
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
УДК 574.24:595.762.12
МОРФОМЕТРИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И ПОЛОВОЙ ДИМОРФИЗМ В ПОПУЛЯЦИЯХ ЖУЖЕЛИЦЫ CARABUS CANCELLATUS L. (COLEOPTERA, CARABIDAE)
Институт проблем экологии и недропользования
АН Республики Татарстан
Россия 420087, Казань Даурская, 28
Поступила в редакцию
Размер тела – фундаментальный морфологический признак, важный в физиологическом, экологическом и социальном аспектах. Для многих видов характерна географически структурированная изменчивость в морфологии, особенно по размерам (Avise, 2000). Однако механизмы, определяющие и поддерживающие географическую изменчивость по размерам очень сложны и не всегда поддаются точному описанию (Case, Schwaner, 1993; Patridge, French, 1996; Baez, Brown, 1997; Malhorta, Thorpe, 1997).
Было предложено несколько механизмов поддержания изменчивости размеров и формы. Они имеют как адаптивную, так и неадаптивную природу: отбор в ответ на географическую изменчивость типа добычи или давления хищника, влияние климата или других факторов среды на скорость роста (Van Der Have, De Jong, 1996), изменчивость силы давления полового отбора и половой диморфизм (Endler, Haude, 1995; Storz et al, 2001). Было сформулировано несколько общих экогеографических правил, касающихся морфометрической изменчивости. Одно из наиболее известных - правило Бергмана, согласно которому размер тела увеличивается с увеличением широты. Это правило подтверждается многими исследованиями на эндотермах, однако многие представители эктотермов демонстрируют прямо противоположную зависимость (Mousseau, 1997; Sota et al, 2000).
МАТЕРИАЛ И МЕТОДИКА.
Объект исследования. Carabus cancellatus (русское название – жужелица полевая) - крупный жук от 18 до 30 мм в длину. Надкрылья выпуклые, с цепочками бугорков и ребрышками. Между каждыми двумя продольными рядами ямок на надкрыльях различимы не более пяти промежутков. Верх медный или бронзовый, редко темно-синий, темно-зеленый или бронзово-черный, 1-й членик усиков и часто бедра красные. В умеренных широтах взрослые особи встречаются с апреля по сентябрь. Кладка в июне-июле. Через два месяца появляются молодые жуки. Зимует имаго (Жеребцов, 2000; Исаев, 2004). Переваривание пищи (часто это падаль), как у большинства жужелиц, внеполостное, с помощью отрыгнутого пищеварительного сока.
Неморальный палеарктический вид, распространен в Центральной Европе, за Уралом встречается спорадически вплоть до Байкала. Интродуцирован в Северную Америку. Преимущественно равнинный, с южностепным ареалом (Воронин, Есюнин, 2006). Обитает также в пойменных и нагорных лесах, предпочитая места с умеренным увлажнением, часто под камнями, бревнами (Сажнев, www. zin. ru/animalia/coleoptera/rus/carabsar. htm). Приурочен к глинистым почвам, хотя встречается также в песчаных почвах под сельскохозяйственными культурами. Принадлежит к брахиоптерам, следовательно, бескрылый, миграционные способности ограничиваются хождением и беганьем. Может распространяться потоками воды во время половодья, расширяя, таким образом, свой ареал. В Финляндии зарегистрировано активное плавание жук этого вида, в результате чего происходит заселение островов. Ночной хищник, однако в репродуктивный период проявляет и дневную активность (Erwin, 2001; Larochelle & Lariviere, 2003; Lindroth et al., 1985; Data from NMNH collection)
Районы исследования. Выборки жуков были взяты из разных частей ареала, в каждом из которых отлов проводился три последовательных сезона.
Методика исследования.
Материал из Удмуртии любезно предоставлен доцентом Ижевского Государственного университета к. б.н. (сборы 2006 – 2008 г. г., 73 особи). Жуки отловлены в национальном парке «Нечкинский», который расположен в долине Средней Камы в пределах Воткинского, Завьяловского и Сарапульского районов Удмуртии. Жуки были собраны в липняке правого и левого коренного берега р. Кама в окрестностях с. Нечкино.
Материал из заказника «Предуралье» (Пермский край Ординский район) собран под руководством д. б. н., профессора Пермского Государственного университета в биотопе березняк.
В Республике Татарстан материал собирали в период 1996 – 2006 г. г. Исследуемые участки располагались на территориях с разным антропогенным воздействием:
- г. Казань – обследовано семь биотопов в разных частях города (четыре биотопа представляли кустарники и три - лесные массивы в пределах города, липняки);
- десять биотопов в пригороде (вязовник, три березняка, три сосняка, три кустарниковых биотопа в окрестностях г. Казани близ поселений Пановка, Бирюли, Нагорный в Пестречинском районе Республики Татарстан);
- агроценозы – поле с ячменем и малинник близ с. Масловка Рыбно – Слободского районы Республики Татарстан и два поля с вико – овсяной смесью близ с. Нармонка Лаишевского района Республики Татарстан;
- естественные биотопы – Зеленодольский район (вязовник и березняк на территории Волжско – Камского государственного заповедника и березняк близ пос. Октябрьский), липняк близ с. Новошигалеево Пестречинского района Республики Татарстан.
Жуков отлавливали стандартным методом ловушками Барбера. В каждом биотопе ставили линии по 10 ловушек, диаметром 8 см, которые находились в рабочем состоянии 5 суток. Таким образом, на территории Татарстана в общей сложности отработано 3250 ловушко/суток, отловлено около 2000 особей исследуемого вида.
Морфометрические промеры проводили под бинокулярным микроскопом МБС – 9 при увеличении 1х8. После определения пола у каждого жука оценивали с помощью окуляр-микрометра шесть мерных признаков: длина надкрылий – расстояние по шву от середины бортика до вершины; ширина надкрылий – расстояние между плечевыми углами левого и правого надкрылий, длина переднеспинки – расстояние по средней линии от основания до вершины, ширина переднеспинки – ширина основания, длина головы – расстояние от шеи до верхней губы, расстояние между глазами. Все результаты приводятся в единицах мерной линейки, цена деления равна 0,1 мм. В общей сложности проанализировано 1792 особи.
Статистическая обработка результатов. Для оценки влияния различных факторов на мерные характеристики жуков использовалась линейная модель. В качестве независимых переменных рассматриваются регион, антропогенная нагрузка и биотоп. Влияние всех остальных переменных считается случайным и суммарно оценивается в виде члена ошибки в модели. Используемые переменные рассматриваются как категориальные, в качестве базы сравнения для них используются: для региона – Республика Татарстан, для антропогенной нагрузки – естественные ценозы, для биотопа – березняк (как наиболее оптимальный для исследуемого вида биотоп). Вклады региона, антропогенной нагрузки и характера биотопа в морфометрическую изменчивость популяций исследуемого вида считаются аддитивными и независимыми.
Влияние перечисленных факторов считается различным для пола жука, кроме того, учитывается и собственно влияние пола. Другими словами, в модели участвует пол жука и парные взаимодействия пола с каждым из перечисленных факторов.
В записи на языке R модель зависимости, в частности для длины надкрылий, имеет вид: Elytra. Length~/fSex (fRegion+fAntrop+fHabitat), где fSex код пола, fRegion – код региона, fHabitat – код биотопа. Контрасты для кодов выбраны так, что базовыми (соответствующими нулевым значениям в матрице модели) для каждого из них являются перечисленные выше значения.
Для оценки значимости факторов использовался дисперсионный анализ построенной модели. Для каждого признака оценивался вклад всех переменных и взаимодействий с указанием доверительных интервалов и значимости (по критерию Стьюдента) и статистики остатков (ошибок).
Полученные оценки и их доверительные интервалы использовались для представления результатов в графическом и табличном видах: взаимодействия сравнивались с базой сравнения (использовался 95% доверительный интервал для нормальной аппроксимации). Кроме того, выводились доверительные интервалы для суммы вклада пола и отдельных переменных.
Учитывая значительную изменчивость размеров тела насекомых, исследование спланировано так, чтобы материал был взят из разных точек ареала, в градиенте урбанизации и возможно большем спектре типов биотопов: весь массив данных по исследуемому виду используется как матрица для факторного анализа в программе R. Это позволяет вычленить в изменчивости размеров жука не только половую компоненту, но и отдельно влияние местообитания в ареала и связанные с ним различия в температурных условиях развития и различиях в длине репродуктивного периода, антропогенное влияние как условия обитания жуков в городе, пригороде и естественных местообитаниях и влияние растительности на размер жуков. Для большей иллюстративности на рисунках каждый тип фактора, влияющего на размеры жука, обозначен своим символом: влияние региона - @, антропогенное влияние - %, влияние растительности - $.
Результаты. На длину надкрылий C. cancellatus значимо влияют условия обитания в Удмуртии, в городе и в ячмене, сдвигая значение этого признака в сторону уменьшения, по сравнению с оптимальными. Значимо увеличивается длина надкрылий у этого жука под влиянием обитания в сосняке, кустарнике и вико – овсяной смеси. Только на длину надкрылий самцов влияют условия агроценоза в сторону увеличения (рис. 1).

Рис. 1. Изменение длины надкрылий у C. cancellatus при действии разных факторов
Ширина надкрылий исследуемого вида жужелиц значимо увеличивается в условиях Удмуртии, под влиянием города и агроценоза, в вязовнике у самок и вико – овсяной смеси у особей обоих полов. Уменьшение признака происходит под влиянием обитания в кустарнике и ячмене (рис. 2).

Рис. 2. Изменение ширины надкрылий у C. cancellatus при действии разных факторов
По длине переднеспинки влияние обитания в Удмуртии значимо уменьшают этот признак, по сравнению с жуками из РТ. Влияние обитания в пригороде выражается в значимом увеличении длины переднеспинки у самок, а в агроценозе – у жуков обоих полов. Влияние типа растительности выражается в том, что и сосняк, и кустарник положительно влияют на этот признак, а ячмень – отрицательно (рис. 3).

Рис. 3. Изменение длины переднеспинки у C. cancellatus при действии различных факторов
Ширина переднеспинки у особей исследуемого вида увеличивается в условиях Удмуртии, пригорода, под влиянием сосняка, кустарника, ячменя и вико - овсяной смеси у саамов, и значимо уменьшается у самок в Предуралье, под влиянием города и агроценоза (рис. 4).

Рис. 4. Изменение ширины переднеспинки у C. cancellatus при действии различных факторов
Длина головы у C. cancellatus увеличивается только при обитании в кустарнике и вико – овсяной смеси, значимо уменьшается под влиянием антропогена в городе и пригороде, а также в ячмене (рис. 5).
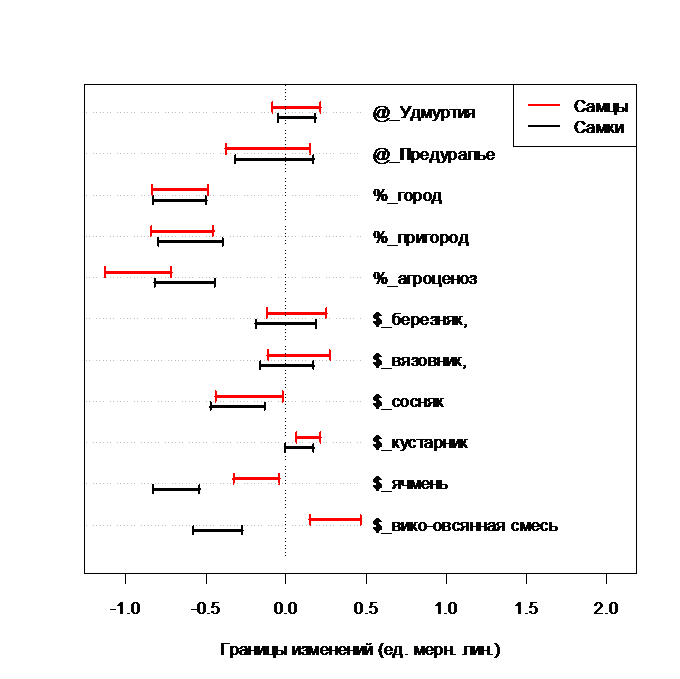
Рис. 5. Изменение длины головы у C. cancellatus под влиянием различных факторов
Расстояние между глазами изменяется у жуков исследуемого вида под влиянием факторов различной природы незначительно: оно больше у самок в Удмуртии, уменьшается у самок в ячмене и увеличивается у самцов в вико – овсяной смеси (рис. 6).
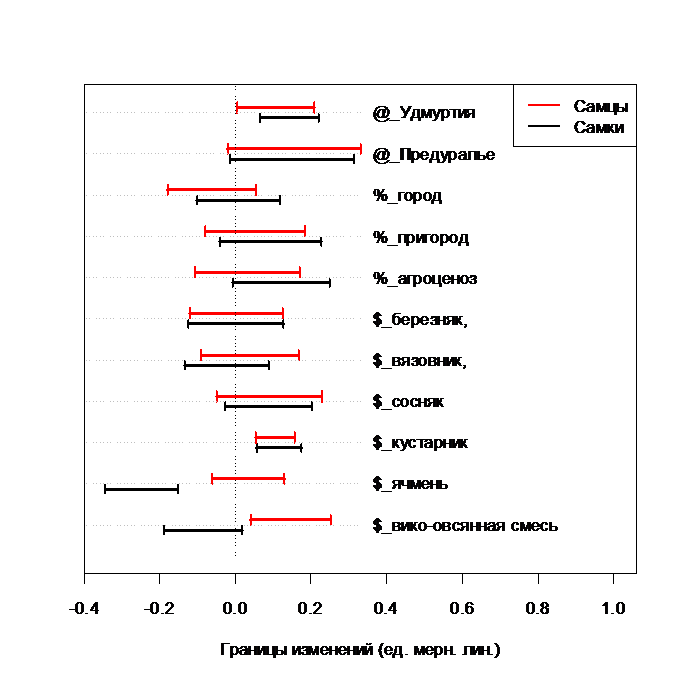
Рис. 6. Изменение расстояния между глазами у C. cancellatus при действии разных факторов.
Проведенный анализ позволяет заключить, что у C. cancellatus регистрируется явно выраженный половой диморфизм по размерам. Величина его может меняться при действии на жуков этого вида факторов различной природы. Это является результатом того, что самки и самцы по – разному реагируют на воздействие таких факторов. Статистически достоверные различия между самками и самцами этого вида жужелиц регистрируются при действии следующих факторов: обитание в городе приводит к значимому ПД по длине надкрылий, переднеспинки, головы и расстоянию между глазами; обитание в пригороде – значимо влияет на ПД только по длине надкрылий; в агроэкосистеме значимый ПД наблюдается по признакам ширины – надкрылий и переднеспинки и длине головы. Растительность биотопа также неоднозначна во влиянии на степень выраженности ПД по разных признакам: обитание в лесных биотопах приводит к значимому ПД только по признакам надкрылий, а обитание в агрокультурах – практически по всем признакам.
На рис. 7. приведены обобщенные данные по половому диморфизму по исследованным признакам у C. cancellatus, когда анализировался весь материал в целом, вне зависимости от действия экологических факторов. Видно, что наиболее диморфны у этого вида признаки надкрылий – длина и ширина.
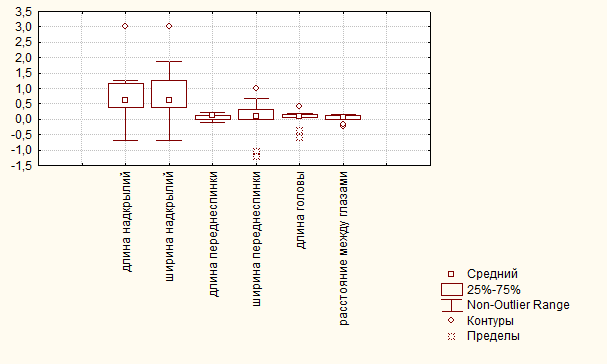
Рис. 7. Значение ПД по разным признакам у C. cancellatus (в среднем при действии всех изученных факторов).
Проведенный компонентный анализ позволил определить ПД по форме у исследуемого вида жужелиц. Авторы, занимающиеся морфометрией объектов, считают, что по результатам компонентного анализа можно вычленить изменчивость формы объектов из общей изменчивости признаков. Первая (главная) компонента отражает изменчивость размеров, а вторая и остальные – изменчивость формы. В табл. 1, 2 представлены результаты анализа главных компонент отдельно в выборках самок и самцов исследуемого вида жужелиц.
Таблица 1. Результаты анализа главных компонент для самок Carabus cancellatus | ||||||
Признак | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 |
Длина надкрылий | -0.48 | 0.00 | 0.45 | -0.34 | -0.46 | -0.49 |
Ширина надкрылий | -0.10 | 0.74 | -0.49 | 0.01 | 0.08 | -0.44 |
Длина переднеспинки | -0.47 | -0.21 | 0.03 | 0.80 | 0.18 | -0.23 |
Ширина переднеспинки | -0.39 | -0.29 | -0.68 | -0.10 | -0.46 | 0.27 |
Длина головы | -0.36 | 0.56 | 0.30 | 0.14 | -0.13 | 0.66 |
Расстояние между глазами | -0.50 | -0.12 | -0.05 | -0.46 | 0.72 | 0.09 |
Доля объясненной дисперсии | 47.1% | 22.3% | 12.4% | 7.3% | 6.0% | 4.9% |
Анализ Таблицы 1 показывает, что аллометрия у самок наиболее выражена для ширины надкрылий, и «компенсируется» второй главной компонентой (которая одновременно «отвечает» за длину головы, изменение которой однонаправлено с шириной надкрылий).
Таблица 2. Результаты анализа главных компонент для самцов Carabus cancellatus | ||||||
PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
Длина надкрылий | -0.44 | -0.26 | 0.26 | -0.16 | -0.78 | 0.18 |
Ширина надкрылий | -0.39 | 0.63 | -0.30 | 0.1 | 0.03 | 0.60 |
Длина переднеспинки | -0.41 | -0.47 | 0.15 | 0.66 | 0.35 | 0.18 |
Ширина переднеспинки | -0.42 | 0.49 | 0.34 | 0.19 | 0.00 | -0.66 |
Длина головы | -0.31 | -0.22 | -0.83 | 0.01 | -0.12 | -0.38 |
Расстояние между глазами | -0.45 | -0.16 | 0.12 | -0.7 | 0.51 | 0.03 |
Доля дисперсии | 47.7% | 15.7% | 14.9% | 8.5% | 7.8% | 5.4% |
Анализ Таблицы 2 показывает, что у самцов аллометрия менее выражена, чем у самок, и наибольшей является для длины головы. Вторая главная компонента отражает разнонаправленное изменение ширины надкрылий и длины головы, а также длины и ширины переднеспинки.
Таким образом, первая главная компонента отражает изменение «масштаба» жужелицы (синхронное изменение признаков с учетом аллометрии). И поскольку PC1 объясняет около половины дисперсии, то все методы анализа, основанные на расстояниях, будут отражать, в основном, именно эту «масштабную» изменчивость размеров, в то время как нас интересует также структурные изменения размеров (изменение формы жужелицы) под воздействием различных факторов. Эту изменчивость формы мы будем оценивать как разность между размерными признаками, и их значениями, полученными из первой главной компоненты. Таким образом, предметом анализа будет отклонение размерных признаков от общей тенденции, характерной в целом для данного вида.
Ниже приведены результаты ординации исходных данных методом главных компонент, а также результаты неметрического многомерного дистанционного шкалирования для изменчивости формы (в указанном выше смысле) с использованием Евклидова расстояния при анализе действия антропогенных факторов на жуков исследуемого вида (рис. 8 -11). Из рисунков видно, что самки и самцы по-разному располагаются в плоскости главных компонент, что говорит о разной структурной организации их выборок. Особенно заметно становится разница при многомерном дистанционном шкалировании, результаты которого отражают степень отклонений формы самок и самцов от общей тенденции, характерной для особей данного вида жужелиц.
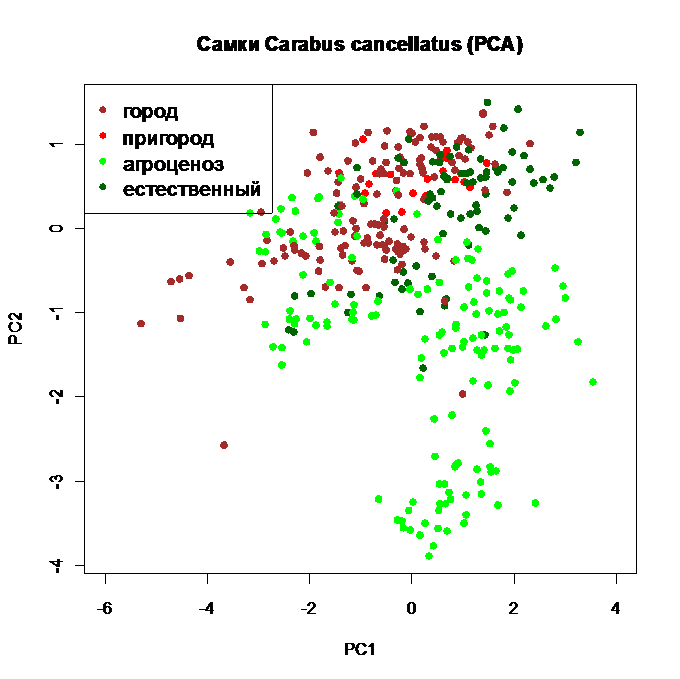
Рис. 8. Ординация самок Carabus cancellatus по размерным признакам методом главных компонент, исходные стандартизованные данные
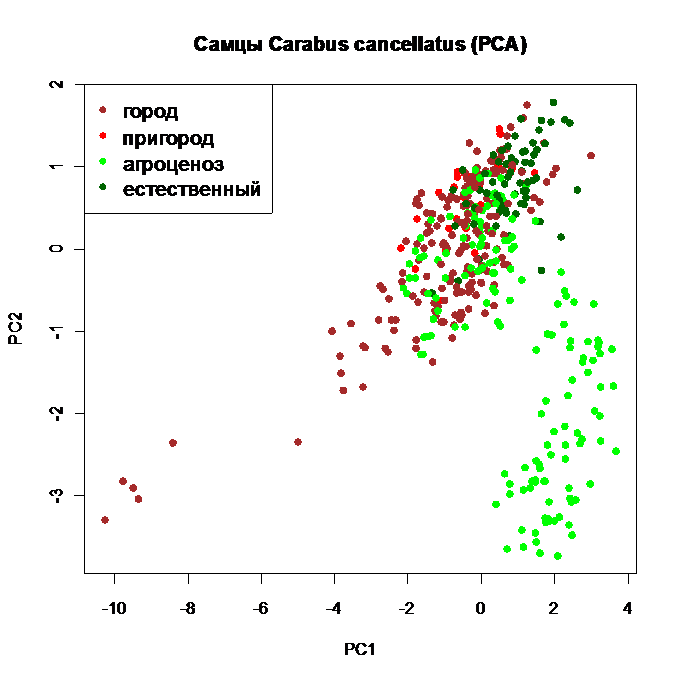
Рис. 9. Ординация самцов Carabus cancellatus по размерным признакам методом главных компонент, исходные стандартизованные данные
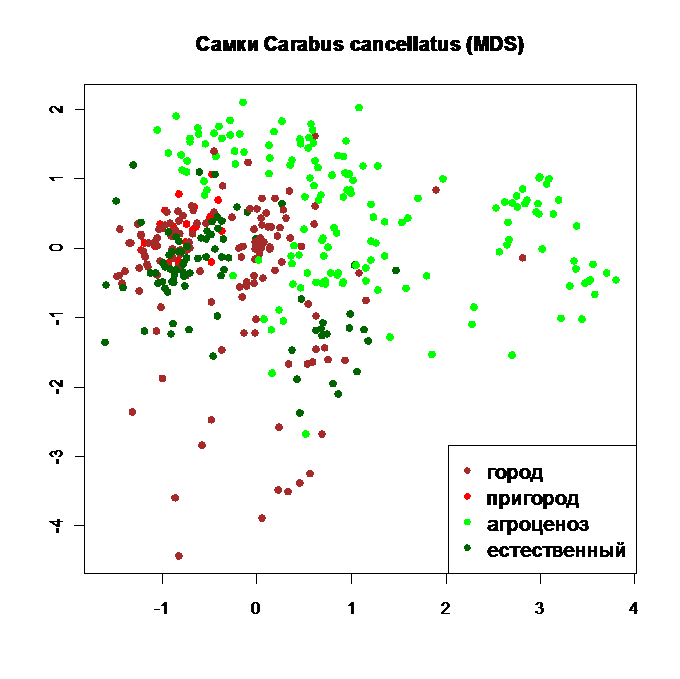
Рис. 10. Ординация самок Carabus cancellatus по отклонениям размерных признаков от общей закономерности методом нелинейного многомерного дистанционного шкалирования
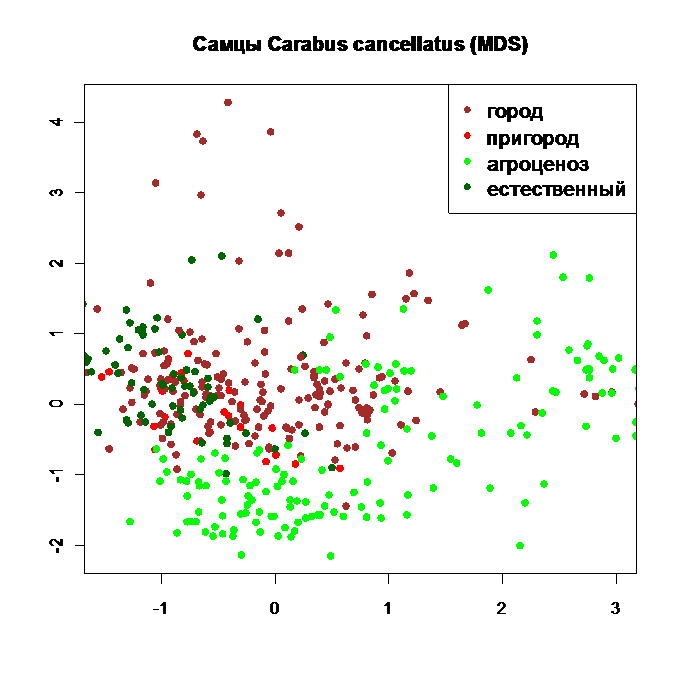
Рис. 11. Ординация самцов Carabus cancellatus по отклонениям размерных признаков от общей закономерности методом нелинейного многомерного дистанционного шкалирования
Обобщенные результаты компонентного анализа по форме представлены на рис. 12. На «иконках» показано, как меняется расположение выборок C. cancellatus при действии факторов различной природы. Анализ рис. 12 приводит к выводу, что половой диморфизм по форме у жужелицы изучаемого вида может принимать различное значение в разнящейся экологической среде.
|
|
|
|
|
|
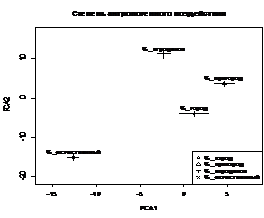





Рис. 12. Положение центров выборок C. cancellatus в плоскости PCA (анализ изменения формы под
действием факторов разной природы). Слева – самки, справа – самцы.
Благодарности. Автор глубоко признателен сотрудникам лаборатории педобиологии Института экологии природных систем АН РТ за помощь в камеральной обработке материала. Особую благодарность выражаю д. б. н. проф. за квалифицированную помощь в статистической обработке материала.
Список литературы
1. , Есюнин – географические виды жужелиц // Экология и основы природопользования. 2006. С. 141 – 144.
2. Жеребцов, жужелиц Республики Татарстан / . – Казань: «Форт-диалог», 2000. - 74 с.
3. Исаев (Insecta, Coleoptera) лесостепи Среднего Поволжья.// , , – Ульяновск: Изд-во УлГУ, 2004.–72 с.
4. Van Voorhies, W. A. Bergmann size clines: A simple explanation for their occurrence in ectotherms. Evolution, V. 50, 1996. P.1259-1264.
5. Avise J. S. Phylogeography: the history and formation of species. Cambridge, M. A: Harvard University Press. 670 p.
6. Baez M., Brown R. P. Testing multivariate patterns within-island differentiation in Polarcis dugestii from Madeira // Journal of evolutionary biology, 1997. V. 10. P. 575 – 587.
7. Case T. J., Schwaner T. D. Island/mainland body size differences in Australian varanid lizards // Oecologia, 1993. V. 94. P. 102 – 109.
8. Endler J. A., Haude A. E. Geographic variation in female preference for mate traits in Poecilia reticulate // Evolution, 1995. V. 49. P. 456 – 468.
9. Erwin T., 2001; canopy. lifedesks. org/pages/605.
10. Larochelle & Lariviere, 2003; Larochelle, A. and M-C. Larivière. 2003. A Natural History of the Ground-Beetles (Coleoptera: Carabidae) of America north of Mexico. Pensoft. Sofia, Bulgaria. 583 p.
11. Lindroth CH. The Carabidae (Coleoptera) of Fennoscandia and Denmark. Volume 15, part 1. E. J. Brill, Scandinavia Science Press Ltd., Leiden-Copenhagen. 1985. 200 p.
12. Malhorta A., Thorpe R. S. Size and shape variation in a Lesser Antilleon anole, Anolis oculatus (Sauria: Iguanidae) in relation to habitat // Biological Journal of Linnean Society, 1997. V. 60. P. 53 – 72.
13. Mousseau T. A. Ectoterms follow converse Bergmann s rule // Evolution, 1997. V. 51. P. 630 – 632.
14. Patridge L., French V. Thermal evolution of ectoterm body size: why get big in cold? // In: Johnston I. A., Bennet A. F., ads. Animals and temperature: phenotypic and evolutionary adaptation. Cambridge, 1996. Cambridge University Press. P. 265 – 292.
15. Sota T., Takami Y., Kubota K., Ishikawa R. Geographic variation in the body size of some Japanese Leptocarabus species (Coleoptera, Carabidae): The “Toppled-domino pattern” along a geographic cline // Entomological Science, 2000. V. 3. P. 309 – 320.
16. Storz J. F., Balasingh J., Bhat H. R., Nathan P. T., Doss D. P. S., Prakash A. A., Kunz T. H. Clinal variation in body size and sexual dimorphism in Indian fruit bat, Cynopterus shinx (Chiroptera: Pteropodidae) // Biological Journal of Linnean Society, 2001. V. 72. P. 17 -31.
17. Van der Have, T. M., and G. de Jong. 1996. Adult size in ectotherms: Temperature effects on growth and differentiation. J. Theor. Biol, V. 183, 1996. P. 329-340.
