
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Каталитическое определение пирофосфата и АМФ
Тенденции секвенирования
Разрабатываемые в последние годы новые методы секвенирования нуклеиновых кислот можно разделить на две основные группы – мономолекулярные и мультимолекулярные. Первые требуют сложного приборного оснащения. Кроме того, до сих пор никому не удалось экспериментально доказать их работоспособность. Всё секвенирование пока-что проводят при помощи мультимолекулярных методов, основанных, как правило, на электрофоретическом разделении смеси молекул ДНК /1/. Значительно реже применяется РНК-секвенирование, при котором используют электрофоретическое разделение не ДНК, а РНК /2, 3/. В последнее время внимание разработчиков новых методов привлекает также пиросеквенирование, не требующее электрофоретического анализа продуктов реакции /4/.
Ускорение и удешевление секвенирования достигается в основном его миниатюризацией и автоматизацией. Первоначально секвенирование по Сэнгеру проводили на обычных приборах для вертикального электрофореза с радиоавтографическим выявлением разделившихся в полиакриламидном геле (ПААГ) молекул ДНК /5/. Секвенаторы первого поколения представляли собой просто большие электрофоретические приборы /6/, так как для уменьшения их размеров необходимо было значительно повысить удельную активность включаемой в ДНК радиоактивной метки и уменьшить энергию излучаемых электронов. Замена метки P32 изотопом P31 позволила вместо большого электрофоретического прибора использовать небольшое предметное стекло для микроскопии /7/, но автоматизировать такую технологию очень трудно. Кроме того, подобный подход требует использования четырёх электрофоретических дорожек – по одной на каждый нуклеотид.
Разработать «однодорожечное» секвенирование удалось благодаря применению разноцветных флуоресцентных меток /8/. Первые флуоресцентные секвенаторы также работали с плоским ПААГ /9/, но сейчас чаще используется капиллярный электрофорез. Совершенствование подобных приборов сводится к простому наращиванию количества капилляров /10/ или к их замене стеклянным блоком, содержащим множество капиллярных канавок /11/. Это ускоряет и удешевляет секвенирование, но стоимость самих капиллярных секвенаторов чрезмерно высока. Даже простейшие модели стоят около 100 тысяч долларов.
Технология пиросеквенирования в основном заключается в промывании препаратов ДНК растворами, каждый из которых содержит один из четырёх дезоксинуклеозидтрифосфатов (дНТФ) /11/. Включение нуклеозида в наращиваемую полимеразой нить ДНК приводит к выделению пирофосфата (PPi). При отсутствии такого включения пирофосфат не выделяется. Новизну данному методу придаёт сочетание двух ферментативных реакций. Первая преобразует PPi в аденозинтрифосфат (ATP), а вторая катализирует ATP-зависимое окисление люциферина, приводящее к излучению кванта света. Квантовый выход данной реакции довольно низок - 1 фотон на 10000 молекул PPi /12/, но высокая чувствительность люминометрии позволила разработать технологию параллельного секвенирования сотен тысяч проб объёмом 75 пиколитров /13/. Тем не менее, и при таких мизерных объёмах проб применение данного метода в значительной степени лимитируется стоимостью реагентов.
Для снижения расхода и стоимости реагентов необходима дальнейшая миниатюризация технологии пиросеквенирования, а для этого, в свою очередь, необходимо повысить чувствительность определения PPi. Не меньшее значение могут иметь чувствительные реакции определения аденозинмонофосфата (AMP), использование которых позволило бы заменить радиоавтографию более удобными методами проявления электрофореграмм, а также уменьшить расход реагентов и размеры разделяющих гелей при РНК-секвенировании. Это определяет актуальность рассмотрения ферментативных реакций, позволяющих выявлять следовые количества пирофосфата и AMP.
Сульфотрансферазная реакция
Первой стадией люминометрического выявления пирофосфата является синтез ATP из 5’- аденозинфосфосульфата (APS), катализируемый сульфурилазой ((sulfate adenylyltransferase, EC 2.7.7.4). Равновесие этой реакции смещено в сторону образования ATP, но in vivo она служит для синтеза APS – промежуточного метаболита, участвующего в усвоении неорганического сульфата /15/.
Следующим соединением этой метаболической цепочки является PAPS (3’-фосфоаденозин-5’-фосфосульфат), синтез которого катализируется APS-киназой (adenylyl-sulfate kinase, EC 2.7.1.25). У большинства бактерий, животных и растений эти ферменты образуют единый комплекс, последовательно катализирующий обе реакции /16, 17/, поэтому при пиросеквенировании используют дрожжевую сульфурилазу, не сцепленную с APS-киназой. Если использовать более распространённые комплексы сульфурилаз с APS-киназами, то ATP будет расходоваться на синтез PAPS.
PAPS во многих ферментативных реакциях является донором сульфатной группы, перенос которой приводит к образованию 3’-фосфоаденозин-5’-фосфата (PAP). Такие реакции обратимы и катализируются сульфотрансферазами. Для определения активности этих ферментов предложено несколько очень чувствительных методов, основанных на переносе сульфатных групп между различными фенольными и нафтольными субстратами /18, 19/. При этом пара PAP-PAPS служит кофактором сульфотрансферазы, т. е. работает в режиме катализатора. В таком случае для выявления пирофосфата можно использовать следующую последовательность реакций:
1. PPi + APS ® SO42- + ATP
2. ATP + APS ® ADP + PAPS
3. Сульфотрансферазная реакция (рис. 1)
Сульфотрансферазная реакция
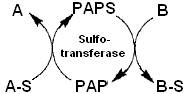
Рис. 1
Чувствительность этой реакции определяется характером используемого субстрата. Для колориметрического определения подходит п-нитрофенилсульфат /18/. Чувствительный флуориметрический метод определения сульфотрансфераз был разработан с применением 2-нафтилсульфата /19/. Добиться значительного повышения чувствительности сульфотрансферазной системы позволила бы разработка хемилюминесцентных субстратов, способных после переноса сульфогруппы окисляться с излучением фотонов. Сейчас подобные субстраты применяются для определения фосфатаз и пероксидаз /20/. Типичным примером такого субстрата является Lumigen APS-5 (сокращение «APS» в данном случае означает Alkaline Phosphatase Substrate, рис.2) /21/.
Lumigen APS-5
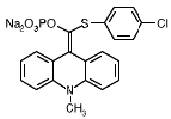
Рис. 2
Хемилюминесцентная индикация PPi, основанная на сульфотрансферазной реакции, по чувствительности может значительно превосходить люциферазную систему, но пока трудно предсказать, какими свойствами будет обладать, например, аналог Lumigen APS-5, содержащий сульфатную группу вместо фосфатной, и сможет ли такое соединение служить субстратом для сульфотрансфераз. Возможно, более подходящими для них окажутся сульфопроизводные хемилюминесцентных субстратов пероксидазного окисления /22/.
Повысить чувствительность сульфотрансферазного выявления PPi можно также регенерацией ATP из ADP. Подобную регенерацию используют в системах бесклеточного синтеза белков и в некоторых других биотехнологических процессах.
Регенерация ATP
Наибольшее распространение получили три системы регенерации ATP из ADP, в которых донорами фосфатных групп служат высокоэнергетические соединения:
1. Ацетилфосфат – ацетилкиназа (AcP-AcK) /23/;
2. Креатинфосфат – креатинкиназа (CP-CK) /24/;
3. Фосфоенолпируват – пируваткиназа (PEP-PK) /25/.
Каждая из этих систем обладает как достоинствами, так и недостатками. Отличаются они, например, по стабильности субстратов в водных растворах. Ацетилфосфат (AcP) гидролизуется наполовину примерно за 8 часов, креатинфосфат (CP) – за 12 дней, а фосфоенолпируват (PEP) – за 3 месяца /26/. Но PEP лидирует не только по стабильности, но и по стоимости (38 $/mM), тогда как AcP стоит 7,9 $/mM, а CP – 9,7 $/mM (по каталогу Sigma Chemical Co., 2003). По доступности лидирует, по-видимому, CP. Его можно приобрести в аптеке под торговым названием «НЕОТОН» (Alfa Wasserman, Италия). Ещё одним достоинством системы CP-CK является высокое сродство креатинкиназы к ADP (Km=0,051 mM). У пируваткиназы и ацетилкиназы Km значительно выше (0,3 и 1,5 mM) /26/. Равновесие всех этих реакций смещено в сторону образования ATP (рис. 3).
Креатинкиназная реакция /26/
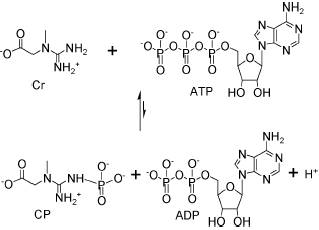
Рис. 3
Регенерация ATP в сульфотрансферазной системе определения PPi может значительно повысить чувствительность метода, так как вместо простого преобразования пирофосфата в эквимолярное количество PAPS будет идти накопление PAPS, пропорциональное количеству циклов регенерации ATP.
Одно из достоинств сульфотрансферазной системы связано с тем, что содержащийся в ней APS является общим субстратом и для сульфурилазного синтеза ATP из PPi, и для киназного синтеза PAPS. Это подходит для выявления пирофосфата, но APS легко гидролизуется до аденозинмонофосфата, поэтому данную систему невозможно использовать для РНК-секвенирования, требующего выявления следовых количеств AMP. В последнем случае более подходящим бифункциональным субстратом может стать фосфоенолпируват, который способен участвовать в регенерации APT не только из ADP, но и из AMP. В последнем случае реакция катализируется ферментом PPDK (pyruvate orthophosphate dikinase или ATP:pyruvate, orthophosphate phosphotransferase, EC 2.7.9.1) и приводит к синтезу ATP из AMP и пирофосфата:
AMP + Phosphoenolpyruvate + PPi ® ATP + Pyruvate + Pi
Дикиназный путь перевода PPi в ATP разработан совсем недавно и ещё не применялся в пиросеквенировании, хотя замена APS-сульфурилазной реакции PEP-дикиназной, судя по всему, вполне возможна. В продаже уже имеется и сам фермент, и наборы для люминометрического определения AMP и ATP, в которых используется эта реакция (Kikkoman Corporation, Japan) /27/. PPDK синтезируют некоторые клостридии, но в тест-системах используется клонированный в E. сoli термостабильный рекомбинантный фермент из Microbispora rosea /28/. Равновесие дикиназной реакции в присутствии пирофосфатазы смещено в сторону синтеза фосфоенолпирувата, что можно использовать для получения дешёвых препаратов PEP из ATP и пирувата /29/.
Для выявления AMP и регенерации ATP пригодны не только реакции, идущие с образованием ATP из AMP и PPi, но и синтез ADP из AMP и полифосфата, катализируемый полифосфат:AMP фосфотрансферазой (PPT) из Acinetobacter johnsonii /30/:
polyPn + AMP → ADP + polyPn−1
Эта реакция была использована для разработки двухферментной системы регенерации ATP из AMP и полифосфата /30/. Данная система в дополнение к PPT содержит аденилаткиназу (AK), обеспечивающую синтез ATP из ADP:
ADP + ADP → AMP + ATP
Последняя реакция позволяет не только регенерировать, но и амплифицировать ATP, что можно использовать для разработки очень чувствительных методов определения различных веществ.
Амплификация ATP
Аденилаткиназный синтез ATP из ADP обратим, поэтому реакцию можно записать следующим образом:
ATP + AMP → ADP + ADP
Если данная реакция будет идти в среде, содержащей не только аденилаткиназу и избыток AMP (AMP-AK), но и систему регенерации ATP из ADP (например, PEP-PK), то количество ATP должно нарастать в геометрической прогрессии.
ATP ®AMP-AK®2ADP®PEP-PK® 2ATP ®AMP-AK®4ADP®PEP-PK® 4ATP ® …
Для дальнейшего рассмотрения и обсуждения системы амплификации (мультипликации) ATP желательно подобрать для неё какое-нибудь краткое обозначение. Сокращение «AMP-AK + PEP-PK» выговорить очень трудно, да и реагенты могут использоваться другие (СP-СK, AcP-AcK и т. п.). Лучше подобрать трёхбуквенное обозначение типа PCR, отражающее суть реакции, т. е. что-нибудь вроде ACR (Adenilatkinase Chain Reaction). Возможно, более подходящей аббревиатурой будет AKM (АденилатКиназная Мультипликация).
Затравкой для АКМ может служить всего одна молекула ATP или ADP, поэтому возможность разработки метода определения PPi для пиросеквенирования будет зависеть в основном от специфичности аденилаткиназы, а не от чувствительности реакции. Не исключено, что подобрать фермент, не вовлекающий в реакцию ни один из dNTP, будет очень трудно. Подобную же проблему пришлось решать и при разработке пиросеквенирования, когда выяснилось, что светлячковая люцифераза даёт фоновое свечение с dATP. Для устранения этого фона пришлось dATP заменить модифицированным аналогом, содержащим тиогруппу в альфа-положении / /.
Аденилаткиназы присутствуют во всех организмах, причём у животных они отличаются как тканевой, так и субстратной специфичностью. Например, аденилаткиназа 3 (AK3) из сердца быка предпочитает вместо ATP использовать GTP и имеет систематическое название GTP:AMP фосфотрансфераза (EC 2.7.4.10):
GTP + AMP = GDP+ADP
Для РНК-секвенирования обычно используется РНК-полимераза фага Т7, которая синтезирует транскрипты с GTP на 5’-конце / /. После РНКазной обработки в присутствии AK3 этот нуклеозид должен запускать амплификацию ATP, поэтому гуанозинтрифосфатная метка молекул в системе AKМ может заменить радиоактивную метку P32, используемую в РНК-секвенировании.
Инициировать АКМ продуктами РНКазного расщепления, включающими AMP, можно и при помощи их фосфорилирования полифосфат:AMP фосфотрансферазой. Но в этом случае реакцию необходимо проводить в два этапа, так как присутствие в АКМ компонентов системы фосфорилирования АМP (полифосфат + PPT) вызовет неспецифическую амплификацию ATP. Если специфичность PPT и АК окажется не слишком высокой, то подобная двухэтапная система выявления продуктов расщепления нуклеиновых кислот подойдёт и для выявления ДНК.
В твердофазных иммуноферментных тест-системах очень важна чувствительность реакции, используемой для обнаружения метки. По этому параметру АКМ может оказаться вне конкуренции, так как для инициации данной каскадной реакции достаточно одной молекулы субстрата или фермента, взаимодействие которых способно привести к синтезу ATP или ADP. Некоторые подобные ферменты и их субстраты рассматривались выше, но в другом контексте:
· сульфурилаза (PPi + APS);
· дикиназа (PPi + PEP);
· PPT (полифосфат + AMP).
В этот перечень можно включить и пирофосфатазу, способную синтезировать пирофосфат из фосфата благодаря обратимости реакции. Обратимость характерна для большинства каталитических реакций, поэтому способностью синтезировать ADP и запускать каскадный синтез ATP должны обладать некоторые фосфокиназы, обычно использующие ADP в качестве донора фосфатной группы / #/.
Чувствительность АКМ позволяет использовать в качестве метки для антигенов и антител белки или реагенты, способные высвобождать всего одну молекулу ATP. Например, ДНК-лигаза бактериофага T4 при взаимодействии с ATP аденилируется. Если таким аденилатом пометить антитела или антигены, то они смогут высвобождать молекулы ATP при добавлении пирофосфата. Список подобных аденилатных меток может быть довольно обширен / /. Не исключено, что ДНК-лигаза или какой-нибудь другой аденилированный фермент сможет заменить сульфурилазу и APS в пиросеквенировании. Формирование аденилатных комплексов, по-видимому, подойдёт и для иммобилизации ATP при проявлении гелей, содержащих РНК. Тогда метод, заменяющий радиоавтографию, можно будет назвать аденографией.
Кофакторные реакции
Использовать AKM в пиросеквенировании будет очень сложно из-за необходимости после каждой замены dNTP удалять все молекулы, способные инициировать цепную реакцию (ATP, ADP и PPi). В случае линейного усиления входного сигнала (PPi) удаление исходного и конечного продуктов реакции менее критично, но повышаются требования к чувствительности всей системы. Сульфотрансферазная реакция без люминометрической регистрации конечных продуктов вряд ли сможет конкурировать по чувствительности с обычной люциферазной системой, но кроме киназного синтеза PAPS можно использовать и другие реакции ATP, приводящие к синтезу кофакторов ферментов. Некоторые из них способны активировать хемилюминесценцию. Вот несколько примеров подобных реакций:
· NAD ® NADP
· пиридоксаль ® пиридоксальфосфат
· рибофлавин ® FMN (riboflavin kinase, EC 2.7.1.26 )
· FMN ® FAD (ATP:FMN adenylyltransferase, EC 2.7.7.2)
В этом списке первые три реакции катализируются фосфокиназами, образующими в качестве побочного продукта ADP, а последняя – аденилилтрансферазой, высвобождающей PPi. Общая чувствительность ферментных систем, построенных на использовании подобных реакций, будет зависеть от скорости регенерации ATP из ADP или PPi, а также от чувствительности ферментативных реакций, в которых участвуют активированные кофакторы.
NADP служит кофактором для многих дегидрогеназ и его легко обнаружить по флуоресценции собственной восстановленной формы или по яркой окраске нерастворимых соединений, образующихся в результате восстановления тетразоловых красителей. Наиболее известна глюкозо-6-фосфат–дегидрогеназная реакция, широко применяемая для определения концентрации глюкозы / /. Служить кофакторами дегидрогеназных реакций способны и фосфорилированные производные витамина B6 / , , /. Для определения подобных соединений можно подобрать чувствительные денситометрические или флуориметрические методы, но они значительно уступают люминометрии, поэтому наибольший интерес вызывает FMN, участвующий в хемилюминесцентном катализе окисления альдегидов некоторыми бактериями.
1. Секвенирование ДНК / , , - 1999. - М., Наука - 429 с.
2. Specific termination of RNA polymerase synthesis as a method of RNA and DNA sequencing / Axelrod V. D., Vartikyan R. M., Aivazashvili V. A., Beabealashvili R. S. // Nucleic Acids Res. – 1978. – Vol.5, №10. – P.3549-3563.
3. Axelrod V. D., Kramer F. R. Transcription from bacteriophage T7 and SP6 RNA polymerase promoters in the presence of 3'-deoxyribonucleoside 5'-triphosphate chain terminators / Biochemistry. – 1985. - Vol.24, №21. - P.5716-5723
4. www.
5. Сэнгер
6. «Macrophor»
7.
8. M. Hankapiller
13. www.454.com
15. http://phm. bio. msu. ru/edocs/micro/16.html - усвоение сульфата
16. Arand M., Robertson L. W., Oesch F. A fluorometric assay for quantitating phenol sulfotransferase activities in homogenates of cells and tissues / Anal Biochem. – 1987. – Vol.163, №2. – P.546-551.
18. A simple colorimetric assay for phenotyping the major human thermostable phenol sulfotransferase (SULT1A1) using platelet cytosols / L. T. Frame, S. Ozawa, S. A. Nowell et al. // ?. - 2000. - Vol.28, Issue 9. – P.1063-1068. (http://dmd. aspetjournals. org/cgi/content/full/28/9/1063)
19. A fluorometric assay for quantitating phenol sulfotransferase activities in homogenates of cells and tissues / Arand M, Robertson LW, Oesch F.// Anal Biochem. 1987 Jun;163(2):546-51.
20. http://www.
21. http://www. /documents/lumigenaps. shtml
22. http://www. /documents/pdf/ChemifluorPres. pdf
23.
26. Creatine phosphate-creatine kinase in enzymatic synthesis of glycoconjugates / J. Zhang, B. Wu, Y. Zhang, P. Kowal, P. G. Wang // Organic Letters. – 2003. – Vol.5, №15. – P.2583-2586.
27.
28. Pyruvate phosphate dikinase from a thermophilic actinomyces Microbispora rosea subsp. aerata : purification, characterization and molecular cloning of the gene / Naoki Eisaki, Hiroki Tatsumi, Seiji Murakami and Tatsuo Horiuchi // BBA - Proteins and Proteomics. – 1999. - Vol.1431, №2. – P.363-373. [Abstract].
29. Enzymatic synthesis of radiolabeled phosphonoacetaldehyde / G. Zhang, K. N. Allen, D. Dunaway-Mariano // Anal. Biochem. – 2003. – Vol.322. – P.233–237.
30. Properties of polyphosphate:AMP phosphotransferase of Acinetobacter strain 210A / C. F.C. Bonting, G. J.J. Kortstee, A. J.B. Zehnder // J. Bacteriol. – 1991. – Vol.173. – P.6484-6488. [Medline].
31. Resnick S. M., Zehnder A. J.B. In vitro ATP regeneration from polyphosphate and AMP by polyphosphate:AMP phosphotransferase and adenylate kinase from Acinetobacter johnsonii 210A // Appl. Environ. Microbiol. – 2000. – Vol.66, №5. – P.2045–2051.
/#/ Biochemical Characterization, Cloning, and Sequencing of ADP-Dependent (AMP-Forming) Glucokinase from Two Hyperthermophilic Archaea, Pyrococcus furiosus and Thermococcus litoralis / Shinji Koga,* Issei Yoshioka,* Haruhiko Sakuraba,† Mamoru Takahashi,* Shinichi Sakasegawa,* Sakayu Shimizu,‡ and Toshihisa Ohshima // J. Biochem. Vol. 128, pp. 1079-1085 (2000), Regular paper; © 2000 by The Japanese Biochemical Society
Molecular characterization of Escherichia coli NAD kinase / Kawai S, Mori S, Mukai T, Hashimoto W, Murata K.// Eur J Biochem. 2001 Aug; 268(15): 4359-65.
… utilized ATP or other nucleoside triphosphates as phosphoryl donors for the phosphorylation of NAD, most efficiently at pH 7.5 and 60 degrees C.
Oka M, McCormick DB. Complete purification and general characterization of FAD synthetase from rat liver // J Biol Chem. 1987 May 25;262(15):7418-22
Kinetic analysis gave Km values of 9.6 microM for FMN and 53 microM for ATP.
Bowers-Komro DM, Yamada Y, McCormick DB. Substrate specificity and variables affecting efficiency of mammalian flavin adenine dinucleotide synthetase // Biochemistry. 1989 Oct 17;28(21):8439-46.
Nucleoside triphosphates other than ATP do not act as substrates or inhibitors
ATP + pyridoxal <=> ADP + pyridoxal 5'-phosphate
ATP + pyridoxine <=> ADP + pyridoxine-5'-phosphate
ATP + pyridoxamine <=> ADP + pyridoxamine 5'-phosphate
