

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
![]()
![]()
![]()
|

|
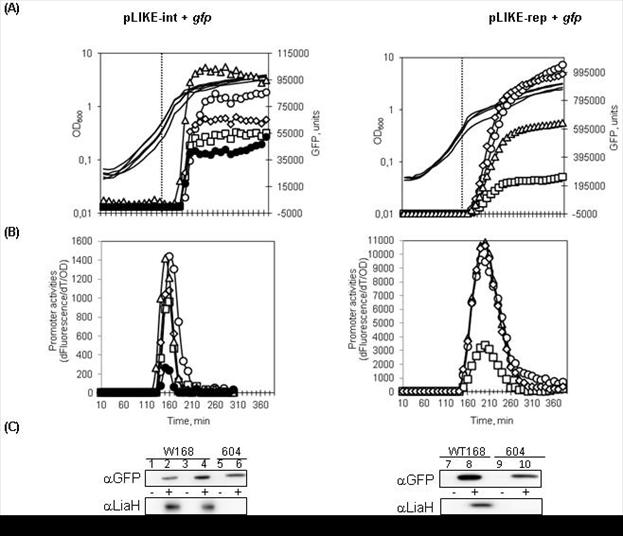
Рис. 11. Рост, показания флуоресценции (А) и активность промотора (B) в штаммах несущих трансляционные конструкции PliaI-gfpmut1 и PliaI(opt)-gfpmut1. Кривые роста показаны без символов. Обозначения экспрессии в разных штамах: (○) (W168), (□) (TMB604), (◊) (TMB1151), (Δ) (TMB1152) (●) (TMB408). Вертикальной пунктирной линией показано время добавление индуктора (OD600~0.4-0.4, бацитрацин в концентрации 30 мкг/мл).
(С) Western blot анализ цитоплазматической фракции клеток, содержащих полученные конструкции с использованием антител к белкам LiaH и GFP. Обозначения: 1,2 - экспрессия конструкций с нативного промотора PliaI, 3-10 – экспрессия конструкций PliaI(opt) в отсутствии (-) и присутствии (+) бацитрацина (концентрация антибиотика 30 мкг/мл).
Таким образом, результаты экспериментов показали положительный эффект оптимизации последовательности Шайна-Дальгарно в промоторе PliaI новой LIKE-системы для экспрессии рекомбиантных белков. Кроме того, наблюдали небольшой положительный эффект делетирования терминатора liaIH-оперона. Для подтверждения достоверности повышения выхода рекомбинантного белка GFP, провели Western-blot анализ с использованием антител к данному белку, а также к белку LiaH. (рис. 11 C). Показали, что оба белка не обнаруживаются в клетках до индукции бацитрацином, что свидетельствует о строгом контроле активности PliaI промотора в клетках btilis. После добавления антибиотика в среду белки GFP и LiaH накапливались в клетках в различной концентрации, что определялось генотипом полученных конструкций (рис. 10, рис. 11).
Таким образом, нами разработан эффективный инструментарий – LIKE-система экспрессии btilis, основанная на стресс индуцибельном промоторе liaIH-оперона и ряд штаммов с делециями liaIH-liaGFSR регулона на хромосоме. Разработанная система позволяет направленно экспрессировать интересуемые гены как в составе плазмидного элемента – репликативная конструкция (pLIKE-rep), так и в составе генома клеток – интегративная конструкция (pLIKE-int). Показано, что максимальный уровень экспрессии PliaI промотора в составе pLIKE-int вектора происходит через 20-30 мин после добавления антибиотика, в то время как в составе вектора pLIKE-rep – через 60 мин, при этом его уровень в 10 раз выше, что объясняется мультикопийностью репликативной плазмиды.
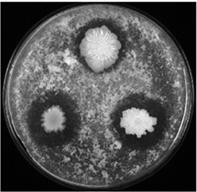
LIKE-система для продукции генов сериновых протеаз. Для тестирования новой системы экспрессии бацилл нами использованы гены сериновых протеаз B. pumilis 3-19. Известен потенциал этих ферментов в биотехнологии и медицине [, с совт., 1998], что определяет необходимость получения чистого продукта в большом количестве. Гены глутамилэндопептидазы и субтилизиноподобной протеиназы клонировали в репликативный вектор pLIKE-rep под контроль PliaI(opt) промотора. Полученными плазмидами трансформировали беспротеазный штамм btilis BG2036 и исследовали активность протеаз на молочном агаре и с применением специфических субстратов. Как видно из рисунка 12 (а), после 16 ч инкубирования рекобинантных штаммов на молочном агаре с добавлением бацитрацина, клетки эффективно продуцируют протеазы в среду.

![]()
![]()
![]()

(б) (с)
![]()
![]()
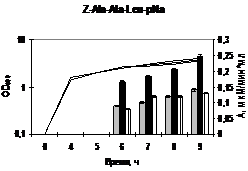
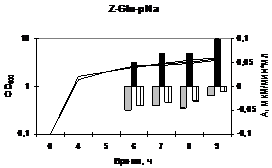
Рис. 12. (а) Колонии исходного штамма B. pumilus 3-19 и рекомбинантных штаммов B. subtilis, содержащих конструкции PliaI-aprBp и PliaI-gseBp на молочном агаре с добавлением антибиотика бацитрацина (30 мкг/мл) после 16 ч инкубирования.
(б) Активность субтилизиноподобной протеиназы исходного B. pumilus 3-19 и рекомбинантного B. subtilis (PliaI-aprBp) штаммов.
(с) Активность глутамилэндопептидазы исходного B. pumilus 3-19 и рекомбинантного B. subtilis (PliaI-gseBp) штаммов.
Исходный штамм показан в белом цвете. Рекомбинантные штаммы при добавлении бацитрацина показаны в чёрном цвете, без добавления – в сером цвете. Стрелочкой указано время добавления индуктора.
Анализ рекомбинантных штаммов, содержащих гены сериновых протеаз под контролем индуцируемого PliaI(opt) промотора на специфических субстратах показал, что активность ферментов повышалась в 2 раза. Полученный результат по активности ферментов можно объяснить тем, что для продукции протеолитических ферментов требуется формирование Sec-каналов и белков для эффективной секреции протеаз в среду. На это указывают результаты культивирования штаммов на молочном агаре. Вероятно, для дальнейшего повышения уровня белка в КЖ необходима оптимизация системы. Таким образом, новая LIKE-система экспрессии бацилл эффективна для производства секреторных рекомбинантных белков.
Итак, основные характеристики новой разработанной LIKE экспрессионной системы - (1) отсутствие экспрессии PliaI промотора если в среду не добавлен индуктор, (2) быстрая и высокая (100-1000-кратная) экспрессия гена под контролем PliaI промотора, (3) возможность использования различных индукторов, (4) позитивное влияние разработанных рекомбинантных штаммов на экспрессию PliaI промотора, (5) возможность применения системы для продукции ферментов используемых в медицине и пищевой индустрии – предполагают её использование, как альтернативной и конкурентоспособной экспрессионной системы btilis.
ВЫВОДЫ
1. Установлена минимальная область регуляции генов протеаз штамма Bacillus pumilus: для гена субтилизиноподобной протеиназы промотор составляет 445 п. о.; для гена глутамилэндопептидазы - 150 п. о.
2. Установлено, что для репортёрных конструкций с минимальной областью регуляции протеаз факторы транскрипции DegU и Spo0A оказывают позитивное влияние, тогда как альтернативный сигма фактор транскрипции SigD – негативное.
3. Инактивация генов протеолитических ферментов приводит к плейотропному эффекту: функционально-активные протеиназы участвуют в формировании пула гидролитических ферментов.
4. Сконструирована новая LIKE-система экспрессии на основе антибиотико-индуцибельного промотора LiaRS двухкомпонентной системы Bacillus subtilis.
5. В составе новой экспрессионной системы экспрессия репортёрного гена gfp повышалась в среднем в 6 раз, а генов сериновых протеиназ в 2 раза.
Публикации по теме диссертации в изданиях, рекомендованных ВАК:
1. Bacillus intermedius с нокаутированным геном субтилизиноподобной протеиназы / , , // Учен. Зап. Казан. ун-та. Сер. Естеств. науки. -2010. - Т. 152, кн. 4. - С. 143-155. (перечень ВАК), автора – 0,5 п. л.
2. Тойменцева делений в промоторе на экспрессию гена глутамилэндопептидазы Bacillus intermedins / , , // Учен. зап. Казан. ун-та. Сер. Естеств. науки. -2010.-Т. 152. кн. 3. - С. 149-158. (перечень ВАК), автора – 0,5 п. л.
3. Черёмин A. M. Выделение регуляторных белков, активирующихся в условиях ограниченного роста бацилл / A. M. Черёмин, , , пова // Учен. зап. Казан. ун-та. Сер. Естеств. науки. -2010. - Т. 152. кн. 4. - С. 156-168. (перечень ВАК), автора – 0,3 п. л.
4. Шарипова филогенетическое положение штамма Bacillus intermedius 3-19 / , , // Микробиология. -2011. - Т. 80. -№ 3. - С. 424-426. (перечень ВАК) автора – 0,05 п. л.
Другие публикации по теме диссертации:
5. Тойменцева гена глутамилэндопептидазы Bacillus intermedius модифицированного в области промотора в клетках Bacillus subtilis / , , // Материалы докладов XIV международной научной конференции студентов, аспирантов и молодых учёных «Ломоносов» Международный молодёжный научный форум «Ломоносов -2007». - Москва, 2007. –С.45.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 |
