
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Практическое занятие 2
Типы рецепторов сетчатки и механизм фоторецепции
(практическое занятие)
Цель занятия: Ознакомление с процессами, происходящими в сетчатке глаза при фоторецеции.
Аудиторное время: 2 часа.
1. Строение сетчатки глаза
Сетчатка состоит из трех слоев, каждый из которых содержит клетки определенного типа. Самый наружный (наиболее удаленный от центра глазного яблока) светочувствительный слой содержит фоторецепторы –палочки и колбочки, частично погруженные в пигментный слой сосудистой оболочки. Затем идет промежуточный слой, содержащий биполярные нейроны, которые связывают фоторецепторы с клетками третьего слоя. В этом же промежуточном слое находятся горизонтальные и амакриновые клетки, обеспечивающие литеральное торможение. Амакриновые клетки – это наиболее разнообразный тип нейронов сетчатки.
Третий слой – внутренний поверхностный слой – содержит ганглиозные клетки, дендриты которых соединены синапсами с биполярными клетками, а аксоны образуют зрительный нерв. Аксон – это отросток нервной клетки, по которому идут нервные импульсы. Синапс – это место контакта между двумя нейронами. Ганглиозная клетка – это нервная клетка (нейрон) сетчатки глаза, способная генерировать нервные импульсы в отличие от других типов нейронов сетчатки (биполярных, горизонтальных, амакриновых).
Сетчатка развивается как вырост переднего мозга, называемый главным пузырьком. В процессе эмбрионального развития глаза фоторецепторный участок пузырька впячивается внутрь до соприкосновения с сосудистым слоем. При этом рецепторные клетки оказываются лежащими под слоем тел и аксонов нервных клеток, связывающих их с мозгом.
2. Строение и функция палочек и колбочек
Палочки и колбочки очень сходны по своему строению: и в тех, и других светочувствительные пигменты находятся на наружной поверхности внутриклеточных мембран наружного сегмента; и те и другие состоят из четырех участков, строение и функции которых кратко описаны ниже.
Наружный сегмент. Это тот светочувствительный участок, где световая энергия преобразуется в рецепторный потенциал. Весь наружный сегмент заполнен мембранными дисками, образованными плазматической мембраной и отделившимися от нее. В палочках число этих дисков составляет 600-1000, они представляют собой уплощенные мембранные мешочки и уложены наподобие стопки монет. В колбочках мембранных дисков меньше, и они представляют собой складки плазматической мембраны.
Перетяжка. Здесь наружный сегмент почти полностью отделен от внутреннего впячиванием наружной мембраны. Связь между двумя сегментами осуществляется через цитоплазму и пару ресничек, переходящих из одного сегмента в другой. Реснички содержат только 9 периферических дублетов микротрубочек: пара центральных микротрубочек, характерных доя ресничек, отсутствует.
Внутренний сегмент. Это область активного метаболизма; она заполнена митохондриями, доставляющими энергию для процессов зрения, и полирибосомами, на которых синтезируются белки, участвующие в образовании мембранных дисков и зрительного пигмента. В этом же участке расположено ядро.
Синаптическая область. В этом участке клетка образует синапсы с биполярными клетками. Диффузные биполярные клетки могут образовывать синапсы с несколькими палочками. Это явление, называемое синаптической конвергенцией, уменьшает остроту зрения, но повышает светочувствительность глаза. Моносинаптические биполярные клетки связывают одну колбочку с одной ганглиозной клеткой, что обеспечивает большую по сравнению с палочками остроту зрения. Горизонтальные и амакриновые клетки связывают вместе некоторое число палочек или колбочек. Благодаря этим клеткам зрительная информация еще до выхода из сетчатки подвергается определенной переработке. Эти клетки, в частности, участвуют в латеральном торможении.
3. Различия между палочками и колбочками
Палочек в сетчатке содержится больше, чем колбочек (120·106 и 6-7·106, соответственно). Распределение палочек и колбочек тоже неодинаково. Тонкие, вытянутые палочки равномерно распределены по всей сетчатке, кроме центральной ямки, где преобладают удлиненные конические колбочки. Так как в центральной ямке колбочки очень плотно упакованы (15·104 на 1 мм2), этот участок отличается высокой остротой зрения. В то же время палочки обладают большей чувствительностью к свету и реагируют на более слабое освещение. Палочки содержат только дин зрительный пигмент, не способны различать цвета и используются преимущественно в ночном зрении. Колбочки содержат три зрительных пигмента, и это позволяет им воспринимать свет; они используются главным образом при дневном свете. Палочковое зрение отличается меньшей остротой, так как палочки расположены менее плотно, и сигналы от них подвергаются конвергенции, но именно это обеспечивает высокую чувствительность, необходимую для ночного зрения.
4. Механизм фоторецепции
Палочки содержат светочувствительный пигмент родопсин, находящийся на наружной поверхности мембранных дисков.
Родопсин (от греч. rhodon – роза и opsis – зрение) – зрительный пурпур, основеной зрительный пигмент палочек сетчатки позвоночных (кроме некоторых рыб и земноводных на ранних стадиях развития) и беспозвоночных животных.
Родопсин представляет собой сложную молекулу, образующуюся в результате обратимого связывания липопротеина (белка) скотопсина с небольшой молекулой поглощающего свет каротиноида – ретиналя. Последний представляет собой альдегидную форму витамина А и может существовать (в зависимости от освещения) в виде двух изомеров.
По химической природе родопсин – это сложный белок (хромопротеид), в состав которого входит 11-цис-ретиналь (хромофорная группа), гликопротеид, то есть белок, соединённый с сахарами, и липиды (опсиновая часть). Молекулярная масса родопсин позвоночных около 40000, головоногих моллюсков - около 70000. Родопсин - основной структурно-функциональный компонент наружного сегмента палочек.
Установлено, что при воздействии света на родопсин один фотон способен вызывать изомеризацию. Ретиналь играет роль простетической группы, и полагают, что он занимает определенный участок на поверхности молекулы скотопсина и блокируют реактивные группы, участвующие в генерации электрической активности в палочках. Точный механизм фоторецепции пока неизвестен, но предполагается, что он включает 2 процесса. Первый из процессов – это превращение 11-цис-ретиналя в полностью-транс-ретиналь под воздействием света, а второй – расщепление родопсина через ряд промежуточных продуктов на ретиналь и скотопсин (процесс, называемый выцветанием).
После прекращения воздействия света родопсин тотчас же ресинтезируется. В начале полностью-транс-ретиналь при участии фермента ретинальизомеразы превращается в 11-цис-ретиналь, а затем последний соединяется со скотопсином. Этот процесс лежит в основе темновой адаптации. В полной темноте требуется около 30 минут, чтобы все палочки адаптировались и глаза приобрели максимальную чувствительность. Однако во время этого процесса проницаемость мембраны наружного сегмента для ионов Na+ уменьшается, в то время как внутренний сегмент продолжает откачивать ионы Na+ наружу, и в результате внутри палочки возрастает отрицательный потенциал, то есть происходит гиперполяризация.
Отрицательные заряды на правой стороне палочки соответствуют потенциалу покоя, а на левой стороне – гипреполяризации.
Это прямо противоположно тому, что обычно наблюдается в других рецепторных клетках, где раздражение вызывает деполяризацию, а не гиперполяризацию. Гиперполяризация замедляет высвобождение из палочек возбуждающего медиатора (посредника), который в темноте выделяется в наибольшем количестве. Биполярные клетки, связанные через синапсы с палочками, тоже отвечают гиперполяризацией, но в ганглиозных клетках, аксоны которых образуют зрительный нерв, в ответ на сигнал от биполярной клетки возникает распространяющийся потенциал действия.
5. Цепь фотопревращений родопсина
Согласно описанному механизму фоторецепции, зрительный акт начинается поглощением родопсином кванта света (максимум спектра поглощения родопсина составляет – около 500 нм). При этом происходит изомеризация 11-цис-ретиналя в полностью транс-форму (см. формулы), что приводит к постепенному разложению (фотолизу) молекулы родопсина, изменению ионного транспорта в фоторецепторе и возникновению электрического сигнала, который передаётся нервным элементам сетчатки. Регенерация родопсина осуществляется или путём синтеза из 11-цис-ретиналя и освободившегося после фотолиза опсина, или при поглощении второго кванта одним из промежуточных продуктов фотолиза, а также в процессе синтеза новых дисков наружного сегмента сетчатки (последний путь для палочек основной).
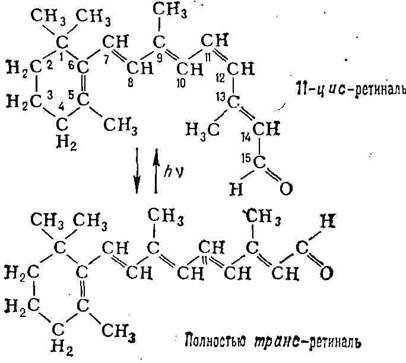
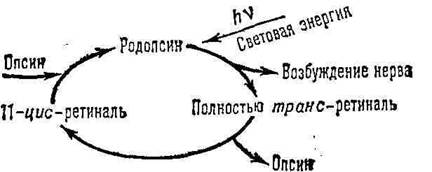
Рис. 1. Цикл основных изменений родопсина в палочках сетчатки
Примечание. В клеточных оболочках некоторых галофильных бактерий обнаружен пигмент, в состав которого также входят ретиналь, гликопротеид и липиды. Этот бактериальный родопсин (структура его окончательно не установлена), по-видимому, участвует в фотосинтезе наряду с другими пигментами бактерий.
Контрольные вопросы
1. Сколько слоев имеет сетчатка?
2. В чем заключаются различия между палочками и колбочками?
3. Как осуществляется механизм фоторецепции.
4. Прокомментируйте цикл основных изменений родопсина в палочках сетчатки.
Домашнее задание
В каких продуктах содержится витамин А?
