
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Посев селекционного материала проводился во второй декаде мая ручными сажалками и селекционной сеялкой Hege 95 с нормой высева 75 тыс. зёрен на 1 га. Посев линий в питомнике холодостойкости проведен в сроки посева ранних зерновых культур (третья декада апреля).
Гибридизация в питомниках холодостойкости, топкроссных и диаллельных скрещиваний произведена методом ручного опыления.
Определение холодостойкости проводилось полевым методом (Корнилов, 1957; Соколов, Ивахненко, 1963). Посев линий был проведен при температуре почвы на глубине 6-8 см в пределах 7-8 оС.
Полевые наблюдения и учёты, лабораторный анализ, учет урожая зерна и зелёной массы проводили по методике, утверждённой экспертно-методической комиссией Самарского НИИСХ. Полученный экспериментальный материал обработан методами корреляционного, регрессионного, одно - и двухфакторного дисперсионного анализа (, 1985).
Комбинационная способность оценивалась в топкроссных и диаллельных скрещиваниях по и др. (1980).
Многофакторный нелинейный регрессионный анализ произведен с помощью компьютерной программы «Аппроксимация экспериментальных данных с автоматическим подбором оптимального типа функции», разработанных в Оренбургском НИИСХ под руководством доктора с.-х. наук .
При определении доли генотип-средовых реакций в формировании количественных признаков качестве инбредного объекта исследований были взяты сорта яровой мягкой пшеницы, в качестве аутбредного объекта – гибриды кукурузы, которые испытывались в восьми экологических средах. По признакам, различия между вариантами по которым были достоверны, проводился двухфакторный дисперсионный анализ (фактор А – сорта, гибриды, фактор В - фоны).
В качестве меры вклада отдельных эффектов в фенотипическую изменчивость мы применяли показатель χ (χ2g=![]() ; χ2e=
; χ2e=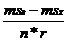 ; χ2g/e=
; χ2g/e=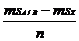 ; χ2z=msZ, где n - количество повторений, c - количество экологических точек, r - количество генотипов; Рокицкий, 1973).
; χ2z=msZ, где n - количество повторений, c - количество экологических точек, r - количество генотипов; Рокицкий, 1973).
Индекс реакции количественного признака на изменение среды определяли по формуле:
I=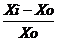 , где Xi и Xo – значение признака, соответственно при раннем и оптимальном сроке сева.
, где Xi и Xo – значение признака, соответственно при раннем и оптимальном сроке сева.
Подсчёты проводились на персональном компьютере с использованием пакета прикладных программ “Agros-2.09”.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Определение оптимальной продолжительности
вегетационного периода
Вегетационный период кукурузы определяется, прежде всего, ее генотипом, а варьирование по этому признаку в пределах рода составляет 70…75 дней у раннеспелых форм и 180…200 и больше у самых позднеспелых (Мелихов, 2004). Физиологически обусловленная положительная связь биологического урожая с продолжительностью вегетационного периода, формализованная V. H.Blackman (1919), с учётом соотношения фотосинтеза и дыхания в конкретных условиях окружающей среды принимает форму криволинейной зависимости в виде синусоиды или параболы (Шевелуха, 1995).
Для определения оптимальной продолжительности вегетации мы использовали такие критерии как число дней от всходов до выхода нитей (вегетативная фаза вегетации) и число дней от всходов до полной спелости (вегетационный период).
В таблице 1 приведены результаты нелинейного регрессионного анализа связи продолжительности вегетационного периода и продолжительности вегетативной фазы с урожаем зерна. В них приведены уравнения регрессии, полученные для каждого года исследований.
Высокий коэффициент детерминации (R2), который варьировал от 86,21 до 97,95 %, указывает на высокую степень соответствия аппроксимированных функций эмпирическим числовым рядам. Решение этих уравнений дает нам некоторую параболическую кривую, вершина которой и является искомым оптимумом. Анализируя, полученные данные мы можем констатировать, что оптимальная продолжительность вегетационного периода за 2006-2008 гг. варьирует от 100,68 до 111,96 дней, а периода от всходов до выхода нитей − от 52,37 до 55,43 дней. Это соответствует параметрам, принятым для среднераннеспелых гибридов кукурузы. При этом уровень урожайности составляет 3,47 – 6,02 т/га.
Таблица 1. Связь продолжительности вегетации (x) с урожаем зерна гибридов кукурузы, ц/га (y)
Год | Уравнение регрессии | Координаты вершины регрессионной кривой | R2 | F | |
Х opt | ŷ | ||||
вегетативная фаза | |||||
2006 | у=1684912-149574х+5302,6х2-93,8х3+0,829х4-0,0029х5 | 53,9±1,53 | 50,22±5,77 | 97,95 | 34,41** |
2007 | у=5828511,0-431955,6х+11999,75х2-148,09х3+0,685х4 | 53,5±0,49 | 52,05±8,10 | 93,47 | 12,53** |
2008 | у=1/(-39,87+2,914х-0,0797х2) | 54,2±0,97 | 41,93±7,24 | 86,57 | 6,38** |
вегетационный период | |||||
2006 | у=6822128-244828,1х+3293,95х2-9,69х3+0,0441х4 | 110,7±1,26 | 50,72±5,77 | 95,86 | 18,47** |
2007 | у=1/(-517,42+18,90х-0,259х2+0,00157х3) | 110,1±0,87 | 50,39±8,10 | 86,21 | 5,93** |
2008 | у=670038,7-12040,7х-167,49х2 +5,550х3 -0,0461х4 | 101,5±0,88 | 43,66±7,24 | 89,79 | 8,04** |
** функция удовлетворяет критерию адекватности Фишера для 1% уровня значимости
Доля генотип-средовых реакций в формировании количественных признаков
Используя молекулярные маркёры на кукурузе (Blanc et al, 2006) и арабидопсисе (Juengei et al, 2005; Malberg et al, 2006) выявлено значительное количество QTL-QTL эпистатических взаимодействий по самым разным признакам. Причём величина эпистатических (и генотип-средовых) взаимодействий примерно в два раза превышает аддитивные эффекты. При этом следует заметить, ещё в конце 50-х – начале 60-х годов было выявлено на дрозофиле (Parsons, 1959) и домашней птице (Hull et al, 1963), что эффекты генотип-средовых взаимодействий у гетерозигот неизменно ниже, чем у гомозиготных родительских форм.
Мы поставили целью проверить эту закономерность на аутбредном (кукуруза) и инбредном (пшеница) растительных объектах.
Как видно из таблицы 2, по всем изучавшимся признакам доля генотипа в формировании фенотипа яровой пшеницы примерно одинакова и равна 10,0-15,5%. По двум признакам (количество зёрен в колосе и высота растения) доля эффектов взаимодействия «генотип/среда» значительно ниже, чем генотипические и, особенно, средовые вклады. По этим признакам отбор может быть эффективным в локальных условиях при наличии контрастных по фенотипу рекомбинантов в гибридных популяциях.
Три признака (урожай зерна, масса зерна с колоса и длина верхнего междоузлия) имеют существенные нагрузки эффектов генотип-средового взаимодей-
Таблица 2. Эффекты (χ) и доля влияния (%) генотипа, среды и взаимодействия «генотип/среда» на формирование количественных признаков у яровой пшеницы
Источник варьирования | Урожай зерна, г/м2 | Количество зёрен в колосе | Масса зерна с колоса, г | Высота растения, см | Длина верхнего междоузлия, см | |||||
χ | % | χ | % | χ | % | χ | % | χ | % | |
Сорта (А) | 8,24 | 10,0 | 0,51 | 11,0 | 0,026 | 11,5 | 1,97 | 15,5 | 1,21 | 11,3 |
Условия среды (В) | 30,7 | 37,1 | 1,67 | 35,9 | 0,059 | 25,8 | 5,88 | 46,4 | 3,60 | 33,5 |
Взаимодей-ствие (А х В) | 24,8 | 30,0 | 0,15 | 3,3 | 0,050 | 21,7 | 0,74 | 5,8 | 3,42 | 31,8 |
Ошибка (z) | 18,7 | 22,9 | 2,31 | 49,8 | 0,094 | 41,0 | 4,09 | 32,3 | 2,52 | 23,4 |
ствия (в 2-3 раза превышающих вклад генотипа). Отбор по ним может быть эффективным лишь вдоль специального экологического вектора (Сюков и др., 2008).
В отличие от пшеницы в экспериментах с гибридной кукурузой по большинству признаков достоверных эффектов генотип-средовых взаимодействий отмечено не было. По признакам «урожай зерна» и «количество дней до выноса нитей» межсортовые различия вносили значительно больший вклад в формирование фенотипа, нежели все изучавшиеся количественные признаки у инбредного объекта (табл. 3).
Таблица 3. Эффекты (χ) и доля влияния (%) генотипа, среды и взаимодействия «генотип/среда» на формирование количественных признаков у кукурузы
Источник варьирования | Урожай зерна, г/м2 | Уборочная влажность зерна, % | Количество дней до выноса нитей | |||
χ | % | χ | % | χ | % | |
Сорта (А) | 0,2927 | 23,2 | 0,6437 | 8,4 | 0,8533 | 16,4 |
Условия среды (В) | 0,3320 | 26,3 | 5,0904 | 66,0 | 3,0680 | 58,5 |
Взаимодействие (А х В) | 0,2500 | 19,8 | 1,1420 | 14,8 | 0,5309 | 10,1 |
Ошибка (z) | 0,3887 | 30,7 | 0,8342 | 10,8 | 0,7884 | 15,0 |
Генотип-средовая составляющая у кукурузы по этим признакам была менее значима, чем межсортовая. По признаку «уборочная влажность зерна» соотношение генотипической и генотип-средовой составляющей было аналогичным таковому у большинства количественных признаков, наблюдаемых у инбредной культуры. Однако вклад взаимодействия «генотип-среда» всё же вдвое ниже, чем у пшеницы. То есть, в целом подтверждается отмеченная ранее тенденция на животных объектах о неизменно более низком уровне вклада генотип-средовых эффектов в формирование количественных признаков у гибридов по сравнению с гомозиготными формами. Учитывая в значительной степени эпигенетический характер взаимодействия «генотип-среда», эту закономерность можно объяснить с позиций гипотезы «эпигенетического баланса».

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 |
