

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
(канд. сель.-хоз. наук)
ЭКОЛОГИЗАЦИЯ ЗЕРНОПРОИЗВОДСТВА ПРИ АКТИВАЦИИ РАННИХ РОСТОВЫХ ПРОЦЕССОВ В СЕМЕНАХ ПОД ДЕЙСТВИЕМ РЕГУЛЯТОРОВ РОСТА
г. Пенза, РГУИТиП
Процесс экологизации неразрывно связан с последовательным внедрением идей сохранения природы и устойчивости окружающей среды в сферах экономики, управления, разработки технологий и многих других. В современном растениеводстве регуляторы роста рассматриваются как экологически чистый и экономически выгодный способ повышения урожайности сельскохозяйственных культур, позволяющий полнее реализовывать потенциальные возможности как отдельных растительных организмов, так и агроценозов.
Регуляторы роста представляют собой физиологически активные вещества биогенного происхождения или синтезированные искусственно. По механизму действия на растения регуляторы роста условно делятся на индукторы (включение под действием вещества процесса, который не шел в клетке в отсутствие данного элемента) и стимуляторы (усиление, активация уже идущих процессов). Однако в ряде случаев одновременное осуществление этих механизмов делает невозможным провести четкую грань между их проявлениями.
Можно охарактеризовать действие фитогормонов на растения как поливалентное: все они влияют на рост и деление клеток, на процессы адаптации и старения, на транспорт веществ, дыхание, синтез нуклеиновых кислот и белков и на многие другие процессы.
Влияние многих гормонов основано на проникновении внутрь клетки через плазматическую мембрану, а затем через цитозоль в ядро. Там молекула гормона образует комплекс с ядерными рецепторами и в результате изменяет матричные синтезы. Ответная реакция на регуляторное действие этого класса гормонов бывает медленной, поскольку она связана с синтезом новых молекул [, 1998; , 2002].
По данным ряда авторов [, 1995; , 2002] сигнальные системы клеток растений воспринимают, умножают и передают на геном первичный внутриклеточный сигнал, что приводит к экспрессии определенных генов и ответу клеток на первичный сигнал (гормон, элиситор). Поэтому действие фиторегуляторов можно рассматривать как сигналы для переключения программы физиологических процессов в организме.
В настоящее время большое внимание уделяется экологическим аспектам применения регуляторов роста растений, которые, как правило, являются малотоксичными соединениями с невыраженной видовой чувствительностью, слабо выраженными кумулятивными свойствами, широким спектром биологического действия. Физиологическое действие многих из них связано с изменением гормонального статуса растений. Выявлено влияние эндогенных регуляторов роста и некоторых их синтетических аналогов на митотическую активность клеток, изменение хромосом, на активность генетического аппарата и белоксинтезирующие системы, а следовательно, и на хозяйственные свойства растений.
В связи с особенностями свойств и проявлений биологической активности стимуляторов роста и фитогормонов в производство допускают препараты, прошедшие токсикологическую проверку и оценку генетического риска, регуляторы используют только на культурах, для которых они рекомендованы, при строгом соблюдении регламента и мер безопасности.
Современная наука создает регуляторы нового поколения, воздействующие на растения в минимальных дозах, всего несколько миллиграммов на 1 гектар, при стоимости гектарной нормы 100 рублей. Такие концентрации, несомненно, являются факторами защиты окружающей среды от загрязнений [, , 1984].
При большом разнообразии природных и синтетических регуляторов роста механизм действия многих из них до конца не изучен и требует проведения дальнейших исследований с целью повышения продуктивности сельскохозяйственных растений путем изменения ростовых показателей и активации физиологических процессов. Поэтому рассматриваемая тема является актуальной.
Для изучения влияния регуляторов роста на начальные этапы прорастания семян ярового ячменя использовались синтетические регуляторы роста – мелафен и пирафен, а также природный полисахарид – пектин.
Мелафен представляет собой меламиновую соль бис (оксиметил) фосфиновой кислоты. Данный препарат был синтезирован в Институте органической и физической химии им. Казанского научного центра РАН , и др.
Формула мелафена:
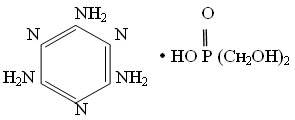
Синтез мелафена прост и технологичен, исходные вещества являются промышленно доступными. Препарат растворим в воде и его водные растворы стабильны при хранении годами. Мелафен обладает высокой эффективностью и широким спектром действия при чрезвычайно низких применяемых концентрациях. Препарат малотоксичен для теплокровных (LD50 2000 мг/кг на мышах и 6000 мг/кг на крысах) и не обладает ДНК – повреждающей и мутагенной активностью в широком диапазоне концентраций (Фаттахов и др., 2000).
Пирафен является аналогом мелафена. Молекулярный вес 251,18. Растворим в воде. Температура плавления 181 – 183 ºС.
Пектин – полисахарид, построенный преимущественно из D-галакту-ронопиранозильных остатков, соединенных 1 → 4-гликозидной связью, полимерные цепи которых этерифицированы этанолом. Для активизации физиолого-биохимических процессов и повышения продуктивности сельскохозяйственных культур используют пектиновые вещества с молекулярной массой 20000 у. е. из Amaranthus cruentus, оказывающие как активирующее, так и ингибирующее действия.
Для полноценного раскрытия механизма влияния регуляторов роста на растения необходимо рассматривать семя как сложную саморегулирующуюся живую систему, с помощью различного рода воздействий на которую можно направленно изменять метаболизм и связанные с ним физиологические функции – рост, развитие, продуктивность и прочие.
Процесс прорастания семян является критическим периодом в жизни растительного организма, во многом обеспечивающим выживаемость того или иного экотипа. Семена, находящиеся в состоянии вынужденного покоя, в период прорастания проходят три этапа: (1) активация метаболизма; (2) подготовка к началу роста растяжением; (3) собственно рост органов проростка.
На первом этапе – этапе гидратации – гидрофобные частицы спонтанно притягивают воду. Этот процесс сопровождается переходом в активное состояние ферментов, витаминов, регуляторов роста, что обеспечивает мобилизацию запасных питательных веществ путем их гидролиза и поступление растворимых веществ к точкам роста. Поэтому быстрота и степень набухания семян связаны с пробуждением зародыша к активной жизнедеятельности (, 1966; Антонова Т. А., 2004).
В результате проведенных исследований было установлено, что процесс водопоступления в семенах ячменя соответствует классическим представлениям и характеризуется кривой набухания, имеющей S-образный вид (рисунок 1).
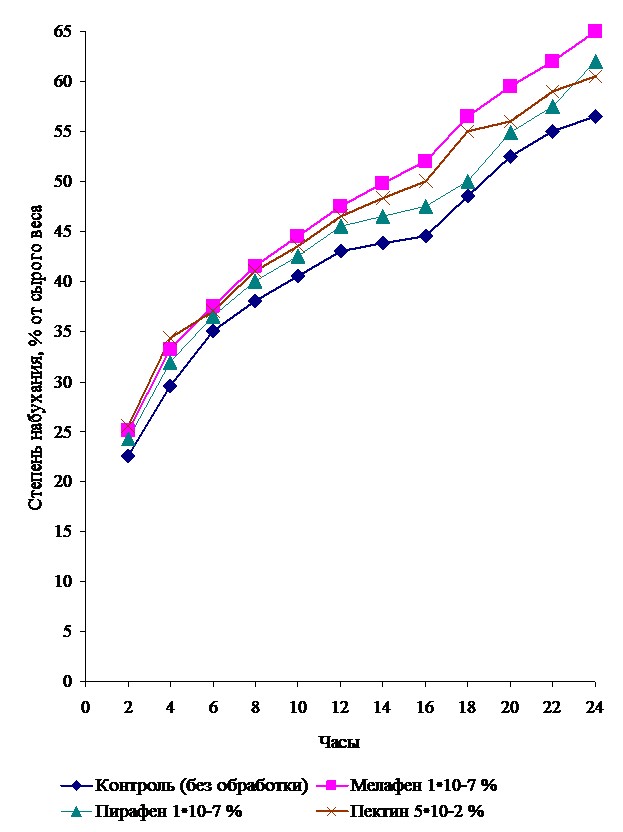
Рис. 1. Степень набухания семян ярового ячменя при прорастании,
% от сырого веса
Наиболее интенсивное набухание отмечалось в первые 6-8 часов намачивания семян. К этому моменту семена ячменя достигали уровня 38,0-41,5% влажности по всем изучаемым вариантам, включая контроль. Превышение контрольных данных в опытных образцах составляло 1,5-4,7%.
Примерно через 12 часов после намачивания (второй этап прорастания – подготовка к растяжению клеток) наблюдается лаг-период, в течение которого скорость поглощения кислорода и воды, а также уровень АТФ не меняются или даже падают. Лаг-период в этом случае может рассматриваться как время, в течение которого первичный стимул (вода) вызывает ответную метаболическую реакцию и активацию или синтез факторов прорастания (гормонов).
Следующая за лаг-периодом фаза характеризовалась повторным увеличением темпов водопоступления, однако степень набухания возрастала не так значительно, как на первых этапах поглощения воды семенем. Иными словами, в возрасте 36-48 часов семена ячменя достигают критической влажности, в связи с чем происходит рост растяжением самого гипокотиля, в результате чего кончик корешка выталкивается из семени.
При прорастании в семенах наблюдается раннее и быстрое формирование мембранных систем, совпадающее по времени с поглощением воды или следующее немедленно за ним [Майер A. M., 1982]. Изменение свойств мембран затрагивает не только активность, связанных с мембранами ферментов, но и свойства самих мембран, например их проницаемость. При обработке семян пектином химические процессы, возникающие в липидах мембран, приводят к изменению проницаемости и сорбционных свойств последних, что вызывает активизацию ферментных систем [ и др., 2006]. Фосфиновая группа молекулы мелафена, обладающая высокой активностью, действует подобно фосфатной группе АТФ при контакте с внешней мембраной растительной клетки и усиливает сигналы к осуществлению метаболических процессов [ и др., 2004]. Пирафен, являясь структурным аналогом мелафена с иным радикальным замещением, возможно, оказывает подобное действие.
Таким образом, в результате проведенных исследований было установлено, что процессы водопоступления в семенах ячменя описываются кривой набухания, имеющей трехфазный характер, что соответствует современным представлениям о набухании семян. Использование различных регуляторов роста не изменяет общей направленности процесса, но увеличивает скорость водопоступления, которая выражается в более раннем достижении пороговых уровней, необходимых для активизации метаболических процессов семени. Именно в результате метаболической регуляции происходит дальнейшее усилении всех активированных систем.
При распаде углеводов в организме идет высвобождение энергии, которая запасается в макроэргических связях АТФ, и АТФ, синтезированная сопряжено с окислением углеводов, поставляет энергию для осуществления всех химических процессов. Кроме того, углеводы являются источником большого числа органических соединений, которые служат исходными продуктами для биосинтеза липидов, белков и нуклеиновых кислот, необходимых для активного роста и развития зародыша семени [Карпова Г. А., 2009].
В семенах многих зерновых культур основной запасной формой углеводов является крахмал (ячмень – 70%) [, 1968]. Реакция гидролиза крахмала ускоряется амилазами – спецефическими ферментами, относящимися к подклассу гидролаз гликозидов (класс гидролаз). Одни из них, находящиеся в эндосперме или зародыше в связанном состоянии, активируются под влиянием набухания, а некоторые ферменты образуются вновь [, 1978]. Новообразование белков-ферментов становится возможным в результате образования в клетках семени и-РНК в первые часы после намачивания [, 2005].
Фермент -амилаза образуется de-novo в процессе прорастания, β-амилаза уже находится в сухих семенах в связанном состоянии. Ведущая роль в разложении гранул крахмала с образованием декстринов и сахаров принадлежит -амилазе (1,4-глюкан-4-глюкогидролаза), расщепляющей активные гранулы крахмала. Под действием β-амилаз (1,4-глюкан-мальтогидролаза) продукты -амилазного гидролиза превращаются в простые сахара. При усилении действия ферментов, прежде всего амилотического комплекса, с высокой активностью -амилазы происходят глубокие физиолого-биохимические изменения в прорастающем зерне [ и др., 2006]
В ходе проведенных исследований установлено увеличение активности - и β-амилаз при обработке семян мелафеном, пирафеном и пектином в первые 72 часа наблюдений (рисунок 2).
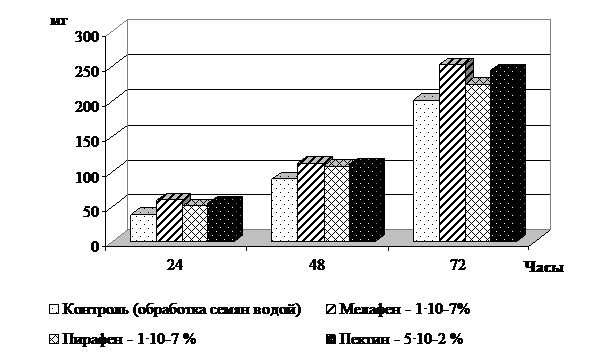
Рис. 2. Суммарная активность α- и β- амилаз в семенах ячменя при прорастании, мг гидролизованного крахмала за 1 час/1 г сухого вещества зерновки
Максимум активности ферментов зарегистрирован на 3 сутки с превышением контроля на 11,4-24,9%. Увеличение активности амилаз под действием регуляторов роста направлено на мобилизацию потенциальных возможностей организма, что способствует интенсивному росту растения и эффективному усвоению питательных веществ клетки [, 2004]. Эти процессы обусловлены интенсификацией дыхания, сопряженного с окислительно-восстановительными реакциями растения.
В организме растения окислительный режим регулирует фермент каталаза. Его интенсивность рассматривается в качестве интенсивности и продуктивности общего обмена. Также каталаза защищает живые организмы от повреждения перекисью водорода, образовавшейся в результате окислительно-восстановительных реакций.
Усиление активности каталазы в проведенных опытах было прямо пропорциональным степени набухания семян (рисунок 3).
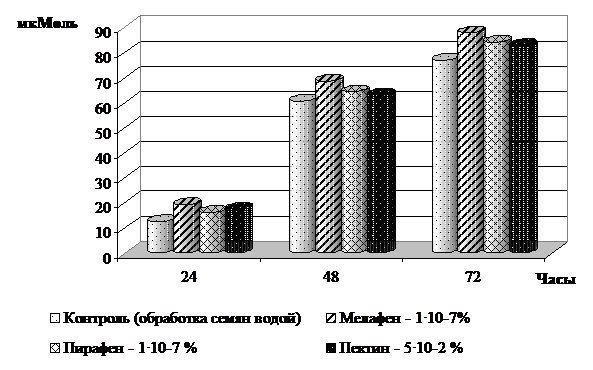
Рис. 3. Активность каталазы в семенах ячменя при прорастании, мкМоль разложившейся Н2О2 за 1 мин/ 1мл ферментного раствора
Наибольшая активность данного фермента наблюдалась на 3 сутки и превысила контрольные показатели на 7,6-14,1%. Максимальное увеличение активности каталазы под действием регуляторов роста относительно контроля было зарегистрировано в 24 часа и составило 30,1-54,7%. В 48 часов эффективность действия регуляторов роста снижалась до 4,1-13,0%, затем незначительно возрастала (7,6-14,1%).
Выявленная динамика активности каталазы, вероятно, объясняется повышенным востребованием энергии на интенсификацию физиолого-биохимических процессов в прорастающем семени. Эта энергия поступает преимущественно при окислении запасных питательных веществ (углеводов и липидов), сопровождающихся образованием большого количества перекисных соединений, утилизируемых каталазой.
Таким образом, природные и синтетические регуляторы роста и развития растений являются мощным и экологически безвредным средством управления онтогенезом растительных организмов, поэтому их применение становится все более перспективным и быстроразвивающимся направлением интенсификации производства.
Выявленная в процессе исследований способность изучаемых факторов активизировать физиолого-биохимические процессы в период прорастания семян представляет практический интерес, так как все искусственно создаваемые человеком экосистемы особенно нуждаются в поддержании на начальных стадиях сукцессии.
Список литературы.
1. Антонова урожая, зимостойкость и качество зерна озимой ржи при использовании мелафена в условиях лесостепи Поволжья. автореферат дисс. ... канд. сельсхоз. наук / . – 2004. – 19 с.
2. Виленский раскрывает свои тайны / , . – М.: Колос, 1984. – 110 с.
3. Карпова использования регуляторов роста и бактериальных препаратов на яровой пшенице / , // Зерновое хозяйство. – 2009. – № 5. – С. 16–17.
4. Кашина влияния фосфороорганического соединения мелафена на рост и энергетические процессы клеток хлореллы: автореферат канд. биол. наук / . – Казань, 2004. – 21 с.
5. Костин минерального питания и росторегуляторы в онтогенезе сельскохозяйственных культур / , , . – М.: Колос, 2006. – 290 с.
6. Кулаева регулируется жизнь растений / // Соровский образовательный журнал. – 1995. – № 1. – С. 20-27.
7. Лебедев растений / . – Киев: «Вища школа», 1978.
8. Майер регуляция прорастания / ; под ред. // Физиология и биохимия покоя и прорастания семян. – М.: Колос, 1982. – С. 99-132.
9. Прокофьев семян как органов запаса. XXVII Тимирязевское чтение / . – М.: Наука, 1968. – 52с.
10. Романов, фитогормонов / // Физиология растений. – 2002. – № 4.
11. Строна семеноведение полевых культур / . – М.: Колос, 1966. – 464 с.
12. Тарчевский системы клеток растений / . – М.: Мир, 2002. – 140 с.
13. Филиппов внешние сигналы передаются внутрь клетки / // Статьи Соросовского образовательного журнала. – М., 1998, серия «Биология». – С. 78-84.
14. Якушкина растений: учебник для студентов вузов, обучающихся по специальности 032400 «Биология» / , . – М.: Гуманитар. изд. центр ВЛАДОС, 2005. – 463 с.
