
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК 630*181.343+ 630*182.8 +631.427
Биомасса и активность почвенных микроорганизмов под разными видами и климатипами кедровых сосен
*, *, **
*Институт почвоведения и агрохимии СО РАН
**Институт леса СО РАН им.
Аннотация
В длительном полевом опыте на юге Красноярского края изучено влияние меж - и внутривидовой изменчивости кедровых сосен на некоторые микробиологические свойства серой почвы (содержание C и N биомассы микроорганизмов и их вклад в С и N органического вещества почвы, базальное и субстрат-индуцированное дыхание, метаболический и дыхательный коэффициент). Под сосной сибирской (Pinus sibirica Du Tour) по сравнению с сосной корейской (Pinus koraeansis Sieboldd et Zucc.) в валовой почве выше оказалась скорость субстрат-индуцированного дыхания почвы (13,4 vs. 11,5 мкл • ч-1 • г-1 почвы), а в почве ризосферы выше были содержание С биомассы (590 vs. 413 мкг С • г-1 почвы) и его вклад в органическое вещество почвы (1,1 vs. 0.9 %). Внутривидовая изменчивость изученных сосен проявлялась по-разному: у сосны сибирской (3 климатипа) по всей совокупности микробиологических свойств в многомерном плане и по содержанию N биомассы микроорганизмов в одномерном плане (11-15 мкг • г-1 почвы), а у сосны корейской (2 климатипа) – только в одномерном плане по базальному дыханию (1,9 vs. 2,3 мкл • ч-1 • г-1 почвы) и его отношению к субстрат-индуцированному дыханию (0,17 vs. 0,19).
Выявленное четкое различие изученных видов сосен по показателям роста и развития деревьев сильнее всего связано с соотношением C/N в биомассе почвенных микроорганизмов и с субстрат-индуцированным дыханием почвы, а внутривидовая гетерогенность роста и развития сосен положительно коррелирует с базальным дыханием почвы и отношением активных к потенциально активным микроорганизмам, отрицательно коррелирует с содержанием микробиального азота и его вкладом в общий фонд азота почвы.
В целом варьирование изученных микробиологических показателей, обусловленное непосредственным влиянием корней сосен на почву, превышает варьирование, обусловленное меж - и внутривидовой изменчивостью их физиолого-биохимических и продукционных особенностей. Поэтому изучению структуры и скорости развития корневых систем растений древесных видов следует уделять особое внимание как при почвенно-экологических исследованиях, так и при планировании технических мероприятий с ними.
Ключевые слова: сосна сибирская, сосна корейская, климатип, серая почва, ризосфера, биомасса почвенных микроорганизмов, дыхательная активность почвы, длительный полевой опыт, Западные Саяны
_________________________________________________________________
Введение
Растущая популярность применения деревьев хвойных видов для рекультивации нарушенных территорий [[1]], формирования защитных лесополос [[2]], озеленения и создания лесных культур [[3], [4]], а также изменение видового состава древесных растений в лесах под влиянием глобального потепления [ [5], [6], [7], [8] ], – все это вместе значительно повысило как важность правильного выбора видов деревьев в практическом плане, так и актуальность изучения взаимодействий в системе «почва ↔ растение» в плане более детального понимания влияния разных видов растений на свойства почв.
Растительный материал, поступающий в/на почву, является основным источником углерода и энергии для большей части почвенных микроорганизмов. Количество и качество растительного материала, поступающего с наземным и корневым опадом и корневыми выделениями, зависит от меж - и внутривидовой гетерогенности продукционных, физиологических и биохимических особенностей растений основных лесообразующих видов [ [9], [10]] и их изменения под воздействием внешних факторов, например, патогенов, рекреационной нагрузки и т. п.
У древесных растений корневые системы очень развиты, поэтому проникают на большую глубину и охватывают большой объем почвы. Соответственно, ризосфера является очень важным интерфейсом взаимодействия растения и почвы, а корневые выделения являются основным источником доступных углеродных субстратов для почвенных микроорганизмов [[11]]. Изменение химического состава и количества корневых выделений может значительно влиять на сообщество почвенных микроорганизмов [[12], [13]].
Разграничить влияние почвы на растения и, наоборот, растения на почву практически довольно сложно. Однако на территории нашей страны есть длительные опыты по выращиванию в одинаковых почвенно-климатических условиях семенных потомств деревьев разных видов и/или насаждений разного географического происхождения [[14], [15]], которые являются уникальными экспериментальными объектами, предоставляя, в частности, возможность изучения влияния растений на свойства почвы и ее микробного сообщества.
Целью этой работы было выявление влияния генотипической изменчивости кедровых сосен на биомассу и активность микробного сообщества почвы, в частности, почвы ризосферы, в условиях длительного (30 лет) полевого опыта.
Объекты и методы исследования
Опыт с географическими культурами (климатипами) кедровых сосен был заложен на серой почве в 1983 г. в Ермаковском лесхозе Красноярского края. Лесоводственные особенности опыта описаны ранее [[16]]. Сеянцы кедровых сосен – сибирской и корейской – были высажены из расчета 10 тыс. растений на 1 га с размещением 1,5 х 0,7 м. Географические культуры представлены тремя климатипами кедра сибирского: таштагольским Кемеровской области, шегарским Томской области и местным ермаковским Красноярского края, а также двумя климатипами кедра корейского: облученским Хабаровского края и чугуевским Приморского края. Перед закладкой опытов почву распахивали, т. е. почва опытных участков является антропогенно-трансформированной серой почвой. Опыт заложен в трехкратной повторности, и потомство каждого климатипа занимает по одной делянке на 3 соседствующих участках.
Отбор образцов почвы проводили в конце августа 2013 г. из слоя 0-20 см (сразу под подстилкой, с максимальной густотой корней) на расстоянии 60 см от ряда, что получалось в пределах подкроновой зоны [[17]]. Один смешанный образец валовой почвы составляли из 6 индивидуальных почвенных монолитов, отобранных случайным образом по делянке климатипа. Ризосферную почву отбирали после осторожного встряхивания корней 1-3 мм толщиной и сбора оставшейся на корнях почвы [[18]].
Почвенно-химические характеристики описаны ранее [[19]].
Содержание азота и углерода микробной биомассы определяли методом фумигации-экстракции [[20], [21]]. Влажную почву фумигировали очищенным от этанола хлороформом СНСl3 в течение 24 ч. при 25оС. Навески контрольной и фумигированной почвы экстрагировали 0,5 М раствором К2SO4 в соотношении почва : экстрагирующий раствор 1:4, суспензии фильтровали через очищенные от аммиака фильтры. Полученные экстракты анализировали на содержание общего азота и углерода по Кьельдалю и бихроматным сжиганием, соответственно. Содержание углерода микробиальной биомассы (Смик) рассчитывали как разницу концентрации Сорг в экстрактах фумигированной и контрольной почвы, деленную на 0,35 [21], а содержание азота микробиальной биомассы (Nмик) рассчитывали как соответствующую разницу в концентрации Nобщ, деленную на 0,54 [20].
Базальное дыхание (CO2) определяли путем измерения выделения СО2 почвой после стабилизации и без каких-либо добавок [[22]]. Субстрат-индуцированное дыхание (CO2-Глю) определяли путем измерения выделения СО2 после добавления глюкозо-минеральной смеси (0,8 мг С на 1 г сухой почвы), чтобы стимулировать преимущественно бактериальный рост [[23]]. По соотношению базального и субстрат-индуцированного дыхания рассчитывали дыхательный коэффициент (QR) [[24]], а по соотношению базального дыхания и биомассы микроорганизмов оценивали метаболический коэффициент (Qмет) [[25]].
Статистическую обработку данных, представленных в виде матрицы логарифмически трансформированных значений измеренных микробиологических свойств почв, проводили методом анализа главных компонент и дисперсионного анализа с помощью пакета Statistica v.6.1.
Результаты и обсуждение
Влияние межвидовой изменчивости сосен
Полученные оценки микробиологических свойств почвы (табл. 1) в целом совпадают с имеющимися в литературе данными для верхних (без подстилки) горизонтов почв лесных экосистем [[26], [27], [28], [29]], но оценок для ризосферы деревьев хвойных видов мало. Дисперсионный анализ не выявил статистически и экологически значимого влияния межвидовой изменчивости кедровых сосен на почти все изученные микробиологические свойства (вклад в общую дисперсию меньше 5%), за исключением глюкозо-индуцированного дыхания (табл. 2). Этот показатель был выше в почве под сосной сибирской (табл. 1), что отражает более выраженную физиологическую реакцию микроорганизмов, растущих и/или способных сразу или через несколько часов начать рост [[30]] (т. е. активных и потенциально активных микроорганизмов [24]), на поступление доступных углеродных субстратов.
Влияние почвы (валовой или ризосферной) на изменчивость изученных микробиологических свойств оказалось более выраженным (табл. 2): оно было выявлено по половине из изученных показателей и обусловливало значительную часть их общей дисперсии. Так, содержание Nмик и его отношение к Nорг, уровень базального дыхания и коэффициент QR в ризосфере сосен обоих видов были в 2 раза повышенными по сравнению с валовой почвой (табл. 1).
Таблица 1. Некоторые микробиологические свойства почвы под разными видами сосны (среднее ± ошибка среднего)
Показатель | Сосна | |||
сибирская | корейская | |||
Валовая почва | Ризосферная почва | Валовая почва | Ризосферная почва | |
Смик, мкг С • г-1 почвы | 445 ± 84 | 590 ± 89 | 561 ± 70 | 413 ± 76 |
Nмик, мкг • г-1 почвы | 13 ± 1,5 | 29 ± 8 | 11 ± 1,3 | 23 ± 7 |
Смик/Nмик | 44 ± 8,5 | 30 ± 11 | 66 ± 9,2 | 22 ± 2 |
CO2, мкл • ч-1 • г-1 почвы | 2,4 ± 0,2 | 4,5 ± 0,1 | 2,1 ± 0,2 | 4,2 ± 1,7 |
CO2-Глю, мкл • ч-1 • г-1 почвы | 13,4 ± 0,4 | 13,9 ± 1,4 | 11,5 ± 0,7 | 10,3 ± 0,9 |
QR | 0,18 ± 0,01 | 0,33 ± 0,03 | 0,18 ± 0,02 | 0,40 ± 0,13 |
Qмет, мкг С-СО2 × мг Смик-1 • ч-1 | 3,7 ± 0,8 | 3,8 ± 0,1 | 2,0 ± 0,3 | 4,8 ± 1,2 |
Cмик/Cорг, % | 1,4 ± 0,3 | 1,1± 0,1 | 1,7 ± 0,1 | 0,9 ± 0,0 |
Nмик/Nорг, % | 1,1 ± 0,1 | 2,2 ± 0,6 | 1,0 ± 0,1 | 2,4 ± 0,5 |
Таблица 2. Результаты одномерного и многомерного дисперсионного анализа микробиологических свойств почвы под кедровыми соснами: доля фактора в общей дисперсии (%) и уровень значимости p
Показатель | Фактор | ||
Вид сосны (A) | Почва (B) | Взаимодействие А ´ B | |
Смик | 2 (0,78) | 1 (0,99) | 10 (0,19) |
Nмик | 4 (0,28) | 47 (0,00) | 1 (0,62) |
Смик/Nмик | 2 (0,59) | 24 (0,03) | 6 (0,23) |
CO2 | 2 (0,47) | 59 (0,00) | 1 (0,96) |
CO2-Глю | 38 (0,01) | 1 (0,68) | 3 (0,37) |
QR | 2 (0,32) | 66 (0,00) | 2 (0,39) |
Qмет | 1 (0,72) | 11 (0,14) | 9 (0,18) |
Cмик/Cорг | 1 (0,81) | 14 (0,11) | 3 (0,42) |
Nмик/Nорг | 1 (0,87) | 53 (0,00) | 2 (0,52) |
Все показатели | (0,14) | (0,03) | (0,07) |
Примечание:
– жирным шрифтом выделены значения с уровнем значимости P ≤ 0,05.
Все это свидетельствует об относительно большем содержании активных микроорганизмов в ризосфере [30] в связи с более комфортной для них обстановкой в плане доступности углеродных субстратов [24]. Значительно суженным в ризосфере оказалось атомное соотношение С/N биомассы, что связано с различным физиолого-биохимическим состоянием микробных сообществ в почве ризосферы по сравнению с валовой почвой, т. к активно растущие клетки обогащены азотными соединениями. Логично предположить, что межвидовые различия сосен будут более выражены в ризосфере. И, действительно, дисперсионный анализ данных только образцов ризосферной почвы выявил влияние видов на Смик (P = 0,056) и Cмик/Cорг (P = 0,072): эти показатели оказались выше под сосной сибирской в 1,4 и 1, 2 раза, соответственно (табл. 1), что свидетельствует о повышенной доступности органических субстратов в ризосфере этого вида [[31]] и о большем вкладе биомассы микроорганизмов в формирование почвенного органического вещества [[32]].
Влияние внутривидовой изменчивости сосен
Полученные оценки микробиологических свойств валовой почвы под деревьями разных климатипов, представленные в таблице 3, в основном, не сильно варьировали под влиянием внутривидовой изменчивости сосен. Тем не менее, многомерный критерий значимости выявил достоверное влияние климатипа под сосной сибирской и отсутствие такового под сосной корейской (табл. 4, 5).
Таблица 3. Некоторые микробиологические свойства валовой почвы под разными климатипами двух видов кедровых сосен (среднее ± ошибка среднего)
Показатель | Сосна сибирская | Сосна корейская | |||
климатип | климатип | ||||
кемеровский | ермаковский$ | томский | хабаровский | приморский | |
Смик, мкг С • г-1 почвы | 551 ± 207 | 491 ± 120 | 322 ± 203 | 577 ± 127 | 544 ± 92 |
Nмик, мкг • г-1 почвы | 11 ± 3 | 12 ± 2 | 15 ± 2 | 10 ± 2 | 11 ± 1 |
Смик/Nмик | 61 ± 19 | 47 ± 10 | 24 ± 7 | 70 ± 13 | 62 ± 15 |
CO2, мкл • ч-1 • г-1 почвы | 2,2 ± 0,4 | 2,9 ± 0,5 | 2,0 ± 0,1 | 2,3 ± 0,2 | 1,9 ± 0,3 |
CO2-Глю, мкл • ч-1 • г-1 почвы | 13 ± 0,8 | 13 ± 0,8 | 14 ± 0,7 | 12 ± 0,8 | 11 ± 1,1 |
QR | 0,17 ± 0,03 | 0,21 ± 0,04 | 0,16 ± 0,01 | 0,19 ± 0,03 | 0,17 ± 0,02 |
Qмет, мкг С-СО2 • мг Смик-1 • ч-1 | 2,7 ± 0,9 | 4,0 ± 1,9 | 4,3 ± 1,8 | 2,2 ± 0,6 | 1,7 ± 0,2 |
Cмик/Cорг, % | 1,7 ± 0,7 | 1,5 ± 0,5 | 0,9 ± 0,3 | 1,7 ± 0,2 | 1,8 ± 0,2 |
Nмик/Nорг, % | 1,1 ± 0,4 | 1,0 ± 0,3 | 1,3 ± 0,2 | 0,9 ± 0,2 | 1,0 ± 0,1 |
$ Климатип из района проведения полевого опыта
Таблица 4. Результаты одномерного и многомерного дисперсионного анализа микробиологических свойств почвы под сосной сибирской: доля фактора в общей дисперсии (%) и уровень значимости p
Показатель | Фактор | ||
Климатип (A) | Почва (B) | Взаимодействие (A ´ B) | |
Смик | 3 (0,89) | 9 (0,45) | 5 (0,85) |
Nмик | 27 (0,03) | 51 (0,00) | 26 (0,04) |
Смик/Nмик | 0,43 | 0,39 | 0,41 |
CO2 | 4 (0,52) | 75 (0,00) | 1 (0,84) |
CO2-Глю | 36 (0,16) | 2 (0,63) | 45 (0,10) |
QR | 3 (0,51) | 73 (0,00) | 12 (0,11) |
Qмет | 1 (0,96) | 1 (0,95) | 3 (0,90) |
Cмик/Cорг | 4 (0,87) | 3 (0,66) | 4 (0,87) |
Nмик/Nорг | 30 (0,09) | 39 (0,02) | 23 (0,14) |
Все показатели | (0,04) | (0,08) | (0,04) |
Примечание:
– жирным шрифтом выделены значения с уровнем значимости P ≤ 0,05.
Таблица 5. Результаты одномерного и многомерного дисперсионного анализа микробиологических свойств почвы под сосной корейской: доля фактора в общей дисперсии (%) и уровень значимости p
Показатель | Фактор | ||
Климатип (A) | Почва (B) | Взаимодействие (A ´ B) | |
Смик | 7 (0,58) | 17 (0,40) | 3 (0,72) |
Nмик | 18 (0,08) | 63 (0,01) | 19 (0,07) |
Смик/Nмик | 1 (0,93) | 54 (0,09) | 1 (0,76) |
CO2 | 40 (0,00) | 49 (0,00) | 25 (0,02) |
CO2-Глю | 19 (0,34) | 9 (0,50) | 1 (0,99) |
QR | 27 (0,01) | 62 (0,00) | 21 (0,02) |
Qмет | 16 (0,09) | 70 (0,01) | 7 (0,22) |
Cмик/Cорг | 1 (0,91) | 72 (0,03) | 1 (0,94) |
Nмик/Nорг | 11 (0,09) | 76 (0,00) | 13 (0,07) |
Все показатели | (0,19) | (0,14) | (0,21) |
Примечание:
- жирным шрифтом выделены значения с уровнем значимости P ≤ 0,05.
Что касается отдельных показателей, то под сосной сибирской влияние климатипа обусловливало около трети общей дисперсии по содержанию Nмик и его отношению к Nорг, а также по субстрат-индуцированному дыханию (табл. 4). Под сосной корейской влияние климатипа проявилось по величине базального дыхания почвы, обусловливая почти половину варьирования этого показателя (табл. 5), и по дыхательному коэффициенту QR (табл. 4, 5), обусловливая почти треть его варьирования. Базальное дыхание почвы под хабаровским климатипом было в 1,6 раза выше, чем под приморским (табл. 3), то есть разница между климатипами по общей метаболической активности почвенных микроорганизмов может являться экологически очень значимой, обусловливая различную интенсивность минерализации органического вещества в почве под ними. По нескольким показателям – содержанию Nмик и Nмик /Nобщ, а также по Qмет – в почве под сосной корейской уровень значимости влияния внутривидовой изменчивости сосен находится в интервале 0,05 ≤ P ≤ 0,10 (табл. 5), что можно тоже счесть статистически значимым, если принять таковым уровень P ≤ 0,10, как это предлагается делать для экологических исследований [[33]]. Однако при анализе совокупности всех показателей влияние климатипа у сосны корейской не оказалось статистически значимым, что может быть связано с меньшим количеством изученных климатипов.
Таким образом, наиболее чувствительные к воздействию продукционных и физиолого-биохимических особенностей деревьев разных климатипов показатели – дыхательная активность почвы и абсолютное и относительное содержание микробиальной формы азота – связаны с минерализацией органического вещества и иммобилизацией азотных соединений. Отметим, что ранее в опытах с сосной обыкновенной мы выявили влияние климатипов по вкладу микробиального углерода в органическое вещество почвы и по соотношению C/N биомассы микроорганизмов [[34]], т. е. по показателям, связанным с эффективностью микробной утилизации и доступностью углеродных субстратов для микроорганизмов. Вероятнее всего, это расхождение связано с физиолого-биохимическими различиями сосен, а также с тем, что влияние климатипов сосны обыкновенной было выявлено при анализе данных по четырем полевым опытам с различными почвенно-экологическими условиями.
Рост сосен и микробиологические свойства почв
Анализ матрицы данных с показателями роста и развития деревьев разных климатипов обоих видов сосен (высотой и ее годовым приростом, диаметром ствола и кроны, числом веток в мутовке) в качестве переменных анализа и изученными почвенно-микробиологическими свойствами в качестве вспомогательных переменных позволил нагляднее представить структуру взаимосвязей между особенностями роста и развития деревьев разных видов и климатипов и микробиологическими свойствами почвы (рис. 1). Четкое межвидовое разграничение сосен по показателям роста и развития деревьев сильнее всего связано с соотношением C/N в биомассе почвенных микроорганизмов и с субстрат-индуцированным дыханием почвы. Так как сосна корейская, несмотря на новую для нее почвенно-климатическую обстановку, по показателям роста и развития деревьев (высота и скорость ее прироста, диаметр ствола и кроны) обгоняет сосну сибирскую [[35]], она выносит и больше азота, в результате чего последний относительно менее доступен для потребления микроорганизмами, и происходит расширение соотношения C/N в их биомассе и торможение метаболической активности [[36]]. Фитомасса изученных сосен различается по своему химическому составу, что тоже может влиять на соотношение C/N биомассы микроорганизмов почвы под разными видами. Что касается субстрат-индуцированного дыхания почвы, то его отрицательная корреляция с показателями роста и развития сосен (рис. 1), а, следовательно, и с поступлением растительного опада и корневых выделений, свидетельствует о меньшей доступности углеродных субстратов для микроорганизмов под сосной сибирской, объясняя причины более выраженной реакции микробов в почве под сосной сибирской на внесение легкодоступных субстратов (табл. 1, 2).
Внутривидовая гетерогенность ростовых показателей обоих видов сосен связана с базальным дыханием почвы и респираторным коэффициентом, т. е. соотношением активных и потенциально активных микроорганизмов: чем выше деревья и интенсивнее прирост, тем больше метаболическая активность микробного сообщества в почве под ними (рис. 1). И, наоборот, чем ниже и в целом менее развиты деревья, тем ниже метаболическая активность почвенных микроорганизмов, но из-за меньшего выноса деревьями – больше иммобилизованного микробной биомассой азота, и больше его вклад в общий азотный фонд почвы.
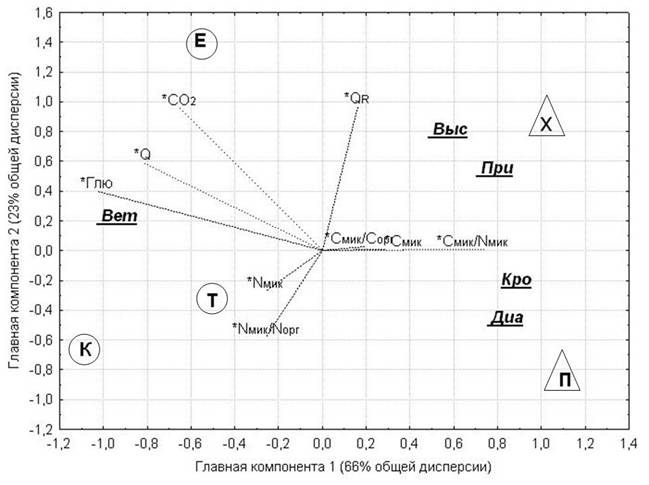
Рис. 1. Проекция микробиологических свойств и климатипов на плоскость первых двух главных компонент, экстрагированных из матрицы свойств древостоев. Обозначения переменных анализа: Выс – высота деревьев, При – годовой прирост высоты, Диа – диаметр ствола, Кро – диаметр кроны, Вет – число веток в мутовке. Обозначения вспомогательных переменных (*) – как в таблице 1. Обозначения климатипов сосны сибирской: Е – ермаковский, К – кемеровский, Т – томский. Обозначения климатипов сосны корейской: Х – хабаровский, П – приморский |
Выводы
За 30 лет развития искусственных фитоценозов влияние роста и развития доминантных видов растений, в данном случае кедровых сосен, было выявлено на некоторых почвенно-микробиологических свойствах. Большие высота и годичный прирост, а также диаметр ствола и кроны деревьев сосны корейской, очевидно, приводили и к большему поступлению на/в почву их растительного материала и, как следствие, более комфортной обстановке для микроорганизмов и их менее выраженной реакции на добавление легкодоступного субстрата к валовой почве. В почве ризосферы межвидовая изменчивость сосен оказала влияние на содержание микробиального углерода и его вклад в органическое вещество почвы. Внутривидовая изменчивость у сосны сибирской проявлялась по содержанию микробиального азота и его вкладу в общий азот почвы, а у сосны корейской климатип оказывал влияние на базальное дыхание почвы и респираторный коэффициент микроорганизмов.
Выявленное четкое различие изученных видов сосен по показателям роста и развития деревьев сильнее всего связано с соотношением C/N в биомассе почвенных микроорганизмов и с субстрат-индуцированным дыханием почвы, а внутривидовая гетерогенность у обоих видов сосен связана с базальным дыханием почвы, содержанием микробиального азота и его вкладом в общий азот почвы.
Заключение
В целом на изученной стадии развития фитоценоза варьирование микробиологических свойств, связанное с непосредственным влиянием корней сосен на почву, существенно превышает варьирование, обусловленное меж - и внутривидовой изменчивостью физиолого-биохимических и продукционных особенностей изученных кедровых сосен. Поэтому изучению структуры и скорости развития корневых систем растений древесных пород следует уделять особое внимание как при почвенно-экологических исследованиях, так и при планировании технических мероприятий с древесными видами.
Исследования выполнены при финансовой поддержке Российского Фонда Фундаментальных Исследований (проект № 13-04-01671) и проекта VI.54.1.4. «Биологические и биогеохимические функции почв как компонента наземных экосистем Сибири» (№ государственной регистрации 01201350243).
Список использованных источников
[1]. Zinn Y. L., Guerra Adriano R., Silva C. A., Faria J. A., Silva T. A. C. Soil organic carbon and morphology as affected by pine plantation establishment in Minas Gerais, Brazil // Forest Ecol. Manage. 2014. N. 318. – P. 261–269.
[2]. , , Сизов свойства черноземов под хвойными лесополосами // Вестник Алтайского государственного аграрного университета. – 2013, № 9 (107). – С. 23- 27.
[3]. , , Буторова показателей роста и генеративного развития кедровых сосен на плантации зеленой зоны города Красноярска // Сибирский лесной журнал. – 2014, т. 2. – С. 81-86.
[4]. Смирнов и натурализация хвойных на Сахалине // Известия высших учебных заведений. Лесной журнал. – 2013, № 5 (335). – С. 213-217.
[5]. , , Назимова и горные леса Южной Сибири. Красноярск. – 2006. – 170 c.
[6]. Penuelas J., Boada M. A global change-induced biome shift in the Montseny mountains (NE Spain) // Global Change Biol. 2003. V.9. N. 2. – P. 131-140.
[7]. Rehfeldt G. E., Tchebakova N. M., Milyutin L. I., Parfenova Y. I., Wykoff W. R., Kouzmina N. A. Assessing population responses to climate in Pinus sylvestris and Larix spp. of Eurasia with Climate-Transfer Models // Eurasian Journ. Forest Res. 2003. V. 6, N. 2. – P. 83-98.
[8]. Shao G., Yan X., Bugmann H. Sensitivities of species compositions of the mixed forest in eastern Eurasian continent to climate change // Global and Planetary Change 2003. V.37.N. 3-4. – P. 307-313.
[9]. Hobbie S., Oleksyn J., Eissenstat D., Reich P. Fine root decomposition rates do not mirror those of leaf litter among temperate tree species // Oecologia. 2010. V.162. P. 505–513.
[10]. Hansson K., Jlsson B. A., Olsson M., Johansson U., Kleja D. B. Differences in soil properties iny and Management. 2011. V. adjacent stands of Scots pine, Norway spruce and silver birch in SW Sweden // Forest Ecology and Management. 2011. V. 262. P. 522-530.
[11]. Kruse J., Simon J., Rennenberg H. Soil Respiration and Soil Organic Matter Decomposition in Response to Climate Change // Climate Change, Air Pollution and Global Challenges — Understanding and Perspectives from Forest Research. Developments in Environmental Sciences. 2013. V. 13. P. 131-149.
[12]. Falcini L., Naumova N. B., Kuikman P., Bloem J., Nannipieri P. Mineralization of different added organic substrates and its effect on native SOM mineralization and the structure of soil bacterial community // Soil Biology and Biochemistry.- 2003.- V.- 35., No.6.- P.775-782.
[13]. Michalet S., Rohr J., Warshan D., Bardon C., Roggy J.-C., Domenach A.-M., Czarnes S., Pommier Th., Combourieu B., Guillaumaud N., Bellvert F., Comte G., Poly F. Phytochemical analysis of mature tree root exudates in situ and their role in shaping soil microbial communities in relation to tree N-acquisition strategy // Plant Physiol. Biochem. 2013. V. 72. P. 169-177.
[14]. Ирошников культуры и плантации хвойных в Сибири. Новосибирск. – 1977. – С. 104-110.
[15]. Ирошников и перспективы интродукции сосны кедровой корейской / , // Леса и лесообразовательный процесс на Дальнем Востоке. Матер. Междунар. конфер., посв. 90-летию Колесникова . – 1999. – С. 101-102.
[16]. , Кузнецова Pinus sylvestris L. и Pinus sibirica Du Tour в географических культурах и лесных питомниках Красноярского края и Хакассии // Хвойные бореальной зоны. – 2012, т. XXVII (3-4). – С. 55-60.
[17]. Weber P., Bardgett R. D. Influence of single trees on spatial and temporal patterns of belowground properties in native pine forest // Soil Biology Biochemistry. 2011. V.43. P.1372-1378.
[18]. Zhao Q., Zeng D.-H., Fan Z.-P. Nitrogen and phosphorus transformations in the rhizospheres of three tree species in a nutrient-poor sandy soil // Applied Soil Ecology. 2010. V.46. P.341–346.
[19]. , , Кузнецова - и межвидовая изменчивость кедровых сосен: взаимосвязь с химическими свойствами почвы в длительном полевом опыте в Западных Саянах // АгроЭкоИнфо. 2014. № 2. http://agroecoinfo. narod. ru/journal/STATYI/2014/2/st_09.doc.
[20]. Brookes O. C., Landman A., Pruden G., Jenkinson D. S. Chloroform fumigation and the release of soil-nitrogen—a rapid direct extraction method to measure microbial biomass nitrogen in soil Soil Biol. Biochem, 1985, 17: 8, 837-842.
[21]. Vance E. D., Brookes P. C., Jenkinson D. S. An extraction method for measuring soil microbial biomass-C // Soil Biol. Biochem. `987.V.19. P.703-707.
[22]. Pell M., Stenstrom L., Granhall U. Soil respiration // Microbiological methods for Assessing Soil Quality. CABI International, 2006. P. 117-126.
[23]. Reischke S., Rousk J., Bååth E. The effects of glucose loading rates on bacterial and fungal growth in soil // Soil Biology & Biochemistry. 2014. V.70. P. 88-95.
[24]. Blagodatskaya E., Kuzyakov Y. Active microorganisms in soil: Critical review of estimation criteria and approaches // Soil Biol. Biochem. 2013. V.67. P.192-211.
[25]. Insam, H., Haselwandter, K. Metabolic quotient of the soil microflora in relation to plant succession // Oecologia. 1989.V.79. P. 174–178.
[26]. Ranger A., Insam H. Microbial activities and biomass along an altitudinal profile in the Northern Tyrolean Limestone Alps in Austria // Phyton. 1996. V. 36, N. 4. P. 83 – 94.
[27]. Papa S., Pellegrino A., Fioretto A. The contribution of bacteria and fungi to soil biological activity in a Pinus pinea wood on Vesuvius mount // Proceedings of the international workshop MEDPINE 3: conservation, regeneration and restoration of Mediterranean pines and their ecosystems. Options Méditerranéennes : Série A. 2007. n. 75. P. 167-173.
[28] Raubuch M., Joergensen R. G. C and net N mineralisation in a coniferous forest soil: the contribution of the temporal variability of microbial biomass C and N // Soil Biol. Biochem. 2002. V. 34, No. 6. P. 841-849.
[29]. , , Чернова биомасса, ее активность и структура в почвах старовозрастных лесов европейской территории России // Почвоведение. – 2011, № 4. – С. 479-494.
[30]. Placella S. A., Brodie E. L., Firestone M. K. Rainfall-induced carbon dioxide pulses result from sequential resuscitation of phylogenetically clustered microbial groups // PNAS USA. 2012. V.109. P.10931-10936.
[31]. Anderson T. H., Domsch K. H.1986. Carbon link between microbial biomass and soil organic matter // Proc. of the Fourth Int. Symp. on Microbial Ecology. Ljubljana,1986. P. 467-471.
[32]. Miltner A., Bombach P. SOM genesis: microbial biomass as a significant source // Biogeochemistry. 2011. V. 111. P.41-55.
[33]. Beninger P. G., Boldina I., Katsanevakis S. Strengthening statistical usage in marine ecology // Journ. Exp. Marine Biol. Ecol. 2012. V.426-427. P. 97-108.
[34]. , , Милютин климатипов сосны обыкновенной на химические и микробиологические свойства почв // Сибирский экологический журнал. – 2009, вып. 2. – С. 287-292.
[35]. Кузнецова , состояние и развитие кедровых сосен в географических культурах на юге Красноярского края // Хвойные бореальной зоны. 2010. Т. XXVII. № 1-2. С. 102-107.
[36]. Blagodatskaya E., Littschwager J., Lauerer M., Kuzyakov Y. Plant traits regulating N capture define microbial competition in the rhizosphere // European Journ. Soil Biol. 2014. V. 61. P. 41-48.
=====================================================================
Цитирование:
, , Кузнецова и активность почвенных микроорганизмов под разными видами и климатипами кедровых сосен // АгроЭкоИнфо. 2014. № 3. http://agroecoinfo. narod. ru/journal/STATYI/2014/3/st_12.doc.
