в стимуляции клеток млекопитающих в ответ на инфекционное поражение.
2.2. Исследование метаболизма 3(R/S)-НЕТЕ (9с) и 18(R/S)-НЕТЕ (26) в клетках дрожжей Dipodascopsis uninucleata и Candida albicans
Для выяснения путей метаболизма 3-гидроксиэйкозаноидов в клетках грибов было проведено исследование деградации рацемической 3(R,S)-НЕТЕ (9с) дрожжами Dipodascopsis uninucleata (UOFS Y 128). Клетки на репродуктивной стадии развития инкубировали с субстратом 3(R,S)-НЕТЕ, после чего определяли степень превращения R- и S-энантиомеров. Остаток 3(R,S)-НЕТЕ был выделен из супернатанта при помощи стандартных процедур с применением ВЭЖХ на обращенной фазе, и энантиомерный состав проанализирован с использованием ВЭЖХ на хиральной фазе. Результаты ВЭЖХ показали (рис.6), что преимущественному метаболизму в дрожжах Dipodascopsis uninucleata подвергается изомер 3(R)-HETE, в то время как 3(S)-HETE практически не расходуется.
Следующим этапом работы явилось изучение продуктов окисления АК и ее производных - 3(R,S)-НЕТЕ (9с), 18(R,S)-НЕТЕ (26) - в клетках Dipodascopsis uninucleata или Candida albicans 1386 (бронхоизолят), которые были инкубированы с жирнокислотными субстратами. Ранее 3,18-DiHETE была обнаружена на стадии развития гиф в дрожжах Candida albicans как продукт трансформации экзогенной АК (Deva et al., 2000).Нами были
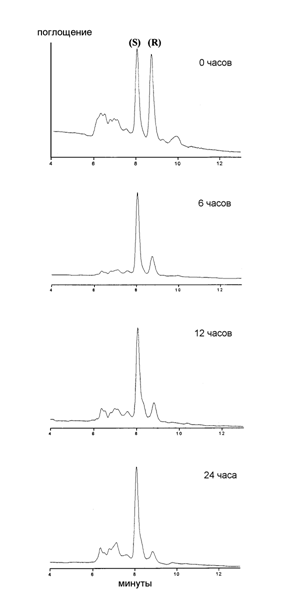
Рис. 6. Деградация 3(R/S)-HETE (9с) культурой клеток дрожжей Dipodascopsis uninucleata. ВЭЖХ на хиральной фазе (Chiral Pack AD) (CH3CN/H2O/H3PO4 95:5:0.1 % об.). Соотношение энантиомеров 3(S)/3(R) в %: после 6 ч 71/29; после 12 ч 78/22; после 24 ч 91/9.
проведены исследования на предмет обнаружения данного метаболита в разных типах дрожжей при добавлении 3-НЕТЕ в питательную среду. Продукты метаболизма дрожжей были выделены при помощи модифицированного (применением этилацетата в качестве растворителя для экстракции) метода (Bligh et al., 1959) с последующей хроматографией на обращенной фазе и препаративной тонкослойной хроматографией полярных компонентов на силикагеле. TMS-производные метиловых эфиров продуктов окисления были проанализированы с применением GC-MS на наличие 3-гидроксиэйкозаноидов. Результаты анализа ионных фрагментов позволили идентифицировать 3,18-DiHETE, 3,18,20-TriHETE (полученные в результате трансформации АК, 3-НЕТЕ и 3,18-DiHETE) как в дрожжах Candida albicans (А), так и в Dipodascopsis uninucleata (В) (рис.7). На наличие 3-гидроксиметаболитов были тестированы как клеточный лизат, так и супернатант; 3-оксилипины были обнаружены в каждой фракции. Важным представляется тот факт, что Di - и Tri-HETE экскретируются в межклеточное пространство и, по-видимому, участвуют в передаче клеточных сигналов.
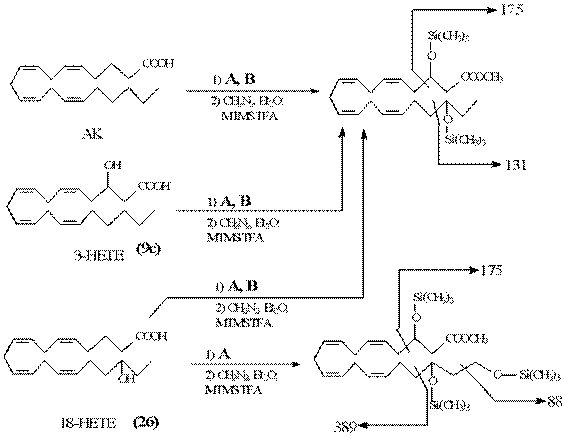
Рис.7. Продукты метаболизма АК, 3-НЕТЕ, 18-НЕТЕ в клетках дрожжей.
При сравнении масс-спектров 3,18-DiHETE, полученной микробиологическим синтезом, со спектром химически синтезированного стандарта 3,18-DiHETE (29) установлена их полная идентичность. В результате проведенных экспериментов можно сделать вывод о том, что (w-3)-гидроксилирование полиненасыщенных кислот является общим процессом для указанных типов дрожжей.
С целью определения скорости метаболизма жирнокислотных субстратов дрожжами Candida albicans оксиметрическим методом (электрод Кларка) была исследована кинетика общего поглощения кислорода клетками условно-патогенного штамма дикого типа 1386 в присутствии субстратов: АК, 3(R/S)-НЕТЕ (9с), 18(R/S)-НЕТЕ (26) в диапазоне концентраций от 5 до 20 мкМ.

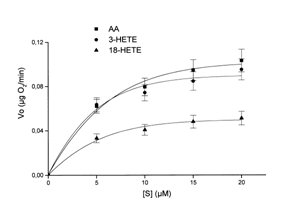
Рис. 8. Кинетические кривые зависимости скорости поглощения О2 клетками Candida albicans от концентрации жирнокислотных субстратов. Условия: 0,1 М фосфатный буфер, предварительно насыщенный О2. Значения Km и Vmax рассчитаны по уравнению Лайнуивера-Берка (1/[S] от 1/V), исходя из результатов 4 экспериментов.
Оказалось, что условные величины Кm для АК и 3(R/S)-НЕТЕ были одного порядка, тогда как для 18(R/S)-НЕТЕ условное значение Кm было в два раза выше, а скорость поглощения кислорода ниже (рис. 8). Можно предположить, что АК и 3(R/S)-НЕТЕ являются более предпочтительными субстратами для клеток гриба Candida albicans, чем 18(R/S)-НЕТЕ, и в первую очередь подвергаются клеточному метаболизму.
2.3. Изучение влияния 3-гидроксилипинов на передачу сигналов в клетках млекопитающих с применением известных репортерных систем*
Следующим этапом биологических исследований было изучение влияния 3(R/S)-НЕТЕ (9с) и 3(R)-HETE (9а) на трансдукцию сигналов в клетках человека. Известно, что _____________________________________________________________________________________* Плазмиды, содержащие конструкцию промотора гена СОХ-2, лигированного с геном люциферазы, и другие векторы, клонированные в E.coli, были любезно предоставлены научным сотрудником лаборатории по исследованию эйкозаноидов Свободного Университета Shankaranarayanan.
циклооксигеназы (COX-1, COX-2) катализируют ключевые реакции биосинтеза простаноидов. Внешние физиологически различные клеточные стимулы, такие как факторы роста, цитокины, регулируют уровень экспрессии генов cox-1 и cox-2. Повышение уровня экспрессии гена индуцибельной COX-2 сопровождает множество патологических и онкологических процессов: ревматоидный артрит, опухоли желудка, кишечника, молочной железы. С целью определения возможности модуляции активности гена cox-2 с помощью 3-гидроксилипинов было исследовано изменение уровня его экспрессии как под воздействием дрожжей Candida albicans, питавшихся субстратом АК, так и в присутствии индивидуальной 3-НЕТЕ в среде инкубации. В качестве экспериментального животного объекта была выбрана культура клеток человека эпителоидной карциномы матки – HeLa клетки, которая в комбинации с C. albicans является моделью вульвовагинального кандидоза.
HeLa клетки подвергали временной трансфекции: 1) репортерным вектором, содержащим конструкцию функциональной области гена cox-2, лигированного с геном люциферазы (prСОХ-2-pLUC), 2) либо аналогичной плазмидой, содержащую функциональную область гена транскрипционного фактора NF-kB (prNF-kB-pLUC).
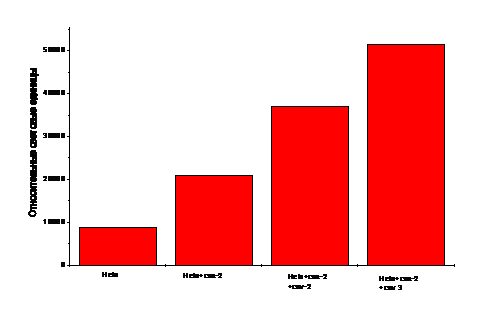
Рис. 9. Контроль - нетрансфицированные HeLa клетки. Мутанты cav2, cav3 (кратность инфекции =5) инкубировали с трансфицированными 1.5 мкг плазмиды -327/+59 prСОХ-2-pLUC HeLa клетками 5-6 ч при 37оС. Внутриклеточную среду анализировали по активности люциферазы-luc/субстрат (Promega) (относительные световые единицы).
Дополнительно применяли D/N плазмиды, содержащие нарушенные функциональные области генов: а) изоформы ядерного рецептора PPARg (PPARg D/N), б) IkB киназы IKK-киназного белкового комплекса, ответственного за фосфорилирование процессингового протеина NF-kB-зависимого сигнального пути (IKK D/N). Трансфицированные HeLa клетки были заражены C. albicans (ослабленные мутанты cav2 и cav3, питавшиеся АК) или обработаны 3(R/S)-НЕТЕ (9с), либо 3(R)-НЕТЕ (9а), затем внутриклеточная жидкость была исследована на люциферазную активность (счетчик-сцинтиллятор). Результаты эксперимента показали, что мутанты Candida albicans, питавшиеся АК, селективно повышали уровень экспрессии гена cox-2 в HeLa клетках (рис.9). Было выявлено, что 3(R,S)-HETE (9с), добавленная в инкубационную среду вместо инфекции, повышала уровень экспрессии гена cox-2 в дозозависимой манере (рис.10), как и было установлено ранее для АК. Аналогичный эксперимент с применением препарата оптически активного изомера 3(R)-HETE (9а) не показал изменений в уровне экспрессии гена cox-2. Опираясь на эти данные, можно сделать вывод о том, что энантиомер 3(R)-HETE (9а) не влияет на уровень экспрессии гена cox-2 в клетках млекопитающих.
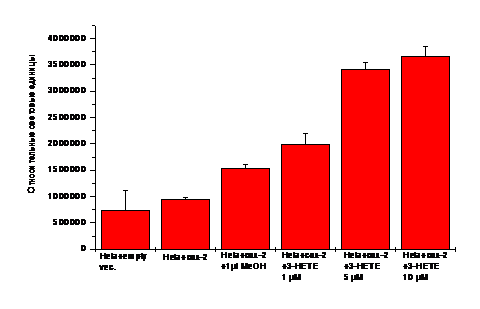
Рис. 10. Контроль - HeLa клетки, трансфицированные пустым люциферазным вектором pGL3. 3(R/S)-HETE в кнцентрациях 1, 5, 10 мкМ инкубировали с трансфицированными 1.5 мкг плазмиды-327/+59 prСОХ-2-pLUC HeLa клетками 2 ч при 37оС. Внутриклеточную среду анализировали по активности люциферазы - luc/субстрат (относительные световые единицы).
Каскад окисления полиненасыщенных жирных кислот вовлечен в такой важный физиологический ответ организма, как развитие воспаления. Семейство ядерных рецепторов, в которое входит PPAR (рецепторы, активируемые веществами, вызывающими пролиферацию пероксисом), является связующим звеном между регуляцией транскрипции некоторых генов, клеточными ответами и производными жирных кислот. PPARg – одна из изоформ ядерного рецептора PPAR. Поэтому представлялось интересным изучение влияния оксилипинов на PPARg-ассоциированный путь клеточных сигналов в СОХ-2-зависимом ответе HeLa клеток, инфицированных ослабленной Candida albicans. Результаты экспериментов по котрансфекции HeLa клеток плазмидой prСОХ-2-pLUC и плазмидой PPARg D/N показали, что ядерный рецептор PPARg участвует в СОХ-2-зависимых сигналах трансдукции, вызываемых мутантом дрожжей Candida albicans cav2, питавшимся АК. Инфекция не повышала уровня экспрессии СОХ-2 в присутствии плазмиды PPARg D/N (рис.11).
В данной работе проводились исследования по изучению влияния 3-гидроксилипинов на сигнальные внутриклеточные механизмы, ассоциированные с действием фактора NF-kB. Последний представляет собой редокс-чувствительный гетеродимерный транскрипционный фактор, связанный с пролиферацией, апоптозом, воспалением, иммунным, стрессовым ответом и другими регулируемыми процессами клеток млекопитающих.
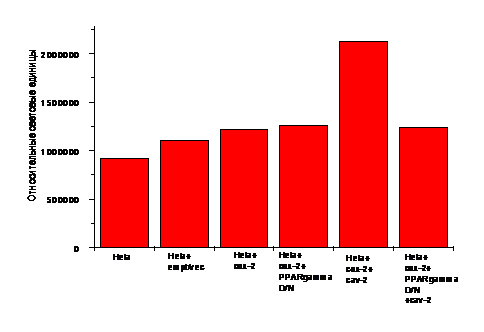
Рис. 11. Контроль - нетрансфицированные HeLa клетки; HeLa клетки, трансфицированные пустым вектором pGL3. Мутант cav2 (кратность инфекции =5) преинкубировали с трансфицированными 1.5 мкг плазмиды -327/+59 prСОХ-2-pLUC и 1.5 мкг плазмиды PPARg D/N HeLa клетками 2 ч при 37оС. Внутриклеточную среду анализировали по активности люциферазы - luc/субстрат (относительные световые единицы).
Эксперименты по котрансфекции HeLa клеток люциферазным вектором (prNF-kB-pLUC) и плазмидой IKK D/N показали, что 3(R/S)-НЕТЕ (9с) влияет на регуляцию экспрессии гена транскрипционного фактора NF-kB. В то же время данное производное жирной кислоты не повышало уровня экспрессии гена nf-kb в присутствии плазмиды IKK D/N (рис.12).
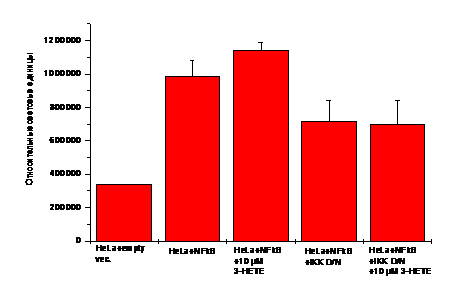
Рис. 12. Контроль - HeLa клетки, трансфицированные пустым вектором pGL3, HeLa клетки, трансфицированные prNF-kB-pLUC. 3(R/S)-HETE в концентрации 10 мкМ инкубировали с трансфицированными 1.5 мкг плазмиды prNF-kB-pLUC и плазмидой 1.5 мкг IKK D/N HeLa клетками 2 ч при 37оС. Внутриклеточную среду анализировали по активности люциферазы - luc/субстрат (относительные световые единицы).
Таким образом, было показано, что и Candida albicans, питавшаяся арахидоновой кислотой, и 3-гидроксилипины, добавленные в среду инкубирования, способны модулировать передачу сигналов в клетках млекопитающих на моделях кандидоза и, следовательно, могут участвовать в формировании воспалительного ответа по сценарию прооксидативного стресса, как и арахидоновая кислота. Также было показано, что механизмы, связанные с ядерным рецептором PPAR и транскрипционным фактором NF-kB, влияют на передачу сигналов, вызываемых микозной инфекцией или 3-гидроксиполиеновыми кислотами в животных клетках.
Выводы
• 1. Разработаны новые схемы препаративного химического синтеза:
• а) оптически активных 3-гидроксилипинов: 3(R)-HTDE, 3(R)-НЕТЕ, 3(R),18(R/S)-DiHETE с использованием хиральных исходных синтонов. Впервые получены кратно меченные тритием аналоги 3(R)-НЕТЕ и 3(R),18(R/S)-DiHETE;
• б) 18-гидроксипроизводных полиненасыщенных жирных кислот: 18(R/S)-НЕТЕ, 18-HODE, 18-HOTrE.
• 2. Разработан способ ферментативного получения дигидроксильных ПНЖК на примере окисления 3-НЕТЕ соевой 15-LOX. Впервые получена 3(R),15(S)-DiHETE, подобраны оптимальные условия для ее выделения с применением ВЭЖХ на хиральной фазе.
• 3. Впервые показано, что дрожжи Dipodascopsis uninucleata и Candida albicans способны синтезировать ди - и тригидроксиполиеновые продукты из моногидроксипроизводных полиненасыщенных жирных кислот. Определены кинетические параметры взаимодействия синтезированных жирнокислотных субстратов и клеток дрожжей Candida albicans, установлено, что 3-НЕТЕ является хорошим субстратом для клеток C. albicans.
• 4. Впервые показано, что 3-НЕТЕ и ее производные подвергаются ферментативной трансформации под действием липоксигеназ и циклооксигеназы млекопитающих с образованием оксигенированных метаболитов – аналогичных продуктам окисления арахидоновой кислоты.
• 5. Выявлено, что 3-гидроксиполиеновая кислота селективно модулирует передачу клеточных сигналов в модельных системах хозяин-патоген. Впервые показано, что при взаимодействии с клетками млекопитающих 3-НЕТЕ является миметиком арахидоновой кислоты и действует как провоспалительный агент.
Список работ, опубликованных по теме диссертации
1. Groza N. V., Ivanov I. V., Romanov S. G., Myagkova G. I., Nigam S. A novel synthesis of 3(R)-HETE, 3(R)-HTDE and enzymatic synthesis of 3(R),15(S)-DiHETE // Tetrahedron. – 2002. - V. 58. - P. .
2. Romanov S. G., Ivanov I. V, Groza N. V., Kuhn H., Myagkova G. I. Total synthesis of (5Z,8Z,11Z,14Z)-18- and 19-oxoeicosa-5,8,11,14-tetraenoic acids // Tetrahedron. – 2002. – V. 58. – P. .
3. Groza N. V., Ivanov I. V., Romanov S. G., Shevchenko V. P., Myasoedov N. F., Myagkova G. I., Nigam S. Synthesis of tritium labelled 3(R)-HETE and 3(R),18(R/S)-DiHETE through a common synthetic route // J. Labelled Comp. Radiopharm. – 2004. – V. 47. – P. 11-17.
4. , , Мягкова синтез омега-гидроксипроизводных растительных жирнокислотных субстратов // Вестник МИТХТ. – 2006. – Т. 1, № 4. – С. 29-32.
5. Groza N. V., Ivanov I. V, Romanov S. G., Myagkova G. I, Nigam S. Chemical synthesis of fungal and mammalian hydroxy eicosatetraenoic acids // Recent BioMedical Advances in Eicosanoid Research: book of abstracts of International Symposium. – Berlin. – 2002. – P. 57.
6. , , Мягкова полиненасыщенных жирных кислот микроорганизмов // Тезисы докладов III съезда Биохимического Общества. – Санкт-Петербург. – 2002. – С. 332.
7. , , Мягкова синтез различных 3(R)-гидроксиоксилипинов // Тезисы докладов VIII Международной научно-технической конференции «Наукоемкие химические технологии 2002». – Уфа. – 2002. – С. 88-89.
8. , , , Мягкова гидроксипроизводных арахидоновой кислоты и их аналогов // Тезисы докладов XVII Менделеевского съезда по общей и прикладной химии. – Казань. – 2003. – С. 222.
9. , , Мягкова моногидроксипроизводных полиненасыщенных жирных кислот в грибах // Сборник «Успехи медицинской микологии» по материалам I Всероссийского конгресса по медицинской микологии. – Москва. – 2003. – Т. I. – С. 277-278.
10. , , Мягкова биологически активных метаболитов грибкового происхождения – HETE(s), DiHETE(s) – и их изотопно меченных аналогов // Сборник «Успехи медицинской микологии» по материалам II Всероссийского конгресса по медицинской микологии. – Москва. – 2004. – Т. III. – С. 142-143.
11. , , Мягкова грибковых бета-гидрокси-полиненасыщенных жирных кислот с ферментами окисления липидов // Тезисы докладов Международной конференции по физико-химической биологии, посвященной 70-летию со дня рождения академика . – Москва. – 2004. – С. 96.
12. , , Мягкова методов химического синтеза аналогов субстратов окисления липидов мембран растений // Тезисы докладов X Международной научно-технической конференции «Наукоемкие химические технологии XXI века». – Волгоград. – 2004. – Т. 1. – С. 243-244.
Подписано в печать _______________ Формат 60х84/16. Бумага писчая.
Отпечатано на ризографе. Уч. изд. Листов 1.0. Тираж 100 экз.
Заказ № ________________
Московская государственная академия тонкой химической технологии
им.
Издательско-полиграфический центр
г. Москва, прос. Вернадского, 86
