
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Материалы для самостоятельной работы студентов
(Составители – , , )
СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ
1. Механизмы электрогенеза миокардиальных клеток
Функциональным элементом сердца является мышечное волокно - цепочка из клеток миокарда, соединенных “конец в конец” и заключенных в общую саркоплазматическую оболочку.
Потенциал покоя (ПП) миокардиальных волокон формируется за счет высокой проницаемости поверхностной протоплазматической мембраны миокардиального волокна для катионов калия. Возникновение потенциалов действия (ПД) обусловлено открытием натриевых потенциалзависимых каналов поверхностной протоплазматической мембраны. Значительный вклад в генерацию ПД миокардиальных волокон вносят потенциалзависимые кальциевые каналы. Возбуждение распространяется по сердцу без декремента, механизм распространения - электрический.
Сердечная мышца неоднородна в своем клеточном составе. Различают типичные (сократительные) и атипичные волокна миокарда. Они различаются по строению, функции и электрической активности. 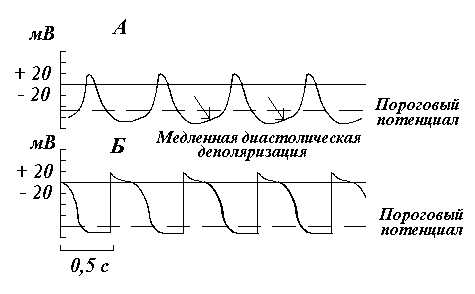
Рис. 1. Конфигурация потенциалов действия атипичной мышечной ткани (А) и типичного миокарда желудочков (Б).
Для типичных миокардиальных волокон (ТМВ), образующих основную массу сердечной мышцы и осуществляющих ее сократительную деятельность, характерна электрическая активность, представленная на рис. 1, Б. На электрограммах отчетливо прослеживается стабильный уровень ПП порядка -90 мВ. Возбуждение ТМВ проявляется потенциалом действия, восходящая ветвь которого отражает изменение потенциала внутри волокна относительно внеклеточной среды от - 90 до +10, +20 мВ. В отличие от поперечнополосатых мышечных волокон, миокардиальное волокно обладает небольшим потенциалом инверсии - не более +20 мВ. Однако наиболее характерные особенности присущи процессу реполяризацни сердечной мышцы. Если в волокнах скелетной мышцы она происходит в течение короткого времени - 3-5 мс, то в ТМВ она происходит в течение 250-350 мсек. На реполяризационной ветви ПД типичных миокардиальных волокон отчетливо выражены три части: начальная быстрая реполяризация, медленная реполяризация (плато), конечная быстрая реполяризация. Реполяризация завершается возвращением мембранного потенциала на устойчивый уровень ПП.
Иначе выглядят мембранные потенциалы, регистрируемые в атипичных волокнах миокарда (АТМВ). В них нет устойчивого уровня поляризации. Электрическая активность АТМВ представляет собой непрерывные колебания мембранного потенциала. По достижении им определенной величины (примерно - 60 мВ) спонтанно начинает развиваться медленная диастолическая деполяризация. На уровне около -40 мВ процесс деполяризации ускоряется, кривая идет круче ( рис. 1, А), что соответствует возникновению в АТМВ потенциала действия. Деполяризация сменяется реполяризацией, в конце которой вновь начинается медленная диастолическая деполяризация, лежащая в основе самопроизвольной ритмической активности сердца.
Свойство миокарда возбуждаться под влиянием ПД, спонтанно возникающих в нем самом, называется автоматизмом (автоматией). Электрические процессы, связанные с ним, происходят в АТМВ. Таким образом, атипичные миокардиальные волокна служат источником автоматизма сердечной мышцы.
2. Особенности распространения возбуждения по миокарду
Атипичная миокардиальная ткань сосредоточена в виде островков в различных отделах сердца, объединяющихся в проводящую систему сердца (рис. 2). Особенности проведения возбуждения по миокарду связаны со способом соединения между собой его отдельных волокон. Под электронным микроскопом между ними обнаружили особые вставочные диски. Они представляют собой дубликатуры мембран, в которых имеются щелевые контакты между соседними волокнами. Открытие вставочных дисков дало основание рассматривать миокард как функциональный синцитий, поскольку электрический импульс (ПД), возникший в одном волокне, распространяется на соседние посредством электрической передачи. Благодаря этому сердцу присущи свойства гомогенной возбудимой системы. Так, в реакциях на одиночные раздражения миокард подчиняется закону “все или ничего”. Электрическая связь существует не только между типичными мышечными волокнами сердца. Аналогично связаны атипичные миокардиальные волокна как между собой, так и с типичными мышечными волокнами. Поэтому возбуждение, ритмически возникающее в АТМВ, легко переходит на ТМВ и распространяется по всему сердцу.
Между скоплениями атипичной миокардиальной ткани установлена четкая субординация: ведущим является то скопление АТМВ, которое расположено в правом предсердии - между венозным синусом и ушком - синоатриальный узел. Когда он автоматически возбуждается, другие островки АТМВ выполняют только функцию проведения. Их собственный автоматизм угнетен. Поэтому синоатриальный узел называют пейсмекером - водителем ритма, так как ритм его автоматических возбуждений определяет частоту сердечных сокращений. Количественной мерой автоматизма различных АТМВ является частота спонтанных возбуждений. Чем она больше, тем выше уровень автоматизма. Основным водителем ритма служит группа из примерно 5000 атипичных волокон, занимающих около 3,5% объема этого узла. Отдельные волокна, входящие в группу, возбуждаются синхронно и синфазно, что обеспечивается электротоническим характером их взаимодействия. Передача возбуждения на другие группы АТМВ синоатриального узла, а также на ТМВ предсердий осуществляется посредством ПД. Благодаря тому, что водителем ритма является не одиночное АТМВ а группа волокон, сердце имеет высокую надежность в поддержании спонтанных колебаний.
С АТМВ синоатриального узла возбуждение переходит на ТМВ предсердий и проводится по ним со скоростью около 1 м/с. Уже через 40 мс от возникновения ПД в водителе ритма все участки предсердий находятся в возбужденном состоянии. Возбуждение распространяется по предсердиям широким фронтом, что обеспечивается наличием щелевых контактов не только между торцами отдельных миокардиальных волокон, имеющих продольное расположение, но и между их боковыми поверхностями. Однако плотность щелевых контактов в торцевых
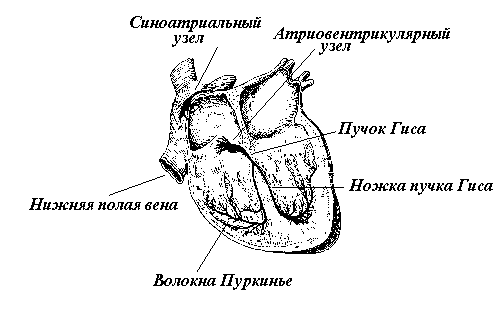
Рис. 2. Схема строения проводящей системы сердца.
вставочных дисках выше, чем в боковых, поэтому по направлению к желудочкам возбуждение движется быстрее, чем поперек предсердий. Тем самым достигается сокращение всего миокарда предсердий с одномоментным выходом волны возбуждения на атриовентрикулярный узел проводящей системы сердца. Как известно, предсердия отделены от желудочков фиброзной тканью, которая не способна проводить возбуждение. Вместе с тем, в этой преграде есть узкая щель - шириной чуть более 1 мм и длиной 1,5-2 мм, в которой расположен атриовентрикулярный узел, проводящий возбуждение из предсердий в желудочки. В местах контакта с типичным миокардом предсердий АТМВ атриовентрикулярного узла очень тонки, вследствие чего им присуще значительное электрическое сопротивление саркоплазмы. В этом одна из причин резкого в 20-50 раз замедления распространения возбуждения в атриовентрикулярном узле по сравнению с предсердиями. Другая причина заключается в том, что АТМВ в верхней части узла имеют не продольное, а поперечное расположение. Следовательно, по направлению к желудочкам возбуждение передается через боковые, а не более эффективные торцевые вставочные диски.
Замедленное проведение возбуждения из предсердий в желудочки обеспечивает важную для нормальной работы сердца паузу между сокращениями. Ее называют атриовентрикулярной задержкой. Желудочки начинают сокращаться примерно через 0,1 с от начала сокращения предсердий. Задержка нужна для того, чтобы кровь, накопленная предсердиями в диастолу, полностью перешла в желудочки до того, как они начнут сокращаться, нагнетая ее в аорту.
Из атриовентрикулярного узла возбуждение поступает в пучок Гиса. Там скорость проведения возбуждения возрастает до 2-3 м/c. Увеличение скорости обусловлено утолщением АТМВ и повышением плотности щелевых контактов во вставочных дисках. Ближе к верхушке сердца от пучка Гиса отходят волокна Пуркинье. Эти атипичные миокардиальные волокна вступают в контакт с ТМВ желудочков. Волокна Пуркинье обладают наибольшим диаметром по сравнению с другими волокнами миокарда. Поэтому скорость проведения возбуждения здесь достигает 4-5 м/с. Возбуждение с отдельных волокон Пуркинье переходит на огромное число ТМВ практически одномоментно, благодаря чему разные участки желудочков сокращаются синфазно.
3. Электромеханическое сопряжение в миокарде
Сокращение миокардиальных волокон, как и волокон скелетных мышц, инициируется потенциалом действия. Тем не менее временные соотношения между параметрами потенциала действия и параметрами сокращения этих мышечных волокон различны. Длительность потенциала действия скелетных мышц составляет несколько миллисекунд, и сокращение их начинается после завершения развития потенциала действия. В миокарде потенциал действия и сокращение в значительной степени перекрываются во времени. Потенциал действия клеток миокарда заканчивается после начала фазы расслабления. Поскольку последующее сокращение может возникнуть только в результате очередного возбуждения, а это возбуждение в свою очередь возможно лишь по окончании периода абсолютной рефрактерности предшествующего потенциала действия, сердечная мышца в отличие от скелетной не может отвечать на частые раздражения суммацией одиночных сокращений, или тетанусом. Это свойство миокарда имеет большое значение для реализации нагнетательной функции сердца: с одной стороны - тетаническое сокращение, продолжающееся больше периода изгнания крови, препятствовало бы наполнению сердца, с другой - тетаническое сокращение сердца эквивалентно его остановке.
Невозможность сердечной мышцы давать тетанические сокращения заставило детально проанализировать вопрос о механизмах регуляции силы сердечных сокращений. Как было отмечено, сократимость сердца не может регулироваться путем суммации одиночных сокращений, сократимость миокарда в отличие от скелетных мышц, не может изменяться путем включения различного числа моторных единиц, так как миокард представляет собой функциональный синцитий, и в каждом его сокращении участвуют все волокна. Однако, эти несколько невыгодные с физиологической точки зрения особенности компенсируются тем, что в миокарде возможность регуляции сократимости обеспечивается путем направленного изменения процессов возбуждения и электромеханического сопряжения.
Как организован механизм электромеханического сопряжения в миокарде? У человека и у млекопитающих структуры, отвечающие за электромеханическое сопряжение в скелетных мышцах, в основном, имеются и в волокнах сердца. Для миокарда характерна система поперечных трубочек (Т-система); особенно хорошо она развита в желудочках, где эти трубочки образуют продольные ответвления. Напротив, система продольных трубочек, служащих внутриклеточным резервуаром Са2+, в мышце сердца выражена в меньшей степени, чем в скелетных мышцах. Как структурные, так и функциональные особенности миокарда свидетельствуют в пользу тесной взаимосвязи между внутриклеточными депо Са2+ и внеклеточной средой. Ключевым событием в сокращении служит вход в клетку Са2+во время потенциала действия. Значение входного кальциевого тока состоит не только в том, что он увеличивает длительность потенциала действия и, как следствие, рефракторного периода: перемещение Са2+ из наружной среды в клетку создает условия для регуляции силы сокращения, так как чем больше кальция оказывается вблизи актина и миозина, тем сильнее сокращается

Рис. 3. Схема, иллюстрирующая соотношение между потенциалом действия (возбуждением) миокардиального волокна, током кальция и активацией сократительного аппарата. Начало сокращения связано с выходом кальция в зону актина и миозина из продольных трубочек в ходе деполяризации мембраны. Кальций, поступающий в кардиомиоцит через кальциевые каналы в фазу плато потенциала действия кардиомиоцита, пополняет запасы кальция в продольных трубочках.
На концентрацию кальция, активирующего контрактильный механизм, существенно влияет его количество в продольных трубочках, при этом показано, что значительная часть входящего в клетку Са2+ пополняет его запасы, обеспечивая достаточную эффективность очередных сокращений.
Таким образом, потенциал действия влияет на сократимость по меньшей мере двумя путями. 1. Он играет роль пускового механизма, вызывающего сокращение путем высвобождения Са2+ преимущественно из внутриклеточных депо. 2. Он пополняет запасы Са2+, создавая благоприятные условия для последующих сокращений.
Как выяснилось, целый ряд агентов оказывает значимое влияние на сокращение миокарда, изменяя длительность потенциала действия, что отражается на поступлении Са2+ внутрь миокардиоцитов. Например, ацетилхолин, выделяющий в окончаниях блуждающего нерва, уменьшая продолжительность потенциалов действия предсердной мышцы, параллельно усиливая проницаемость поверхностной мембраны клеток синоатриального узла для калия, вызывая тем самым их гиперполяризацию и уменьшение входного тока Са2+, вызывает уменьшение частоты и силы сердечных сокращений (отрицательный хронотропный, инотропный, дромотропный эффекты - см. ниже.). Напротив, норадреналин, выделяющийся в окончаниях симпатических нервных волокон, усиливая проницаемость для Са2+ , вызывает повышение частоты и увеличение силы сердечных сокращений (положительный хронотропный, инотропный, дромотропный эффекты - см. ниже).
Так называемый феномен лестницы (нарастание силы сокращений при их возобновлении после временной остановки) также связан с наращиванием внутриклеточной фракции Са2+. Сила сокращения сердца быстро изменяется при изменении содержания Са2+ во внеклеточной жидкости. Удаление Са2+ из внешней среды приводит к полному электромеханическому разобщению. Ряд веществ, блокирующих вход Са2+ во время потенциала действия, оказывает такой же эффект, как и удаление Са2+ из внешней среды. К таким веществам относятся двухвалентные катионы (Ni2+, Со2+, Mn2+), а также некоторые органические соединения - антагонисты кальция (верапамил, нифедипин). При повышении внеклеточного содержания Са2+ или при действии фармакологических препаратов, увеличивающих вход Са2+ во время развития потенциала действия, сократимость сердца увеличивается. Механизм действия сердечных гликозидов (дигоксин, строфантин) частично связан именно с увеличением внутриклеточной фракции Са2+.
4. Биофизические основы электрокардиографии
Особенности распространения возбуждения по сердцу отображаются в электрокардиограмме (ЭКГ), которая имеет характерную форму (рис. 4). Элементарной моделью генератора ЭКГ является электрический диполь. При распространении возбуждения по миокарду формируется множество диполей, которые закономерно изменяют свои количественные характеристики и направление. В каждый момент времени возникают новые диполи, исчезают прежние. В результате на поверхности сердечной мышцы создается сложная мозаика распределения электрических потенциалов. Результирующий дипольный момент миокарда, равный векторной сумме отдельных диполей, получил название интегрального электрического вектора сердца (ИЭВС). Ритмичный характер автоматизма водителя ритма, а также передача возбуждения посредством электрических синапсов обусловливают синфазность возбудительного процесса в миокардиальных волокнах. Поэтому ИЭВС имеет сравнительно большую амплитуду прежде всего при деполяризации желудочков, чем создается высокий уровень биопотенциалов, отражающих сердечную деятельность даже на поверхности тела. Ежемоментно амплитуда и направление ИЭВС различны. Измеряя их, врач получает сведения о движении волны возбуждения по сердцу, что позволяет ему оценить свойства миокарда и в случае нарушений сердечной деятельности понять их природу.

Рис. 4. Записи ЭКГ в трех стандартных отведениях и схема, показывающая связь между распространением возбуждения в сердце и возникновением некоторых зубцов электрокардиограммы (по Барду).
При одновременной подаче электрических потенциалов, отражающих распространение возбуждения по миокарду, на пластины “Х” и “У” электроннолучевого осциллографа на его экране возникают фигуры Лиссажу, образующиеся в результате сложения взаимно перпендикулярных колебаний регистрируемых потенциалов (рис. 5).
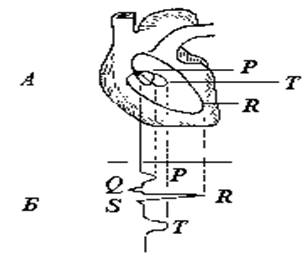
Рис. 5. Соотношение векторэлектрокардиограммы (А) и электрокардиограммы (Б).
В данном случае фигуры Лиссажу представляют собой траектории движения ИЭВС, описываемые его концом на плоскости, перпендикулярной направлению распространения возбуждения по миокарду. Такое исследование электрической активности сердца называется векторэлектрокардиоскопией (ВЭКС). На векторэлектрокардиограмме выделяют обычно три эллипсоподобные фигуры. Самая мелкая из них отображает деполяризацию предсердий, самая крупная - деполяризацию желудочкой, средняя - их реполяризацию.
Амплитуду вектора оценивают посредством измерения его проекций на координатные оси. Любое из отведений ЭКГ есть не что иное как проекция интегрального электрического вектора сердца на соответствующую координатную ось.
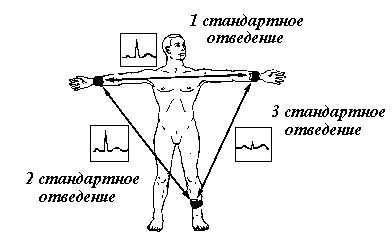
Рис. 6. Схема стандартных электрокардиографических отведений (треугольник Эйнтховена)
В разнообразных способах отведения ЭКГ воплощены различные системы координат. Широко распространена координатная система, элементом которой является равносторонний треугольник (рис. 6). Она предложена основоположником электрокардиографии В. Эйнтховеном и получила название системы стандартных отведений. При ее практической реализации накладывают электроды на левую ногу и обе руки. Первым отведением считают регистрацию разности потенциалов между правой и левой рукой, вторым - между правой рукой и левой ногой, третьим - между левой рукой н левой ногой. Применяются и другие способы отведений ЭКГ, для чего используются иные координатные системы. Вне зависимости от способа отведения ЭКГ ее зубцы отражают электрическую активность сердца в соответствующий момент сердечной де-ятельности: зубец Р формируется при возбуждении предсердий, комплекс QRS - при возбуждении желудочков, зубец Т - при их реполяризации. Таким образом, отклонения от нормы, обнаруживаемые в том или ином элементе ЭКГ, можно адресовать соответствующему отделу сердца.
Важным параметром ЭКГ служат временные интервалы. По ним оценивают скорость распространения возбуждения в каждом из отделов проводящей системы сердца. Изменения скорости проведения наблюдаются при повреждениях сердечной мышцы. Даже мелкий очаг поражения миокарда (диаметром 5-10 мкм) вызывает задержку в распространении возбуждения на 0,1 мс.
В стандартных отведениях зубец Р имеет амплитуду не более 0,25 мВ и длительность 0,07-0,10 с. Интервал PQ, отображающий атрио-вентрикулярную задержку, составляет 0,12-0,21 с при частоте сердечных сокращений порядка 70 /мин. Комплекс QRS наблюдается в течение всего времени, пока возбуждение распространяется по желудочкам - от 0,06 до 0,09 с. Зубец Q в трети наблюдений отсутствует в нормальной ЭКГ, а когда обнаруживается, не превышает 0,25 мВ. Зубец R обладает максимальной амплитудой среди других элементов ЭКГ. Она составляет 0,6-1,6 мВ. Зубец S -часто отсутствует, но иногда достигает 0,6 мВ. Он появляется в тот момент, когда деполяризация охватывает участки желудочков, прилежащие к предсердиям. Основание желудочков возбуждается в последнюю очередь. Сегмент S-T при пульсе 65-70 /мин составляет 0,12 с. Длительность зубца Т немного больше - от 0,12 до 0,16 с. Его амплитуда находится в пределах 0,25-0,6 мВ.
Зубец Р возникает на ЭКГ примерно за 0,02 с до начала сокращения предсердий, а комплекс QRS - за 0,04 с до начала сокращения желудочков. Следовательно, электрические проявления возбуждения предшествуют меха-ническим.
Имея ряд ЭКГ, по крайней мере, две, снятые в 1 и 3 отведениях, можно синтезировать ИЭВС. В медицинской литературе его называют электрической осью сердца - отрезок прямой, соединяющий два сечения миокарда, облада-ющих в данный момент наибольшей разностью потенциалов. Направление электрической оси сердца в ходе распространения возбуждения по миокарду постоянно изменяется. Принято определять среднюю электрическую ось сердца. Так называют вектор, который можно построить в промежутке между началом и окончанием деполяризации миокарда желудочков. По расположению средняя электрическая ось близка анатомической оси сердца. Построение средней электрической оси дает представление о положении сердца в грудной полости. Отклонения оси вправо или влево служат признаками изменений миокарда соответствующего желудочка.
