
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
У осетровых рыб прежде не рассматривались вопросы пластичности поведенческих функций, отнесенных к раннему периоду онтогенеза.
3.1. Изменение характера поведения молоди осетра в зависимости от предварительной адаптации в поле химического модельного стимула (морфолина)
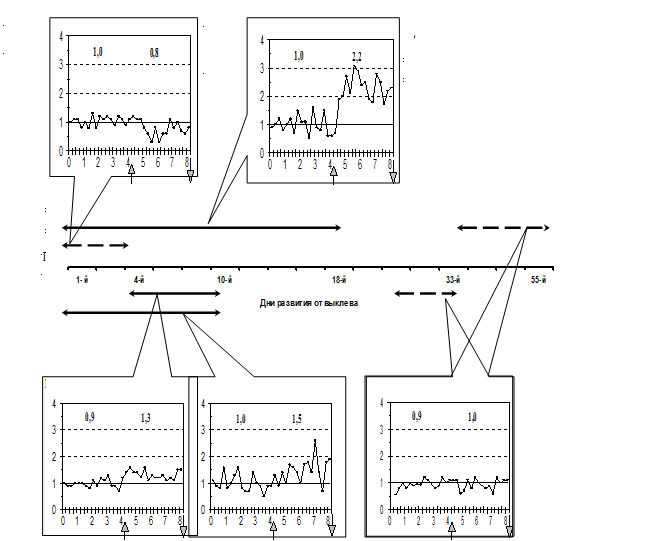
Качественное и количественное выражение реакции молоди на химический сигнал зависело от сроков и длительности предварительной адаптации этому сигналу, а также от его интенсивности. При минимальном сроке адаптации (с 1 по 4-й дни после выклева) реакция составляла 93% от фона (p <0,05) (избегательная реакция), что аналогично неадаптированному контролю (81%). Сдвиг реакции в сторону привлечения, по сравнению с фоном, обнаружен у личинок, адаптированных с 6-го по 10-й дни (134%), а также при увеличении срока адаптации до 10 и 18 дней, начиная от выклева – соответственно 151% и 224% (p < 0,001). Интенсивность реакции на концентрацию 10-9 М была выше, чем на 10-8 М. Различия в интенсивности поведенческого ответа на разные концентрации сигнала наиболее значительны (на 86%) при самой продолжительной адаптации к химическому сигналу (p < 0,001). Результат адаптации к химическому сигналу после 25-го дня развития, независимо от срока выдерживания личинок при тестировании был оценен как привыкание: поведение молоди на стимульном 104% (8 дней) и 105% (30 дней) и фоновом (100%) интервалах не различалось (Рис. 4).
Реакция предпочтения химического фона рыб, зависит как от внешних, так и гормональных влияний (Hasler, Scholz, 1983; Nevitt, Dittman, 1994). Это обусловило направление дальнейших исследований, в которых поведение, зависимое от предварительной адаптации к химическому сигналу, изучали при специфическом и неспецифическом воздействии химических агентов, вызывающих изменение уровня тиреоидных гормонов в период адаптации.
|
|
|
Рис. 4. Динамика двигательной реакции молоди осетра в поле химического стимула (морфолин, 10-8 - М 10-9 М) после адаптации личинок к стимулу в различные сроки раннего онтогенеза.
3.2. Оценка запечатления химического стимула личинками осетра с различным тиреоидным статусом
Уровни гормонов у личинок осетра изменяли, воздействуя тиомочевиной или тиомочевиной и тиреоидными гормонами (Т3) в течение 14-ти дней в чувствительный (1-й - 14-й дни) или нечувствительный (20-й - 35-й дни) периоды. В результате воздействия ингибитора тироксин снизился по сравнению с контролем, в среднем, на 22% (25,4 - 18,6 %), трийодтиронин - на 75% (86,4 - 63,1%). Добавление в воду Т3 привело к увеличению тканевого содержания Т3 на стадии активного питания на 200 и 220%.
Вызванное «гипотиреоидное» состояние в чувствительный период в значительной степени снизило положительную реакцию (120%). Если же в период адаптации личинок к химическому фону гормональный фон повышали с помощью Т3, то в этом случае по интенсивности реакция была аналогична реакции контрольных рыб и достоверно (p < 0,001) отличалась от реакции «гипотиреоидных» рыб (181%).
Реакция рыб зависела от уровня тиреоидных гормонов не только на предличином этапе развития, но и в более поздний период. Повышение гормонального фона у старшевозрастных личинок (с 20-го по 35-й дни) при их адаптации к химическому фону среды приводила при тестировании к предпочтению этого фона. Интенсивность реакции составила 161% (p < 0,001).
3.3. Влияние линдана и водорастворимых фракций нефти на поведенческую реакцию молоди осетра, связанные с «химическим опытом» в ходе личиночного развития
Воздействие линданом в концентрациях (0,1 мг/л и 0,5 мг/л) и нефтью (5 мг/л) в чувствительный для запоминания химических стимулов период приводило к тому, что рыбы демонстрировали индифферентную реакцию. В случае воздействия любого из токсикантов в нечувствительный период положительная реакция сохранялась, хотя ее интенсивность была меньше контрольной. После воздействия линданом интенсивность реакции составила 123%, что существенно отличалось от поведения после воздействия в чувствительный период (p < 0,001); после воздействия нефти - 120% (p <0,05). Таким образом, тестирование молоди, обработанной токсикантами в различном возрасте, показало необратимость повреждающего воздействия линдана и нефти в предличиночный период развития в сравнении с более поздним периодом.
Заключение
1. Впервые биохимическими методами показано наличие материнских тиреоидных гормонов у осетровых рыб (осетра и севрюги) в генеративной, эмбриональной и личиночной ткани. Не исключено, что уровни и соотношение форм гормонов в ооцитах должны отражать экологию вида (Tagawa et al., 1990; Mellinger, 1994), что подтверждают наши данные: соотношение форм гормона, при котором содержание Т4 превышает Т3, характерно для пресноводных видов с относительно невысокой скоростью развития, в том числе, для осетра и севрюги. Эти виды не различались по абсолютным величинам гормонов в яйцеклетках в расчете на единицу массы, что, вероятно, обусловлено общностью условий обитания, диктующих определенный уровень метаболизма. Можно ожидать, что у данных видов осетровых, но из других популяций, гормональные характеристики будут несколько иными.
2. Анализ показателей тиреоидных гормонов в генеративной ткани, эмбрионах и личинках в сопоставлении с морфологическими и физиологическими данными позволил с количественной стороны оценить их функциональное значение в эмбриогенезе. Нами установлено, что поступление Т4 в генеративную ткань самок осетра активизируется на III стадии зрелости и не прекращается в завершающий период созревания, ведущий к овуляции (V стадия). Снижение варьирования показателей по мере продвижения стадий эмбрионального развития свидетельствует о «стабилизации» эндокринного фона, обеспечивающего нормальное развитие зародышей лишь в определенном, более узком диапазоне величин и «отбраковку» уклонений. Уточнить сроки включения материнских тиреоидных гормонов в метаболизм зародыша удалось при сравнении динамики гормональных показателей в потомствах с высокой и низкой степенью оплодотворяемости икры (осетр), а также при аномальном развитии (севрюга). Расходование гормонального «запаса» начинается не позднее периода закладки нервных валиков, что подтверждено позднейшими исследованиями на рыбах с привлечением молекулярно-генетических методов (Liu, Chan, 2002). Рост показателей тиреоидных гормонов у эмбрионов на стадии выклева и выклюнувшихся особей предполагает возможность внефолликулярного синтеза тиреоидных гормонов в конце эмбрионального периода. Вторая фаза повышения Т4, наблюдаемая у личинок осетра при завершении периода желточного питания, очевидно, зависит от фолликулярного синтеза, поскольку совпадает по времени с первыми признаками активности щитовидной железы предличинок по гистологическим критериям [3, 8, 11,12, 18, 19,20].
3. В условиях хронического воздействия на предличинок осетра токсикантов, принадлежащим к различным классам - пестицида линдана и нефти - выявлена одинаковая по характеру реакция организма: фазовое смещение максимума Т4 со стадий 43-44 на 39-40 стадии, как свидетельство преждевременной активизации щитовидной железы. В поздние сроки развития предличинок снижение концентрации Т4 и перераспределение соотношения Т3/Т4, видимо, отражает нарушение функционирования различных звеньев тиреоидной оси, связанных с производством, конверсией и включением гормонов в метаболизм. Изучение динамики гормона стресса кортизола в тканях личинок осетра не только позволило уточнить биохимическими методами сроки активизации интерреналовой ткани, отнесенных к первым дням развития (Баранникова, 1974; 1975), но и показало согласованный характер изменений тироксина и кортизола в предличиночный период с одновременным максимумом, что свойственно «критическим периодам» метаморфоза рыб. Эти данные, а также увеличение амплитуды и изменение фазы волны кортизола (смещение на более ранние сроки развития) при специфическом хроническом воздействии, вызывающем дефицит тиреоидных гормонов (тиомочевина), появление фазового сдвига на эти же сроки Т4 в условиях хронического воздействия других стрессорных агентов - антропогенных поллютантов, предполагает взаимодействие тиреоидной и глюкокортикоидной осей у осетра уже в предличиночный период [12,20, 21].
4. У потомств различного качества отклонения от «нормы» в уровнях гормонов на стадии оплодотворения (Т4 и Т3 до 50%), выклева (Т3 до 50 %) и перехода на активное питание (Т4 до 67%) сопровождались торможением развития и морфологическими нарушениями, а после начала питания увеличением смертности и снижением начального роста. Эксперименты, в ходе которых производилось воздействие гормонами в хроническом и краткосрочном режиме, подтвердили выявленную в ходе наблюдений связь морфофизиологических и гормональных параметров развития, а также позволили оценить степень пластичности функциональных показателей в зависимости от содержания тиреоидных гормонов и кортизола. Удалось показать, что одним из самых явно выраженных фенотипических последствий увеличения содержания в организме тиреоидных гормонов в конце периода желточного питания является увеличение роста массы тела. При этом гормональное воздействие у молоди вызвало перераспределение гормонов в сторону Т3 и снижение в более чем в 10 раз фоновых значений кортизола. Направленность изменений после обработки гормонами соответствует эндокринному статусу ювенильных рыб с высоким темпом роста (Gomez, et al., 1997; McCormick, 1999) [3-6, 8, 15-17, 23, 24].
5. Проявлением развивающихся адаптивных изменений в результате воздействия тиреоидных гормонов во второй половине предличиночного развития явилось перераспределение лейкоцитарного состава (увеличение относительной доли лимфоцитов) на фоне снижения патологических изменений в целом организме и на клеточном уровне. Одной из наиболее вероятных причин, вызывавших эти изменения, может являться ускорение созревания клеток белой крови, отвечающих за противодействие процессам воспаления и аллергизации организма, что тестировано нами по увеличению доли зрелых форм гранулоцитов. Тиреоидные гормоны и кортизол оказывали одинаковый эффект на личинок в отношении клеточной устойчивости организма, но их выживаемость при переходе их к активному питанию зависела только от тиреоидных гормонов. Одной из наиболее вероятных причин снижения смертности у осетра на начальном этапе питания после гормональной обработки является более успешная адаптация к пище, поскольку тиреоидные гормоны у личинок рыб стимулируют активность пищеварительных ферментов (Кim, Brown, 1997, 2002), а их активность у осетра резко увеличивается к началу питания (Плотников, Проскуряков, 1984). Гипертрофия энтероцитов пилорического отдела, наблюдаемая у молоди осетра после кратковременной стимуляции тиреоидными гормонами, является косвенным подтверждением того, что последние стимулируют созревание пищеварительной функции [4-6, 8, 15, 16, 23, 24].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 |
