
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
К вопросу о геноме последнего общего предка всех земных живых организмов.
В настоящее время среди учёных нет единого мнения о том, что собой представлял последний общий предок всех нынеживущих на Земле организмов. Тем не менее, наиболее общепринятая точка зрения состоит в том, что это был довольно сложный организм, геном которого состоял из более, чем 1000 генов (Ouzounis et al. 2006). Именно эта бактерия, которая к настоящему времени полностью вымерла, и дала начало двум ныне процветающим царствам прокариот – эубактериям и археям (Doolittle 2000). Однако, существует и альтернативная точка зрения, помещающая корень земной жизни среди эубактерий. При таком сценарии археи представляют из себя сравнительно поздно обособившуюся от эубактерий ветвь, быстро проэволюционировавшую под действием мощного пресса естественного отбора возникшего, например, из-за необходимости защиты от антибиотиков, синтезируемых другими бактериями (Valas and Bourne 2011). Автору ближе альтернативная точка зрения, тем более, что в последнее время появляется всё больше аргументов в её пользу. Это и анализ аминокислотного состава рибосомных белков наиболее древних организмов (Fourneier and Gorgarten 2010), и анализ вставок и делеций в общих для всех живых организмов протеинах (Gupta 2001, Lake et al. 2009), и эволюция протеасомы (Cavalier-Smith, 2006), и ряд других соображений. С другой стороны, исследования последних лет показывают, что скорость «тиканья» молекулярных часов у разных таксонов бактерий может различаться в разы (Kuo and Ochman 2009), что делает в существенной степени непригодным традиционный метод поиска на неориентированном филогеническим дереве эволюции его предпологаемого корня путём нахождения его геометрического центра, особенно, в условиях отсутствия надёжно классифицированных палеонтологических данных в породах времён архея. А ведь именно на соответствующих филогенических деревьях (особенно, построенных по рРНК и протеинам, связанным с синтезом белков) и молекулярных часах в значительной степени и держатся основные аргументы сторонников помещения истоков дерева жизни между археями и эубактериями! К сожалению, более глубокое рассмотрение этого интересного вопроса заслуживает отдельной статьи, поэтому, лучше сразу перейдём к результатам, полученных автором по обсуждаемой теме с помощью нового, никем пока не применявшегося (по крайней мере, насколько это известно автору) до сих пор метода.
Идея нового метода заключается в том, что в глубокой древности структура генома живых организмов была гораздо проще, чем в наши дни, когда клетка научилась в совершенстве управлять своими генами, что позволяет ей обеспечивать их синхронную работу, даже если они расположены на разных хромосомах! Но раньше, у истоков жизни, это, по видимому, было не так. Система управления экспессией генов была настолько примитивной, что для её эффективной работы все совместно функционирующие гены, образующие единые белковые комплексы, должны были быть расположены в одном кластере, и работать преимущественно на принципах самосборки. Отголоски тех далёких времён, по всей видимости, до сих пор можно проследить при формировании наиболее древних и консервативных элементов клетки - рибосомы, АТФ-синтазы, жгутика вместе с обеспечивающим его вращение базальным телом и т. д. Если изложенная выше гипотеза верно описывает последовательность формирования современного генетического аппарата прокариот, то можно надеяться, что некие её «родимые пятна» могли дожить до наших дней. В частности, у наиболее архаичных бактерий, геном которых обладает минимальными отличиями от генома общего предка всех живых существ на Земле, должна быть наиболее ярко выражена его оперонная структура и, следовательно, можно ожидать наибольшей средней длины участков генома, на которых направление считывания генов не меняется. К настоящему времени секвенированы уже более тысячи штаммов бактерий, что даёт нам возможность провести их анализ и выявить среди них наиболее экстремальные в смысле максимизации указанного выше параметра. Результаты такого анализа говорят о том, что описанное свойство в наибольшей степени присуще геномам клостридий. Любопытно, что к почти аналогичным выводам (корень дерева жизни лежит внутри типа фирмикут, к которому, в частности, принадлежат и клостридии) независимо пришёл и Гупта на основе совершенно другого метода (Gupta 2001). Близки к этому и выводы Лейка и его группы (Lake et al. 2009) о том, что корень находится между клостридиями и актинобактериями, особенно, если учесть, что от класса клостридий его отделяет всего лишь крохотная (2 аминокислотных основания) вставка в гене PyrD, которая вполне может оказаться результатом конвергентной эволюции, тем более, что это единственный ген, поляризация которого в данной работе по существу внутренне противоречива (Valas and Bourne 2009).
Если мы начнём анализировать особенности геномов клостридий, то сразу же бросается в глаза ещё одно, на первый взгляд, странное обстоятельство. Их кольцевой геном в плане распределения направления чтения генов чётко делится на две части, каждой из которых соответствует своя половинка кольца. Как могло возникнуть столь низкоэнтропийное распределение? Какая сила заставила столь странно упорядочиться геном клостридий? Один из возможных ответов заключается в том, что изначально РНК (или, возможно, ДНК) геном был одноцепочечным, но замкнутым, причём, на протяжении своей большей части, он был свёрнут сам на себя за счёт образования связей через комплиментарные основания. Подобные структуры, (правда, гораздо более скромные по размерам), характеризующиеся наличием склеившихся «стеблей», соединяющих участки с выпячивающимися одноцепочечными «листьями» (их иногда так же называют петлями) широко представлены и в РНК современных организмов, например, в их тРНК. На Рис.1 схематически показана возможная схема организации одноцепочечного генома организма, от которого произошла вся жизнь на Земле. Синим цветом показана клеточная мебрана к которой прикреплена собственно полинуклеотидная цепочка (показана красным цветом) с минимум двумя небольшими петлями и одним длинным стеблем.

Рис. 1
Понятно, что при таком устройстве генома как транскрипция, так и репликация могли осуществляться лишь в одном направлении – от 5' к 3'. Так же понятно, что при этом единственная цепочка состоит из двух половинок, взаимно комплиментарных друг к другу. Далее, представим, что единственная цепочка генома периодически реплицировалась путём полимеризации новой цепочки, комплиментарной к исходной. Очевидно, что при этом вновь синтезированная цепочка должна была быть комплиментарна к исходной, а в функциональном плане была полностью ей аналогична с той лишь разницей, что «рабочей» становилась уже её другая половинка. И всё было бы хорошо, но однажды по какой-то причине вновь синтезированная цепь не отделилась от родительской, а «приклеилась» к ней обратно, одновременно разорвав связи между комплиментарными участками одной и той же цепочки. В результате, получился ставший впоследствии классическим кольцевой бактериальный геном. Он оказался гораздо более устойчивым к разрушающим воздействиям, и по этой причине быстро вытеснил архаичный одноцепочечный геном, который остался в виде реликта лишь в некоторых вирусах.
Как можно проверить правдоподобность описанного выше сценария? Если двухцепочечный геном действительно произошёл от одноцепочечного описанным выше способом, то первые организмы с двойной спиралью из нуклеиновых кислот должны были обладать геномом, в котором все гены одной половинки кольца были ориентированы в одну сторону, а все гены другой его половинки – в противоположную. Именно картину, максимально приближенную к описанной, мы и наблюдаем в геноме клостридий. Причём, среди клостридий, в свою очередь, есть отряд галанаэробных (Halanaerobiales) бактерий, представители которого выделяются экстремальностью значений признака анизотропности распределения направления чтения генов по половинкам хромосомы даже на фоне других отрядов клостридий. В частности, представитель данного отряда Acetohalobium arabaticum DSM 5501 имеет геном, у которого степень анизотропии достигает 90%, то есть, в каждом полукольце его хромосомы на 1 ген с направлением «против шерсти» встречается около 10 генов с противоположным направлением считывания. На Рис. 2 приведена графическая диаграмма распределения его генов по различным нитям ДНК из которой видно, насколько сильно различается плотность локализации генов на различных цепочках в зависимости от их расположения. В нижней и верхней точках кольца хромосомы заселённость условных «внешней» и «внутренней» нитей меняется резким скачком.
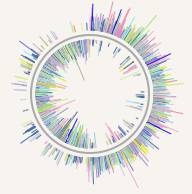
Рис. 2
Возможно, приведённые факты убедят далеко не всех скептиков, поэтому, попробуем поискать дополнительные аргументы в пользу высказанной гипотезы. Достаточно очевидно, что если она справедлива, то это означает, что сразу же после перехода от одноцепочечного к двухцепочечному геному должна была возникнуть не только резкая ассимметрия в направлении чтения генов, но и сам набор генов в обоих половинках хромосомы изначально должен был быть одинаковым. Но затем неизбежно должен был начать работать естественный отбор, убирающий ненужные дупликаты генов, так как на их месте в большинстве случаев гораздо выгоднее было хранить другие, приобретённые уже после указанного перехода гены. Однако, с учётом того, что пути потомков общего предка постепенно расходились, отмирание каждой лишней копии гена должно был происходить примерно с равной вероятностью либо на одной, либо на другой цепочке. К чему это должно было привести на практике? Очевидно, к тому, что у разных видов бактерий одни и те же гены часто должны были располагаться на разных цепочках хромосомы практически идеально симметрично друг от друга относительно места прикрепления хромосомы к мембране. Наблюдается ли подобный эффект в отряде галанаэробных бактерий? Да, ответ утвердительный. Сравним, например, локализацию одних и тех же кластеров генов, расположенных на разных нитях ДНК у бактерий Halanaerobium praevalens DSM 2228 и Halanaerobium sp. 'sapolanicus'. Кластер, начинающийся с гена tRNA-adenosine deaminase (COG0590FJ), и заканчивающийся геном tetrapyrrole (Corrin/Porphyrin) methyltransferase (COG0313R) расположен у этих бактерий на разных цепочках, но на расстояниях от центра начала репликации хромосомы, отличающихся менее, чем на две десятые процента от её общей длины. Аналогично, кластер из трёх генов, начинающийся с гена extracellular solute-binding protein family 3 (COG0834ET), и заканчивающийся геном amino acid ABC transporter ATP-binding protein (COG1126E) расположен в них на разных цепочках на расстояниях, отличающихся менее, чем на одну десятую процента от общей длины хромосомы. Огромный кластер, включающий более 30-ти генов, который начинается с участка хромосомы, кодирующего протеин arginyl-tRNA synthetase (COG0018J), и заканчивается участком, кодирующим протеин lipid-A-disaccharide synthase (COG0763M) локализован на расстояниях от начала хромосомы, отличающихся менее, чем на четыре десятых процента от её длины и так далее (список можно продолжить). Вероятность того, что такие множественные совпадения являются случайными, ничтожно мала, несомненно, для них должна существовать какая-то причина. Автору не удалось придумать изложенным фактам никакого другого объяснения, кроме близости галанаэробных бактерий к общему предку всего живого на нашей планете. При этом, можно отметить, что указанный отряд бактерий является для клостридий достаточно уникальным в том смысле, что его представители являются грам-отрицательными, то есть, обладают двойной внешней мембраной, что, вообще говоря, нехарактерно для фирмикут вообще, и для клостридий в частности. Этот факт тем более любопытен, что он в какой-то мере ликвидирует противоречия между выводами, следующими из анализа вставок и делиций, помещающими истоки дерева жизни внутри (или около) фирмикут, и анализом Кавальер-Смита, приводящим множественные аргументы в пользу того, что наш общий предок обладал двойной мембраной.
Стоит отметить, что если грубая общая схема, изложенная выше, верна, это означает, что, так сказать, «сердце генома», то есть, его наиболее архаичная и консервативная часть, ассоциирующаяся с районом прикрепления хромосомы к клеточной мембране, должна содержать преимущественно самые древние гены, ответственные за наиболее базовые функции внутриклеточной фабрики – размножение, добычу энергии, синтез белков и т. д. Во многом, так оно и есть. С другой стороны, продвигаясь по хромосоме в обе стороны от места её прикрепления к мембране, можно ожидать, что мы в какой-то степени сможем проследить историю жизни на Земле, по крайней мере, до той поры, пока геном ещё более-менее сохранял свою архаичную низкоэнтропийную структуру. Но это, очевидно, уже тема для другой статьи...
Список литературы.
Cavalier-Smith T. 2006. Rooting the tree of life by transition analyses // http://www. /content/1/1/19
Doolittle W. F., 2000. Uprooting the tree of life // Scientific American 282 (6): 90-95
Fourneier G. P., Gorgarten J. P., 2010. Rooting the Ribosomal Tree of Life // shiva. msu. montana. edu/courses/mb428_537_2010_fall/ToL. pdf
Gupta, R. S. 2001. The branching order and phylogenetic placement of species from completed bacterial genomes, based on conserved indels found in various proteins // Inter. Microbiol. 4, 187-202.
Kuo C. H., Ochman H., 2009. Inferring clocks when lacking rocks: the variable rates of molecular evolution in bacteria // http://www. /content/4/1/35
Lake J. A., Skophammer R. G., Herbold C. W., Servin J. A. 2009. Genome beginnings: rooting the tree of life // Philos Trans R Soc Lond B Biol Sci 2009, 364(1527):2177-2185.
Ouzounis C. A., Kunin V., Darzentas N., Goldovsky L., 2006. A minimal estimate for the gene content of the last universal common ancestor–exobiology from a terrestrial perspective // Res Microbiol. 2006; 157:57–68
Valas R. E., Bourne P. E., 2011. The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon // http://www. /content/6/1/16
Valas R. E., Bourne P. E., 2009. Structural analysis of polarizing indels: an emerging consensus on the root of the tree of life // http://www. /content/4/1/30
